
Abstract
Background:
Jingmen tick virus (JMTV) and Tacheng tick virus-1 (TcTV-1) are emerging tick–borne viruses that have been recently confirmed to be etiological agents of human disease in China. However, the ecology of JMTV and TcTV-1, especially their association with ticks in wildlife and livestock, remains largely unknown in Turkey.
Materials and Methods:
Eight hundred thirty-two tick specimens in 117 pools were collected in Turkey between 2020 and 2022 from wildlife (Miniopterus schreibersii and Rhinolophus hipposideros; n = 10, 1.2%; Testudo graeca; n = 50, 6%) and livestock (Ovis aries and Capra aegagrus hircus; n = 772, 92.7%). The specimens were individually screened for JMTV and TcTV-1 using nRT-PCR assays targeting the partial genes.
Results:
JMTV was detected in one Ixodes simplex pool and two Rhipicephalus bursa pools collected from central and Aegean provinces, respectively. TcTV-1 was identified in five Hyalomma aegyptium pools collected in Mediterranean provinces. No coinfection was detected in the tick pools. Maximum likelihood analysis of JMTV partial segment 1 sequences reveal that these sequences form a separate cluster together with viruses previously characterized in Turkey and the Balkan Peninsula. The phylogenetic analysis of the TcTV-1 nucleocapsid sequences indicates that they are closely related to viruses in ticks, sheep, cattle, and humans in China, but form a separate group among themselves.
Conclusion:
This study provides the first molecular evidence of TcTV-1 in Hy. aegyptium in Turkey. In addion, these findings indicate that JMTV and TcTV-1 extend ticks species and geographic distributions. Thus, multiregional surveillance in livestock and wildlife is needed to evaluate potential tick vectors and the human health impact of these viruses in Turkey.
Indroduction
Human and animal health are increasingly threatened by emerging and/or resurgent viral, bacterial, and protozoan pathogens, with two new pathogens appearing annually that only affect human populations (Kapuscinski et al., 2021). Most such pathogens are transmitted through blood-sucking insects such as ticks, mosquitoes, black flies, and bed bugs. Ticks, which are obligate ectoparasites in the class Arachnida, are the second-most common human and animal pathogen vector after mosquitoes. Climate change, contact with vertebrate hosts, and ecological and anthropogenically induced activities have expanded the geographic distribution of ticks and tick-borne pathogens (Socha et al., 2022).
Recently, an increasing number of new tick borne viruses (TBVs) have been found worldwide, such as severe fever with thrombocytopenia syndrome virus, Bourbon virus, Dugbe virus, Heartland virus, Crimean-Congo Heamorrhagic fever virus (CCHFV), and Nairobi sheep disease virus (Dinçer et al., 2019; Ergünay et al., 2020; Socha et al., 2022). Many of these viruses can cause mild to severe symptoms and even death in humans, livestock, and wildlife (Bartíková et al., 2017).
Jingmen tick virus (JMTV), a newly identified tick-borne, segmented RNA virus, was first described in ticks in the Jingmen region of Hubei province in China in 2010 (Qin et al., 2014). JMTV has a linear, positive-sense, single-stranded RNA genome of four separate segments (S1, S2, S3, and S4). S1 and S3 encode nonstructural proteins (NSP1 and NSP2) that are genetically related to nonstructural proteins (NS3 and NS5) of classic flaviviruses of the family Flaviviridea, whereas S2 and S4 have no homology with the sequences of any known viruses (Qin et al., 2014; Shi et al., 2015).
Since its first documentation in China in 2010 (Qin et al., 2014), JMTV has been reported in specimens from several countries, including China (Guo et al., 2020; Meng et al., 2019; Yu et al., 2020), North America, Brazil, French Antilles, Japan, UK, Trinidad and Tobago, and Kenya (Kobayashi et al., 2021; Maruyama et al., 2014; Ogola et al., 2022; Sameroff et al., 2019; Temmam et al., 2019). In addition to domestic animals such as cattle (Souza et al., 2018), JMTV has been characterized in wild animals, including monkeys, rodents, and bats (Guo et al., 2020; Maruyama et al., 2014; Shi et al., 2015; Yu et al., 2020). JMTV RNA has also been identified in primate samples, suggesting that it may infect humans as well (Ladner et al., 2016).
