
Abstract
Background:
Hepatitis E virus (HEV) is a zoonotic pathogen. HEV has been found to be widely prevalent in rabbits. Its isolates are classified into HEV-3, rabbit subgenotype (HEV-3ra). The routes of human infection with HEV-3ra remain unclear; however, foodborne transmission is possible when asymptomatically infected animals enter the food chain. The prevalence of HEV infection in slaughtered rabbits and the presence of HEV in rabbit meat were evaluated in this study.
Materials and Methods:
In three slaughterhouses in Hebei province, China, samples of rabbit blood were collected during the slaughter process, and muscle, liver, and cavity juice were collected from the rabbit carcasses. Anti-HEV antibody in serum samples was detected using enzyme-linked immunosorbent assay. HEV RNA was tested in all samples by reverse transcription nested PCR (RT-nested PCR). The final amplicons of RT-nested PCR were sequenced and phylogenetically analyzed.
Results:
Of the 459 serum samples, 50 [10.9%, 95% confidence interval (CI): 8.1–13.7] were positive for anti-HEV antibody, and 17 (3.7%, 95% CI: 2.0–5.4) were positive for HEV RNA. HEV RNA was detected in 7 of 60 liver samples (11.7%, 95% CI: 3.3–20) and 2 cavity juice samples from semi-eviscerated carcasses, but was not detected in any muscle sample from either the eviscerated or semi-eviscerated carcasses. All the detected HEV strains belonged to HEV-3ra and related most closely with the rabbit HEV sequence previously reported in China.
Conclusion:
A portion of rabbits were in the viremia period of HEV infection at the slaughter age, resulting in the possibility of HEV carriage by rabbit carcass, particularly semi-eviscerated carcass containing liver. These findings suggest a potential risk of HEV transmission from raw rabbit products entering the food chain, whereas the presence of HEV appeared to be lower in the eviscerated carcass than in the semi-eviscerated carcass.
Introduction
Hepatitis E is an infectious disease prevalent in both developing and developed countries. The causative agent of hepatitis E is hepatitis E virus (HEV). HEV infects humans and a wide variety of animals. HEV harboring in some mammal animal reservoirs can transmit across species and cause hepatitis E in humans. Thus, hepatitis E has been recognized as a zoonotic disease. The well-documented HEV strains to infect humans belong to the family Hepeviridae, subfamily Orthohepevirinae, and genus Paslahepevirus (Purdy et al., 2022). The only species Balayani within genus Paslahepevirus currently includes eight distinct genotypes (HEV-1–8) (Ahmed and Nasheri, 2022; Purdy et al., 2022; Smith et al., 2020). Among them, HEV-3, -4, and -7 are zoonotic, and HEV-3 and -4 are recognized as the predominant causative agents of hepatitis E in industrialized countries (Smith et al., 2020; Wang and Meng, 2021). Foodborne transmission through consumption of animal meat is likely the most important route of HEV-3 and -4 infection for the general population with no occupational contact with animals (Wang and Meng, 2021).
Pigs are the primary reservoir animals for HEV-3 and -4, and rabbits have been found to be widely distributed in many countries (Ahn et al., 2017; Burt et al., 2016; Xia et al., 2015). HEV isolates from rabbits are classified into HEV-3, as a unique rabbit HEV subtype (HEV-3ra) (Smith et al., 2015). It was found that HEV-3ra can infect pigs and macaques experimentally (Cossaboom et al., 2012; Liu et al., 2013; Liu et al., 2017), suggesting its potential zoonotic risk. The discovery of human HEV strains closely related to rabbit HEV supports the possibility of zoonotic transmission from rabbits to humans (Abravanel et al., 2017; Kaiser et al., 2018).
Three solid organ transplant recipients in Switzerland were found to be infected with HEV-3ra and two of them developed chronic hepatitis E, confirming that rabbit HEV can infect and cause chronic hepatitis E in humans, especially in a setting of immunosuppression (Sahli et al., 2019). Remarkably, the majority of reported patients infected with HEV-3ra had no direct contact with rabbits, suggesting foodborne or waterborne infections. Studies have gradually demonstrated the significance of HEV transmission from rabbits to humans.
