
Abstract
Background:
Over 200 different animal species are susceptible to infection by tularemia, highly infectious disease caused by Francisella tularensis. Hares (Lepus sp.) and small rodents, such as common vole (Microtus arvalis), have been acknowledged as the most significant sources of human tularemia infection in most European countries. Our objective was to verify ability of these species in predicting incidence of human tularemia in a Central European country, the Czech Republic.
Materials and Methods:
We used 17 years of data on densities of European hare (Lepus europaeus) and common vole, and climate variability to test effects of these factors on temporal dynamics of tularemia incidence. The data were obtained from annual reports available from online e-repositories.
Results:
The analysis showed that 33% of the yearly variation in human tularemia incidence was explained solely by the abundance of European hare in the Czech Republic during 2007–2022. Density of common vole and North Atlantic Oscillation index, a measure of climate variability, did not significantly explain tularemia incidence. While hare population declined severely during 1993–2022, we did not detect any clear accompanied decrease in the prevalence of tularemia in humans and hares.
Conclusion:
Contrary to expectations, only hares proved capable in predicting yearly dynamics in human tularemia incidence in the Czech Republic. We call for continued monitoring of infection rates in hares and advocate the use of hunter estimates of hare abundance as a cheap and effective means of predicting the risk of tularemia.
Introduction
In Europe, human tularemia incidence is highly variable in both space and time. Over 200 different animal species are susceptible to the disease (Colville and Berryhill, 2007). Among these, hares (Lepus sp.) have been acknowledged as the most significant pool of human tularemia infection in most European countries (Maurin and Gyuranecz, 2016). Rodents, such as common vole (Microtus arvalis), also constitute a significant source of infection, particularly in areas of high vole biomass and pronounced outbreak dynamics as in Scandinavia (Hestvik et al., 2015) and central Spain (Luque-Larena et al., 2017). Vole is an important driver of other tick-borne diseases such as Lyme disease and tick-borne encephalitis (Tkadlec et al., 2019).
Our understanding of tularemia epidemiology comes largely from studies of prevalence and ways of transmission (Hestvik et al., 2015), spatiotemporal distribution (Desvars et al., 2015; Eliasson et al., 2002; Pikula et al., 2004), clinical aspects (Maurin and Gyuranecz, 2016), and bacteriological studies on Francisella tularensis, the cause of tularemia (Macela et al., 2012). Less effort has been devoted to understanding direct effects of hare and rodent populations on tularemia prevalence (Luque-Larena et al., 2017; Mörner, 1992; Rodríguez-Pastor et al., 2017).
We tested whether reported relationships between the incidence of human tularemia and hare and vole abundance reported from Sweden (Mörner, 1992) and Spain (Luque-Larena et al., 2017; Rodríguez-Pastor et al., 2017), respectively, hold for the Central European country, the Czech Republic. There, both species belong to key elements of the herbivore community of the agroecosystems. We further analyzed interannual trends in tularemia incidence in humans and hares and trends in hare density and harvest size to verify congruence in trends among these variables.
Materials and Methods
We used 17 years of data on European hare (Lepus europaeus), common vole, and climate variability to test whether any of these factors may predict the incidence of tularemia in the Czech Republic. Yearly tularemia incidence was obtained from publicly available reports annually published by The National Institute of Public Health in the Bulletin of Centre for Epidemiology and Microbiology (
We used late winter hunter counts of hares as an approximation of hare abundance. Every year, hunters estimate the population size of hares by counting all seen individuals, while walking through hunting territories on a given day in February or March. These estimates provide valuable information in wildlife management (Hušek et al., 2021). Hunter estimates were obtained from yearly hunters' reports (
We obtained indices of vole density in common crops (alfalfa, clover, grassland, wheat, barley, and rape) in October-November for the years 2006–2022 from the Central Institute for Supervising and Testing in Agriculture, which organizes annual counts of active burrow entrances per hectare in crop fields during walks along four strips of 100 m length and 2.5 m width at ∼10 sites in each district (for further details, see Tkadlec et al., 2019). These counts are performed twice a year, in spring and autumn. We preferred the autumn data following Tkadlec et al. (2019). Indices were downloaded from
Before analysis, we Box-Cox transformed the data to ensure stationarity using package MASS (Venables and Ripley, 2002). Parameter λ was estimated from the data. We removed temporal trends in time series data by fitting smoothing splines with function
We used the likelihood ratio to test the presence of temporal autocorrelation in incidence data by comparing a null model (intercept only) with a model accounting for the autoregressive process of order 1 (AR1) using package nlme (Pinheiro et al., 2022). A model accounting for (AR1) was not supported over the null model (likelihood ratio = 0.09, p = 0.77). Hence, we analyzed the effects of explanatory variables on tularemia incidence with ordinary least square linear regression. We adopted the forward selection, keeping only statistically significant explanatory variables in the model. As explanatory variables, we considered hare and vole density in year t and t-1 and winter NAO. Assumptions of fitted linear regression models were validated using package gvlma (Penna and Slate, 2019).