More importantly, JMTV has been detected in acute phase serum specimens of CCHFV-positive patients (Emmerich et al., 2018; Ternovoi et al., 2020) and from patients in China with tick bite histories using high-throughput sequencing of skin biopsies and blood samples (Jia et al., 2019; Taniguchi, 2019).
Two new JMTV-like viruses, Alongshan virus (ALSV) and Yanggou tick virus, have been found in febrile human patients in China (Wang et al., 2019). ALSV has also been detected in Ixodes persulcatus ticks, Culex tritaeniorhynchus, and Anopheles yatsushiroensis mosquitoes in China, Ixodes ricinus ticks in south-eastern Finland and France (Kuivanen et al., 2019), and in I. persulcatus ticks in the southern European Russia and north-west Russia (Kholodilov et al., 2020). In short, JMTV and JMTV-like viruses appear to have a worldwide geographic distribution.
Tick-borne orthonairoviruses, classified in the family Nairoviridae, have attracted increasing attention from global health organizations due to causing fatal illnesses in humans and animals (Garrison et al., 2020). Four viruses in this genus (CCHFV, Dugbe virus, Nairobi sheep disease virus, and Kasokero virus) have been reported as causing disease in humans. Tacheng tick virus-1 (TcTV-1) is a serotype of Tamdy orthonairovirus, which was first isolated in China from Dermacentor marginatus using metagenomic surveillance (Li et al., 2015). Other members of this serotype, which are detected in ticks and of largely unknown medical significance, include Burana virus, Huangpi tick virus 1, and Wenzhou tick virus (Maes et al., 2018). TcTV-1, with a genome organization similar to Bunyaviruses, has a negative-sense, single-stranded RNA genome encoding three segments: small (S), medium (M), and large (L).
TcTV-1 was first detected in 2019 in a patient with a history of tick bites. Viral RNA has also been detected in livestock (cattle, sheep) and ticks, including D. marginatus, Dermacentor silvarum, Dermacentor nuttallia, and Hyalomma asiaticum (Liu et al., 2020). In 2021, a tick bite patient in China was reported to have TcTV-1 and Ricketttia raoultii coinfection (Zhang et al., 2021). Finally, in addition to ticks, humans, and livestock, TcTV-1 has been identified in spleen samples of great gerbils (Rhombomys opimus) in China (Ji at al., 2023).
Turkey, located at the intersection of Europe, Asia, and Africa, has various climatic zones while bird migration routes have resulted in the distribution of various tick species and TBVs that infect human and animals (Bursalı et al., 2012). The most important tick-borne virus, CCHFV, was first detected in 2002 in Anatolia and Eastern Thrace. Since then, many studies have investigated CCHFV distribution and presence in animals, vector ticks, and humans (Leblebicioglu et al., 2016; Yesilbag et al., 2013). Research into other TBVs, such as Tamdy virus, JMTV, and Meram virus, recently described in ticks, seem to be overshadowed by CCHFV (Brinkmann et al., 2018; Dinçer et al., 2022; Dinçer et al., 2019; Ergünay et al., 2020). Consequently, potential tick vectors, reservoir animals, and these viruses' geographic distributions remain largely uncertain. Accordingly, we conducted molecular screening in ticks collected from wildlife (bat and tortoise) and livestock (sheep and goat) in several provinces of Turkey for the presence of two TBVs: JMTV and TcTV-1.
Materials and Methods
Tick collection and processing
Tick specimens were collected between 2020 and 2022 at six locations in Mersin, Antalya, and Osmaniye the Mediterranean region; Kırklareli in Thrace; Ankara in central Anatolia, and Izmir in the Aegean region (Table 1). Samples were collected using forceps from sheep (Ovis aries), goats (Capra aegagrus hircus), and tortoises (Testudo graeca), whereas bats were caught individually using hand nets and checked for attached ticks before being released immediately.