The prevalence of HEV in farmed rabbits has been widely investigated. However, data on the HEV presence in rabbit meat entering the food chain are lacking. Recently, Bigoraj et al. (2020) reported a high prevalence of HEV in rabbit liver entering the food chain, suggesting that consumption of undercooked or raw rabbit meats may represent sources of human infection. The purpose of this study was to investigate rabbit infection rates at slaughter and the presence of HEV in rabbit carcass in Hebei Province, China, to assess the potential risk of HEV transmission through rabbit meat consumption.
Materials and Methods
Sample collection and processing
Rabbit samples were collected from three slaughterhouses (designated A, B, and C) located in different regions in Hebei Province, China. The slaughterhouses routinely purchase rabbits aged about 3–6 months from nearby farms and supply processed rabbit meat to market. The slaughtering streamlined procedures include stunning, bleeding through hanging by the neck, evisceration, head and foot removal, washing, and freezing for storage. The slaughterhouses mainly sell rabbit meat in two forms of whole carcasses: eviscerated carcass, in which all internal organs have been removed, and semi-eviscerated carcass, which contains some edible organs such as the liver, heart, and kidneys. In the study, rabbit blood, muscle, livers, and cavity juice were collected for HEV detection (Fig. 1). All samples were collected during the slaughter process.
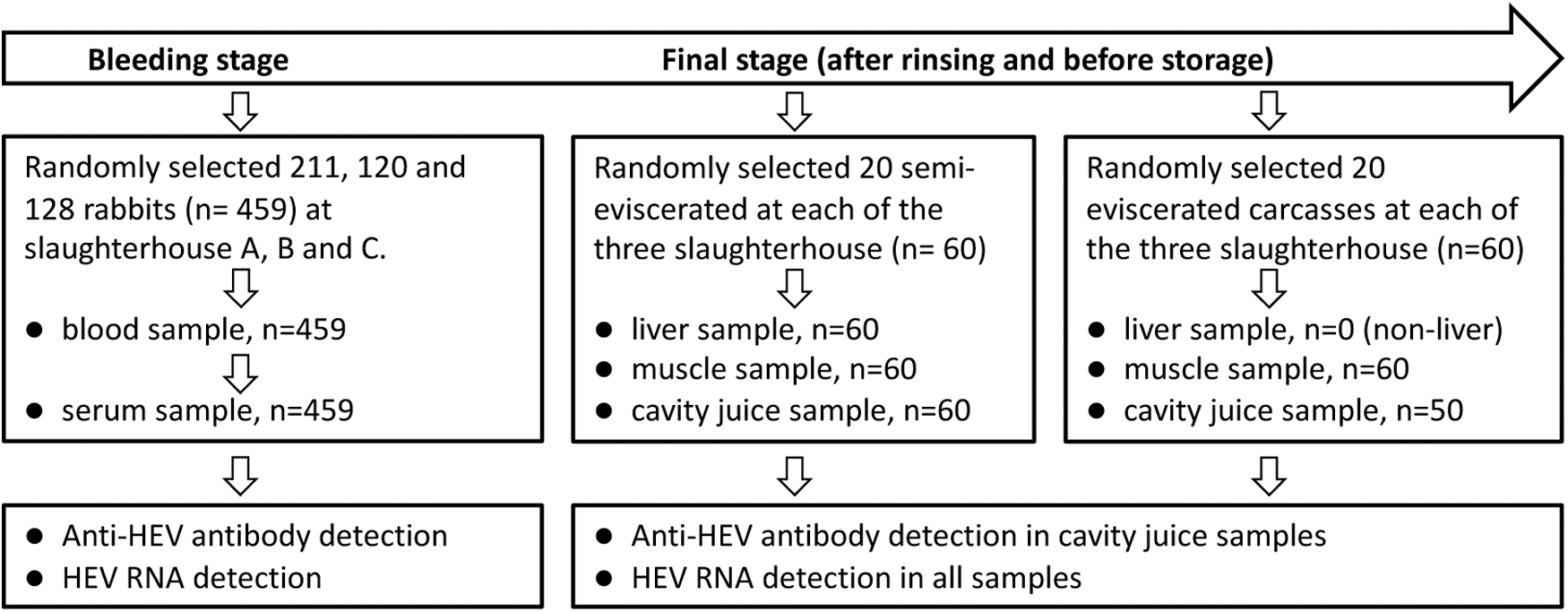
Diagram of sample collection at slaughterhouses during the process of rabbit slaughtering. HEV, hepatitis E virus.