Finally, we used 30 years of data (1993–2022) on human tularemia cases, hare abundance, and annual harvest size of hares and 8 years of data (2011–2018) on tularemia infection rate in hares to evaluate congruence in long-term trends among these variables. Infection rates in hares were monitored annually during the study years by the State Veterinary Administration, by testing a prescribed number of randomly collected hares in each of the 76 districts. We obtained tularemia incidence rates in hares from annual reports (
Results
We found that only the effect of hare density t-1 on tularemia incidence was significant over the period 2007–2022 (β ± standard error = 301.4 ± 114.8, t = 2.63, p = 0.02, R 2 = 0.33; Fig. 1a, b), but not the effect of hare density t (t = −1.10, p = 0.29), voles t (t = −0.33, p = 0.75), voles t-1 (t = −1.11, p = 0.29), or winter NAO (t = 0.97, p = 0.35). While hare population size and annual harvest size declined severely during 1993–2022 (Fig. 2a, b), we did not detect any clear trend in the prevalence of tularemia in hares and humans (Fig. 2c, d).

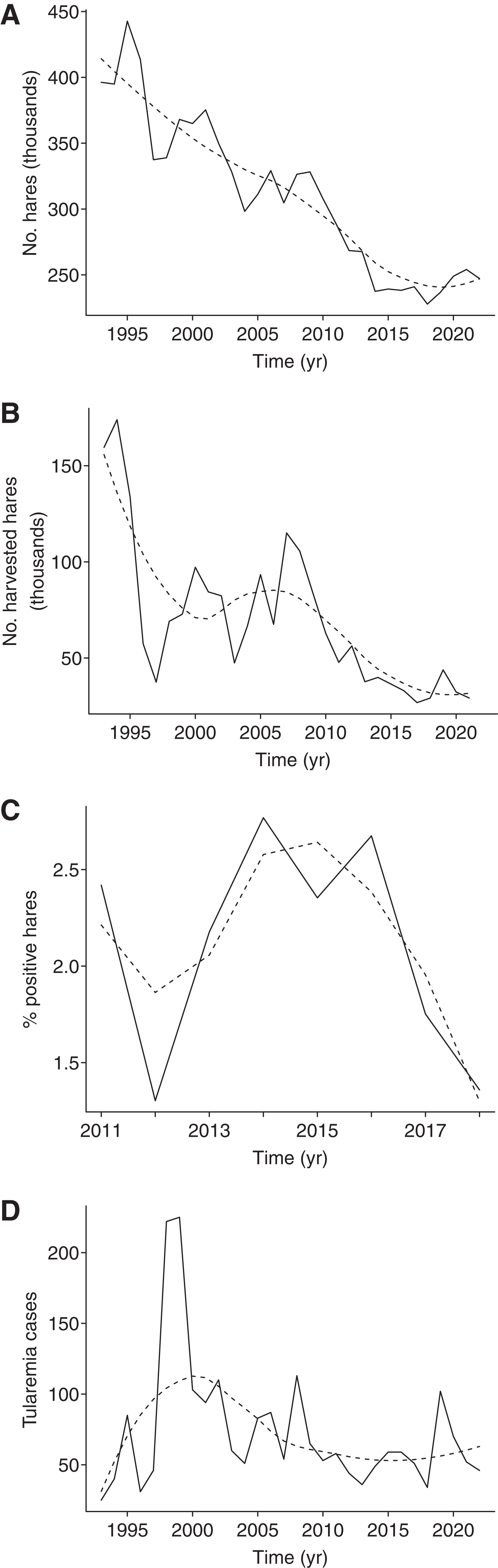
Trends in original time series (dashed line) of
Discussion
We demonstrated clear support for the positive effect of hare density in the previous late winter on tularemia incidence. The 1-year lag in the hare effect suggests, that there is an age effect of hares on the risk of infecting humans by tularemia. Perhaps, it is mainly adult hares that become agents of infection, either because of a higher probability of direct contact (e.g., with hunters during the autumn hunting season) or because of the lower survival of yearling hares, preventing them from effectively spreading infection. We found that hare density explained 33% of the variation in tularemia incidence, which corresponds well to hares being identified as a source of 30% of human infections in the Czech Republic during 1993–2019 (Liptáková et al., 2020).
We propose that 67% of the variability in human tularemia incidence not explained by hare density may be attributed to other effects such as the infection rate of hares, use of protective measures (e.g., gloves) when handling hares, and encounter rate of other species of infected animals and tularemia vectors (mainly ticks). Although we failed to detect any significant effect, either direct or delayed, of vole density on tularemia incidence, we argue that voles may play an important role as an infection reservoir in the Czech Republic.
Admittedly, 17 years of national scale annual data used in our study may not be sufficient for the detection of a statistically significant effect of voles. Also, the vole burrow index may not properly reflect true vole densities, especially when vole numbers are very low or very high (Lisická et al., 2007). The importance of common voles in dynamics of human tularemia in the Czech Republic has been supported by the finding of voles causing 13% of human infections during the period 1993–2019 (Liptáková et al., 2020).
We found that, although hares and their harvest declined dramatically during 1993–2022, the incidence of human and hare tularemia did not show any clear corresponding decrease, as also indicated by a previous study (Hestvik et al., 2015). To better understand these factors, we call for continued monitoring of infection rates in hares and advocate the use of hunter estimates in predicting the risk of tularemia in the Czech Republic.
Footnotes
Acknowledgments
We thank an anonymous reviewer for comments that improved clarity of the article.
Authors' Contributions
Both authors conceived the idea. Jan H. analyzed the data and wrote the article. Both authors provided comments on the text and approved the release for publication.
Author Disclosure Statement
No conflicting financial interests exist.
Funding Information
This work was financially supported by Ministry of Culture of the Czech Republic (DKRVO 2019–2023/6.VIII.e, National Museum, 00023272).