Locality of Collection Region, Collection Dates, Host and Tick Species
The samples for the study were field-collected and host-seeking, or detached from animals. Collection was conducted near farms, animal shelters, or human dwellings with the informed consent and cooperation of relevant breeders or caretakers. Neither local nor regional Ethics Committee approval was required in this setting. Before starting the study, permission for bat capture was provided by the Animal Research Ethics Committee of Hacettepe University (Permit No. 2019/66). No live bats were harmed during the study.
Since positive samples were to be inoculated in cell cultures, the tick samples were placed individually into eppendorf tubes without preservatives or alcohol and then transferred to the laboratory in dry ice. Morphological identification of ticks was performed using taxonomic keys (Estrada-Peña et al., 2017; Walker et al., 2020) before storage at −80°C until processing.
Tick pools of one to seven individuals were created based on collection area, host, species, and developmental stage. Each pool was homogenized using tungsten carbide or stainless steel beads (QIAgen, Hilden, Germany) in 200–500 μL of Dulbecco's phossphate-buffred saline, supplemented with 1%
Detection of JMTV and TcTV-1 in ticks
To detect JMTV and TcTV-1 presence, the tick pools were individually screened using two nested RT-PCRs, targeting the NS5-like protein or nucleoprotein on segment 1 and segment S, respectively, as described previously (Liu et al., 2020; Yu et al., 2020). First-round PCRs were as follows: initial 4 min denaturation at 94°C, followed by 40 cycles at 94°C for 45 s, 54°C for JMTV, and 53°C for TcTV-1 for 1 min, and 72°C at 1 min, with a final extension at 72°C for 10 min. Second-round PCRs followed the same conditions except for primer annealing (53°C for JMTV and 52°C for TcTV-1).
The products of the second-round PCRs (370 and 328 base pairs in length for JMTV and TcTV-1, respectively) were separated using 1% agarose gel electrophoresis stained with SYBR safe DNA gel stain (Thermo Fisher Scientific, Germany) and visualized in a ChemiDoc XRS+ imaging system (BioRad, Germany). DNase/RNase free water was included as a negative control. The nRT-PCRs and viral nucleic acid extractions were performed using biosafety level 2 class cabinets at various times and places.
Virus isolation
Virus isolation was conducted by inoculating positive tick homogenate onto semiconfluent monolayers of African green monkey kidney cells (Vero E6, ATCC: CRL-1586), provided from the cell culture collection of the Department of Virology, Faculty of Veterinary Medicine, Ataturk University. The filtered tick homogenate (500–600 μL) was inoculated onto cells in 25 cm2 flasks. After cell adsorption for an hour, 5 mL of Dulbecco's modified Eagle's medium, supplemented with 5% fetal bovine serum, l-glutamine, 100 U/mL penicillin, and 100 μg/mL streptomycin, was added. The cells were incubated in 5% CO2 atmosphere at 37°C and checked daily for cytopathic effects. Finally, blind passages were performed every week, and the culture fluids were tested for viral nucleic acids at each passage.
Sequences and phylogenetic analysis
The PCR products of expected size in each assay were purified using a commercial kit (GeneJET; Thermo Scientific, Lithuania) according to the manufacturer's instructions. Sequencing was performed in ABI PRISM 3500xL Dx Genetic Analyzer (Thermo Fisher Scientific, Germany) using the second-round PCR primers. Raw sequences were analyzed using Geneious bioinformatic software (Version 11.1.5; Biomatters Lts., Auckland, New Zealand). To search for similarity, the sequences were compared to other JMTV and TcTV-1 sequences available from GenBank Database (
Results
Tick collection and processing
During 2020–2022, 832 ticks were collected from the following provinces in Turkey: Kırklareli, Mersin, and Ankara (l. simplex and I. vespertilionis, n = 10, 1.2%), Osmaniye (T. graeca; Hy. aegyptium, n = 50, 6%); Izmir and Antalya (O. aries and C. aegagrus hircus; R. bursa n = 700, 84%, D. marginatus n = 70, 8.4%, and H. parva n = 2, 0.24%) (Table 1). The most common tick species detected were R. bursa, followed by D. marginatus, Hy. eagyptium, and others.