Blood samples were drawn from bleeding rabbits suspended on a moving steel rope. One rabbit was selected at a random interval of 4–5 rabbits. A sterile tube was used to collect 5–6 mL of blood from each individual rabbit. Finally, a total of 459 blood samples were collected from three slaughterhouses.
After washing and before freezing for storage, 20 eviscerated and 20 semi-eviscerated carcasses from each of the three slaughterhouses were randomly selected for muscle, liver, and cavity juice sampling. The liver, ∼25 g of muscle, and at least 1 mL of cavity juice were collected simultaneously from each of the 60 semi-eviscerated carcass, whereas only muscle and cavity juice were collected from eviscerated carcasses. Muscle samples were collected from the carcass abdomen along both sides of the open section. Cavity juice, the residual liquid in cavities, was collected from the carcass's chest or abdominal cavity using disposable sterile tips. Because 10 of the 60 eviscerated carcasses did not have enough cavity juice, only 50 such samples were collected (Fig. 1).
The samples were immediately transported to the testing laboratory in cold boxes with ice after collection. In the laboratory, blood samples were centrifuged at 1000 × g, 10 min for serum collection. Whole livers and ∼25 g of muscles were cut and sliced; ∼2 g slices of each sample were collected at random and soaked in RNAlater™ Stabilization Solution (Invitrogen). The samples of rabbit sera, meat, livers, and cavity juice were stored at −80°C until further use.
Enzyme-linked immunosorbent assay for anti-HEV antibody
The sera were tested for anti-HEV antibody using a HEV-Ab enzyme-linked immunosorbent assay (ELISA) Kit (Wantai Biopharmaceutical, Beijing, China). The sandwich ELISA Kit is based on recombinant open reading frame 2 (ORF2) antigens that allow the detection of all antibody classes (immunoglobulin G, immunoglobulin M, and immunoglobulin A) of HEV-1–4 (Zhao and Wang, 2016). The performing procedures were in accordance with the manufacturer's instruction as described in previous studies (Li et al., 2021; Wang et al., 2021).
RNA extraction and reverse transcription-nested PCR for HEV RNA detection
Approximately 200 mg of each liver or muscle sample was homogenized in 1 mL tissue lysis buffer supplied in the TRIzol Up plus RNA Extraction Kit (TransGen Biotech, Beijing, China), and clarified by centrifugation at 12,000 × g for 15 min at 4°C. The supernatants were re-centrifuged. The second supernatants were collected for RNA extraction. Viral RNA was extracted from 200 μL serum, cavity juice or supernatants of tissue homogenates and recovered in 40 μL of elution buffer using TRIzol Up plus RNA Kit (TransGen Biotech) according to the manufacturer's instructions. The RNA extracts were used for complementary DNA (cDNA) synthesis immediately or stored at −80°C.
Aliquots of 8 μL of the RNA extracts were used to perform reverse transcription in a volume of 20 μL reaction mixture of TransScript First-Strand cDNA Synthesis SuperMix (TransGen Biotech) with oligo (dT) as the primer. A 10 μL volume of the cDNA product was used for nested PCR. A set of in-house primers were used to amplify a fragment of 348 nt in the ORF2 region of HEV RNA. External primers: SEBO2 (5′-TCY AAT TAY GCC CAR TAY CGG GTT G-3′), SEEO2 (5′-CCC TTR TCY TGC TGY GCR TTC TC-3′); internal primers: SEBI2 (5′-GTY ATG CTY TGY ATC CAT GGC T-3′), SEEI2 (5′-AGC CGA CGA AAT YAA TTC TGT C-3′). This primer set detects HEV-1, -3, and -4, but is more specific for HEV-3ra (Geng et al., 2011b).