A total of 10 tick specimens from bats were collected: 7 from Lömbürdekini Cave in Ankara province, 2 from Sinobiç Cave in Mersin province, and 1 from Koyunbaba Cave in Kırklareli province. Of which, seven adults and two nymphs specimens were obtained from Miniopterus schreibersii species while one larva was Rhinolophus hipposideros species. Ticks collected from tortoises comprised adults (n = 33), nymphs (n = 12), and larvae (n = 5). In addition, tick specimens from sheep and goats were identified as adults (n = 652), nymphs (n = 65), and larvea (n = 55) (Table 1).
JMTV and TcTV-1 detection in ticks
To detect JMTV and TcTV-1 viral RNAs, the tick pools were individually screened using two nRT-PCRs, targeting NS5-like proteins or nucleoproteins on the segment 1 and segment S, respectively, as described previously (Liu et al., 2020; Yu et al., 2020). Minimum infection rate was detected with 3.6% and 6% for JMTV and TcTV-1, respectively. Positive specimens came from the following provinces: Izmir (pool codes: 10 and 23; 25%), Ankara (pool code: 54; 12.5%), and Osmaniye (pool codes: 57, 59, 61, 63, and 66; 62.5%) (Table 2). These pools included various tick species, including I. simplex (n = 1, 12.5%), R. bursa (n = 2, 25%), and Hy. aegyptium (n = 5, 62.5%). No coinfection was detected in any tick pool.
Tick Pools with Detectable Jingmen Tick Virus and Tacheng Tick Virus-1
JMTV, Jingmen tick virus; TcTV-1, Tacheng tick virus 1.
Two R. bursa pools (10 and 23) from Izmir province and one I. simplex pool (54) from Ankara province tested positive for JMTV. Pairwise comparisons demostrated 86.0–99.0% nucleotide identities with JMTV isolates previously found in Turkey, while the pairwise comparisons among the three JMTV isolates was 95.0–98%. Phylogenetic analysis showed that the three JMTV sequences formed a single group, together with viruses previously characterized in Turkey (Fig. 1). Unfortunately, it was not possible to isolate JMTV in the current study.

Maximum likelihood analysis of the JMTV partial segment 1 coding sequences (338 bp). Tree is constructed using Tamura 3-parameter model for 500 replications. Global viruses are represented by GenBank accession number, abbreviation, isolate/strain identifier, host/tick species, and country of origin. In this study, JMTV sequence obtained from Ixodes simplex (bat tick) is shown with black circle and others are shown with black diamond. YGTV and ALSV are included as outgroups. ALSV, Alongshan virus; JMTV, Jingmen tick tick virus; YGTV, Yanggou tick virus.
Finally, TcTV-1 sequences were found in five pools (57, 59, 60, 63, and 66) of female Hy. aegyptium ticks from T. graeca in Osmaniye province. The TcTV-1 sequences in the current study had 96.0–98% nucleotide identity among themselves and slightly lower similarity (86.0–99.0%) to samples previously detected in China. The phylogenetic tree (Fig. 2) based on the partial S segment indicated that the Turkish TcTV-1 sequences formed a separate group together with other strains previously characterized in ticks, sheep, cattle, and tick-bitten patients in China.
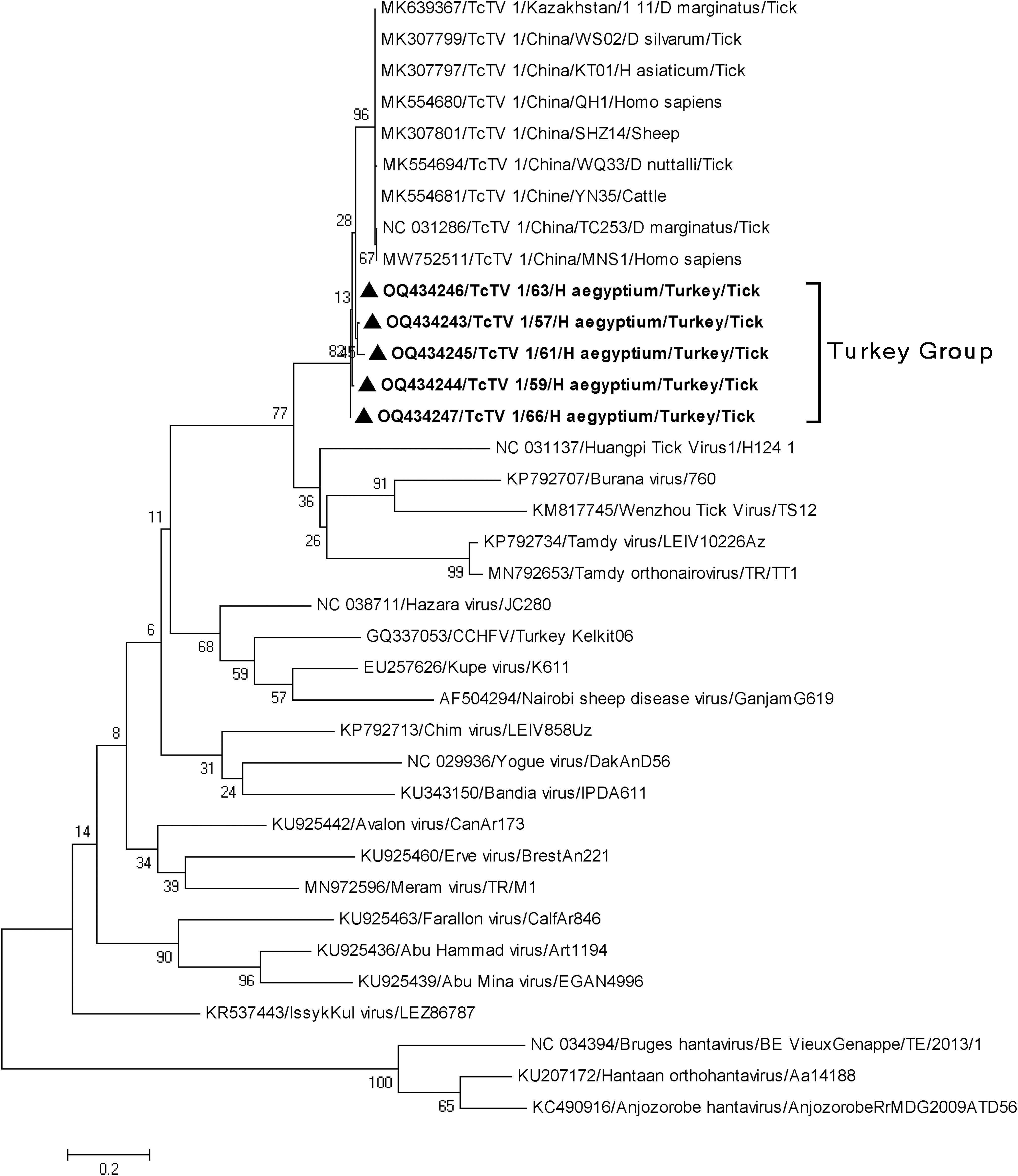
Maximum likelihood analysis of the TcTV-1 partial S segment coding sequences (285 bp). Tree is constructed using Tamura 3-parameter model for 500 replications. Global viruses are represented by GenBank accession number, abbreviation, isolate/strain identifier, host/tick species, and country of origin. In this study, TcTV-1 sequences are shown with black triangle. Hantaviruses are included as outgroups. TcTV-1, Tacheng tick virus-1.