The sample processing, extraction, and the RT-nested RNA procedures were essentially the same as those described previously (Geng et al., 2019b) except that RNAlater Stabilization Solution was used to protect the viral RNA intact. To validate the procedure for HEV RNA detection and exclude the possible inhibitory effect of the samples, samples detected negative for HEV RNA were further used for the following experiments: 50 μL of 104 genome equivalents (GE)/mL in phosphate-buffered saline HEV-3ra supernatants prepared from rabbit feces were injected into 1 g liver and 1 g muscle samples, respectively, and 5 μL of the 104 GE/mL virus supernatant was spiked into 0.5 mL of the serum and cavity juice, respectively.
Then, RNA isolation and reverse transcription nested PCR (RT-nested PCR) were performed on the prepared samples. The spiked rabbit HEV could be consistently detected in all the tested samples, indicating that no sample had an inhibitory effect, and the sensitivity of the assay for serum, cavity juice or tissue supernatant was at least 100 GE/mL.
Sequencing and phylogenetic analysis
The final amplicons of RT-nested PCR were purified and bidirectionally sequenced with the internal PCR primers commercially produced by Beijing Genomics institution (Beijing, China). After removing the primer sequences from each end, the final 304 nt sequences were aligned with the corresponding regions of reference sequences (Kaiser et al., 2018; Smith et al., 2020) available in GenBank. Phylogenetic analysis was performed with a neighbor-joining algorithm (Molecular Evolutionary Genetics Analysis, MEGA-X).
Statistical analysis
Data were analyzed using SPSS 22.0 (SPSS, Inc., Chicago, IL). The chi-square test was used to compare the prevalence of anti-HEV antibody, HEV RNA-positive rates between samples of different types. Results having p values <0.05 were considered significant.
Results
Seroprevalence of HEV among rabbits from slaughterhouses
Of the 459 serum samples collected from the three slaughterhouses, 50 sera were positive for HEV-Ab, resulting in an overall seroprevalence of 10.9% [95% confidence interval (CI): 8.1–13.7]. Seropositive animals were found in all three slaughterhouses, with varied positive rates ranging from 2.8% (95% CI: 0.6–5.1) to 20.8% (95% CI: 13.5–28.2) (Table 1).
Anti-Hepatitis E Virus Antibody and Hepatitis E Virus RNA Detection in Sera of Slaughtered Rabbits
CI, confidence interval; HEV, hepatitis E virus.
Anti-HEV antibody detection was performed on all 110 cavity juice samples collected from the cavity of rabbit carcass, and no cavity juice sample tested positive.
HEV RNA detection in rabbit blood, liver, muscle, and cavity juice
All serum samples were subjected to HEV RNA detection. Of the total 459 samples, 17 were positive, indicated that 3.7% (95% CI: 2.0–5.4) of the rabbits from the slaughterhouses were viremic at the time of slaughter. The percentage of HEV-viremic rabbits varied among slaughterhouses, ranging from 0.9% (95% CI: 0–2.3) to 7.5% (95% CI: 2.7–12.3) (Table 1).
Of the 60 livers collected from the rinsed semi-eviscerated rabbit carcass, 11.7% (95% CI: 3.3–20) were positive for HEV RNA (Table 2). Of the 120 muscle samples, none was positive for HEV RNA. Of the 110 cavity juice samples collected from the cavity of rabbit carcass, 2 samples (1.8%, 2/110) were detected positive for HEV RNA. In comparison with blood samples, the prevalence of HEV RNA was statistically higher in liver (p < 0.01), lower in muscle (p < 0.01), and there was no difference between blood and cavity juice (p > 0.05) (Table 2).
Hepatitis E Virus RNA Detection in Serum, Liver, Cavity Juice and Muscle of Slaughtered Rabbits
The two positive cavity juice samples came from two liver-positive semi-eviscerated carcasses. Because the eviscerated carcasses did not contain any organs, nonliver samples were taken from the semi-eviscerated carcasses. In terms of carcass type, of the 60 semi-eviscerated carcass, 7 (11.7%, 95% CI: 3.3–20.0) were HEV RNA positive, whereas none of the eviscerated carcass tested positive. The difference in HEV RNA-positive rates between these two types of carcasses was statistically significant (p < 0.05) (Table 3).
Comparison of the Hepatitis E Virus RNA Detecting Results Between Semi-Eviscerated and Eviscerated Carcasses
Genetic analysis of HEV strains
The final products of RT-nested PCR were sequenced. Of sequences obtained from the 348 nt PCR amplicons after removal of the primer sequences at each end, 23 were unique, whereas the other three were repeated. The 23 novel sequences have been deposited in the GenBank databases under accession numbers MW916509 to MW916531. Sequence alignment showed that the 23 novel sequences had 84.5–98% nucleotide sequence identity. Phylogenetic analysis showed that all the newly obtained sequences clustered within HEV-3, in the clade of the known rabbit HEV strains (Fig. 2).

Phylogenetic tree of the HEV strains from rabbits at slaughterhouses in Hebei Province of China. The tree was constructed based on a partial nucleotide sequence of the open reading frame 2 region with reported HEV sequences in GenBank as references. Filled circles indicate strains isolated in the current study. Reference sequences are labeled with the GenBank accession number/host/region/subgenotype (or none). The tree was constructed using the neighbor-joining method with MEGA-X.
Discussion
The positive rates of anti-HEV antibody among samples from three slaughterhouses in Hebei Province, China, ranged from 2.8% to 20.8% (Table 1). The average seroprevalence of HEV infection was 10.9% in slaughtered rabbits aged 3–6 months. These results are consistent with the previously reported seroprevalence of HEV infection (range 0–57%) in rabbits from rabbit farms in China (Geng et al., 2011b; Han et al., 2014; Zhao et al., 2009), and in agreement with reports from a number of other countries describing the seroprevalence of rabbits from farms or slaughterhouses (Bigoraj et al., 2020; Cossaboom et al., 2012; Lhomme et al., 2013).
Rabbits, like other food animals, are subjected to a preslaughter examination to screen for infectious diseases or infections caused by zoonotic pathogens. However, because HEV infection in rabbits usually manifests as an asymptomatic infection (Li et al., 2021), it is unlikely for a rabbit to be rejected for HEV infection. In the study, the average positive rate of HEV RNA in serum samples was 3.7%, ranged from 0.9% to 7.5% across the three slaughterhouses investigated. This result is also in accordance with previous studies on rabbits in farms (Geng et al., 2011b; Sooryanarain et al., 2020), and provides further evidence that HEV-infected rabbits can enter the slaughterhouses for rabbit meat production.
All rabbits sampled in this study were 3–6 months old, the regular slaughter age. It is known that the prevalence of HEV infection in farmed animals is affected by their age. Studies on the dynamic of HEV spread in pigs from birth to slaughtering showed that active HEV infection occurs naturally in most pigs around the age of 2 months; the highest prevalence of HEV was in pigs around the age of 3–4 months; most pigs at slaughter age (5–6 months) have experienced HEV infection and viremia has disappeared, indicating an obvious decrease in the rate of infection in slaughtered pigs and pork products (Capai et al., 2019; Feng et al., 2011). In contrast to pigs, the general seroprevalence was lower in rabbits at slaughter age, whereas the positive rate of HEV RNA was not necessarily lower. Since when only a small proportion of rabbits have experienced infection, a relatively large proportion could be in viremia or susceptible to HEV at slaughter. In addition, some rabbits showed a prolonged viremia period after HEV infection (Liang et al., 2022). The HEV RNA-positive rate of 0.9–7.5% in the sera of these rabbits was comparable with that in pigs at slaughter age (Capai et al., 2019; Feng et al., 2011).