Discussion
In this study, we investigated the presence of TBVs, recently identified as human pathogens, in ticks obtained from livestock and wildlife in Turkey. A total of 832 ticks from 6 species were removed from livestock (O. aries and C. aegagrus hircus) and wildlife (T. Gracea, M. schreibersii, and R. hipposideros) from five provinces in four geographic regions of Turkey. The tick pools were individually screened for presence of JMTV and TcTV-1 using nRT-PCRs. Viral RNA detection was performed in 8 (6.8%) tick pools. Sequence analyses of the PCR products revealed JMTV and TcTV-1 (Table 2).
JMTV- and JMTV-like viruses have recently been detected in many countries, including China, Romania, Japan, Turkey, Uganda, Kenya, and Brazil (Dinçer et al., 2019, Emmerich et al., 2018; Kobayashi et al., 2021; Maruyama et al., 2014; Ogola et al., 2022; Sameroff et al., 2019; Temmam et al., 2019; Zhang et al., 2022). In addition to various arthropod vectors, such as ticks and mosquitoes, JMTV has been found in a broad range of animal hosts, including cattle, sheep, goats, tortoise, rodents, primates, and recently bats (Guo et al., 2020; Ogola et al., 2022; Pascoal et al., 2019; Qin et al., 2014; Temmam et al., 2019; Yu et al., 2020).
Bats (order Chiroptera) are the second largest mammal group, with over 1240 species. Through their ectoparasites, mainly ticks, bats can transmit highly fatal and infectious viruses affecting both human and animal health, such as Issyk-Kul virus, Hendra virus, Nipah virus, and Ganjam virus (Brinkmann et al., 2020). More recently, bats have been identified as a reservoir for SARS-CoV-2, which caused the Covid-19 pandemic.
JMTV has been detected in blood and urine samples of various bat species (Guo et al., 2020; Temmam et al., 2019). Previously, we characterized JMTV in different provinces in Turkey from various tick species, including Rhipicephalus, Haemaphysalis (Dinçer at al., 2019) and Hyalomma spp. (Dinçer at al., 2022), gaining new data about JMTV prevalence in ticks collected from livestock. However, to our knowledge, our study provides the first report of JMTV in bat ticks in Turkey. We detected JMTV in I. simplex collected from M. schreibersii in central Anatolia (Ankara province). The JMTV sequences we obtained from wildlife and livestock were closely related to viruses previously detected from Turkey and formed a group (Fig. 1). Both our data and previous studies (Guo et al., 2020; Temmam et al., 2019) demostrate that both bat species and their ticks may be natural hosts for JMTV.
However, because the present study was conducted with limited tick samples collected from bats in several provinces, large screening studies are needed to gain more information about JMTV circulation and transmission in bats and their ticks. Finally, detection of JMTV in ticks collected from both bats and domestic animals indicates that JMTV is circulating in wildlife and spreading over a wider geography than previously known in Turkey. As mentioned above, bats were a key host for SARS-CoV-2, which has been the most critical global health issue in recent years. Therefore, detection of JMTV in ticks collected from bat hosts in Turkey indicates a very different situation from its previous detection in domestic animals.
Initially, JMTV isolation was performed using mammalian and insect cell lines, such as C6/36 and DH82 (Qin et al., 2014). However, virus growth was inadequate in both cases. Later studies demostrated JMTV proliferation and replication in an embryo-derived cell line BME/CTVM23, while the JMTV Anatolian strain was isolated and maintained in Vero E6 cells (Dinçer et al., 2019). In the present study, we were unable to isolate from JMTV due to insufficient positive pools. Other cell lines, such as Vero E6, C6/36 (Ogola et al., 2022), and Vero and BHK-21 (Zhang et al., 2022), have been tried for isolating JMTV, but these attempts were unsuccessful, possibly because of insufficient viable virus copies or use of unsuitable cell lines. To ensure successful isolation, future studies should determine cell tropism and optimal cell lines for JMTV.