Both the seroprevalence and the positive rate of HEV RNA in rabbits were different among the slaughterhouses (Table 1). In Hebei Province, slaughterhouses are licensed and administrated under “Hebei Province Regulations on Livestock and Poultry Slaughtering,” which makes the slaughtering procedures of the three slaughterhouses similar. However, the sources of the rabbits entering the slaughterhouses were different, as the slaughterhouses generally purchased rabbits from nearby rabbit farms. Previous studies have shown that HEV seroprevalences tend to differ significantly among rabbits from different farms in the same geographic area, owing to differences in housing practices, hygienic conditions of farms, and age of rabbits (Cossaboom et al., 2012; Geng et al., 2011a; Wang et al., 2013). Thus, the difference in HEV prevalence among the three slaughterhouses is more likely because of rabbit sources rather than slaughtering procedures.
The presence of HEV viremic rabbits during slaughter has been identified as a risk factor for the increased HEV seroprevalence among slaughter workers (Geng et al., 2019a), and could serve as an entry point into the food supply chain (Sooryanarain et al., 2020). In general, the slaughtered rabbits are processed into eviscerated or semi-eviscerated carcasses, both of which are ready-to-cook rabbit meat supplied to the market. We randomly selected eviscerated and semi-eviscerated carcasses to collect liver, muscle, and cavity juice for HEV detection (Fig. 1). HEV RNA was detected in the liver and cavity juice samples, confirming that rabbit meat with HEV could enter the food supply chain.
Of note, HEV RNA was detected in 11.7% (7/60) of the liver samples, but none of the 120 muscle samples tested had detectable HEV RNA (Table 2). The HEV RNA positive rate in rabbit livers was also statistically higher than in blood samples. This result is very similar to that of previous studies in pigs (Feurer et al., 2018; Geng et al., 2019b; Pallerla et al., 2021) and rabbits (Bigoraj et al., 2020). Having a higher HEV-positive rate in the livers than in other tissues seems to be common in HEV animal reservoirs, possibly because the liver is the primary target of HEV and the main site of HEV replication. Thus, the consumption of animal livers may pose a higher risk for HEV transmission.
HEV RNA was found in two cavity juice samples. During sample collection, we observed that there was more residual fluid in the cavity of semi-eviscerated carcasses than eviscerated carcasses. Both HEV RNA-positive cavity juice samples came from semi-eviscerated carcasses, and the livers of these two carcasses also tested positive for HEV RNA. Thus, HEV RNA was found in seven semi-eviscerated carcasses but not in any of the eviscerated carcasses, demonstrating a substantial difference between the two carcass types (Table 3). These observations indicate that the possibility of HEV in the eviscerated carcass is significantly reduced. The results may suggest that the organs should be removed from rabbit carcass before they enter the food chain. Consumers should thoroughly cook rabbit meat before eating it, and rabbit livers should be prepared with more care.
Consistent with previous studies conducted in China, all detected HEV sequences from the rabbit slaughterhouses in this study belong to HEV-3ra. Unlike swine HEV, for which the correlation between pork consumption and the zoonotic transmission of HEV has been established (Pavio et al., 2017), the role of HEV-3ra in causing clinical cases of hepatitis E has been unclear until recently, when several HEV-3ra strains were found in humans (Abravanel et al., 2017; Horvatits et al., 2021; Kaiser et al., 2018; Rivero-Juarez et al., 2020). However, until now, no rabbit HEV has been found in humans in China, where HEV-4 is the dominant genotype that causes human infection (Tang et al., 2023; Wang et al., 2016). The rarity of HEV-3ra infections in humans may be because rabbit products are consumed less frequently than pork products (Abravanel et al., 2017; Kaiser et al., 2018).
In this study, the three slaughterhouses were not randomly selected, but were those currently operating and permitting sample collection for the research. This limitation may result in sampling bias. Nonetheless, the study confirmed the possibility of raw rabbit meat with rabbit HEV entering the food chain. The potential risk of HEV transmission through consumption of rabbit products should be considered.
Footnotes
Authors' Contributions
Y.G. and C.Z. designed the experiment, analyzed the data and wrote the article. H.Z., X.L., C.W., and T.S. performed the experiment. C.W., X.L., G.Y., and H.Z. investigated the slaughterhouses and collected samples. All authors have read and approved the final version of the article.
Author Disclosure Statement
The authors declare no conflict of interest.
Funding Information
This research was funded by the Natural Science Foundation of Hebei Province, China (Grant No. H2021201015).