TcTV-1 was first islolated and characterized in D. marginatus in China (Li et al., 2015). TcTV-1 has also been found in ticks, including D. marginatus, D. silvarum, D. nuttalli, Hy. asiaticum reticulate, and Rh. turanicus (Liu et al., 2020). TcTV-1 was also detected in a patient with tick-bite history showing clinical signs of fever, rash, chills, myalgia, and arthralgia. This suggests that TcTV-1 may cause a febrile illness syndrome in humans (Liu et al., 2020).
Tortoises, known to be longest-living reptile, are one of ticks' most preferred hosts. The spur-thighed tortoise, T. graeca, is a major tortoise species found in wild ecosystems in many regions, including North Africa, the Middle East, Asia Minor, and southeastern Europe (Graciá et al., 2017). As a popular household pet, they are directly collected from wildlife and traded in many countries, which facilitates transmission of tick-borne diseases to humans and other domestic animals (Norte et al., 2022). Ranaviruses and herpesviruses are known as common infections of tortoises. Recently, JMTV was detected in Kenya in tortoise blood and ticks, including Amblyomma sparsum and Am. nutalli (Ogola et al., 2022). While Hy. eagyptium occasionally feeds on humans and other species, it is mostly found in all life stages on tortoises (Kar et al., 2017).
Nairoviruses, such as CCHFV, TAMV, and Meram virus, have been chracterized in Hy. eagyptium collected from different provinces of Turkey (Brinkmann et al., 2018; Ergünay et al., 2020). This was the first study to detect and characterize TcTV-1 in Turkey, as a tick-related nairovirus other than CCHFV, TAMV, and Meram virus. In the present study, TcTV-1 was detected in Hy. eagyptium collected from T. greace in the Turkey's Mediterrean region (Osmaniye province). Phylogenetic analyses (Fig. 2) based on the partial S segment demostrated that TcTV-1 sequences formed a separate group that shares a common origin with viruses characterized in ticks, sheep, cattle, and tick-bitten patients in China.
To our best knowledge, TcTV-1 has not been previously reported outside of the Far East (China). While there is still very limited data on TcTV-1's global epidemiology, its detection in the present study in wildlife in Turkey, 6000 km from China, where it is endemic, demonstrates that the TcTV-1 is circulating across a much wider area than its known geography.
JMTV and TcTV-1 sequences were only detected in female tick specimens in this study. Due to the study plan and evaluation of the tick specimens in pools, origin of the these sequences could not be identified exactly and may be from environmental sources. JMTV and TcTV-1 partial sequences were found in ticks detached from wildlife and livestock animals, and in pools where blood-fed specimens were included. Thus, tick infested vertebrates may be the source in some or all detected viral sequences (Brinkmann et al., 2018; Ergunay et al., 2020).
In conclusion, our study provides significant molecular data on JMTV and TcTV-1 in several regions of Turkey. We provide the first report of JMTV in I. simplex collected from M. schreibersii and TcTV-1 in Hy. aegyptium collected from T. graeca. Considering their emergence in recent years, more in-depth epidemiological studies are needed to elucidate the reservoirs, enviromental determinants, and vectors of these viruses in wildlife and livestock. Given that these TBVs can cause febrile illness syndrome in humans, the clinical and scientific communities need to be prepared for a potential future epidemic.
Footnotes
Authors' Contributions
E.D.: conceptualization, validation, data curation, and writing—original draft; M.O.T.: investigation, formal analysis, writing, and methodology; O.H.: specimen collection; D.Y.: specimen collection; M.B.N.: laboratory assays; T.Z.S.: specimen collection and laboratory assays; and T.Y.: specimen collection. All authors have read, revised, and approved the article.
Ethical Approval
The tick specimens evaluated in the study comprised field-collected host-seeking or questing ticks and those removed from domestic or wild animals and cooperation of the caretakers or wildlife preservation experts. No local or regional Ethics Committee approval was required. Before starting the study, ethical approval for bat capture was provided by the Animal Research Ethics Committee of Hacettepe University under permits no 2019/66. No live bat was harmed for this study.
Author Disclosure Statement
No conflicting financial interests exist.
Funding Information
No funding was received for this article.