
Abstract
Background:
Developing methods for the isolation and replication of dengue virus (DENV), based on nonhematophagous insect models to assess virus–host interaction, would contribute, for instance, to the creation of drugs or vaccines and eventually to the control of the disease. In this regard, nonhematophagous mosquitoes have been used as biological hosts for the isolation of DENV because they are specific and sensitive to a low viral load and viral particles with low infectivity. However, implementation of these models is mainly affected by the complexity of the establishment of the entomological colonies.
Materials and Methods:
In this study, the susceptibility of DENV-4 infection in Plodia interpunctella larvae was evaluated. Ten larvae, previously inoculated with supernatant from DENV-4-infected C6/36 cells, were processed to determine viral replication by the optical density and 2−ΔΔCt methods at different time intervals (1 and 7 days postinoculation).
Results:
A prospective increase in viral replication was observed, which did not influence the survival and development of P. interpunctella.
Conclusion:
These results demonstrate the infectivity of DENV-4 in P. interpunctella, thus becoming an option as a biological model for the study of this etiological agent.
Introduction
Dengue is a disease caused by dengue virus (DENV), flavivirus transmitted by hematophagous females of the genus Aedes (Diptera: Culicidae) (Solomon and Mallewa, 2001). Four antigenically distinct serotypes (DENV-1–4) are known to affect humans, and it has been reported that a permanent immunity to the homologous serotype is induced during primary infection by some of these serotypes. But this immunological mechanism is a risk factor for the development of severe symptoms from reinfection of a heterologous serotype (Halstead, 2014).
DENV infections have increased since the isolation of the first strain in Africa in 1943 and are considered a global public health problem (Culshaw et al., 2017). DENV affects >100 countries, causing ∼390 million cases each year worldwide (Bhatt et al., 2013). There is a current need for specific drugs to treat dengue, and vaccines are under development; therefore, lowering the incidence of these arboviruses depends largely on the techniques used to control mosquito vectors (Dong and Dimopoulos, 2021).
Detection of infectious DENV could contribute to patient treatment, public health decision making, and the prevention of the introduction of the virus into nonendemic countries (Li-Teh et al., 2020). Vertebrate or invertebrate cell lines are frequently used for viral isolation and replication, which could increase the sensitivity of molecular or immunological tests in flavivirus typification (Avilés-Vergara et al., 2020; Guzmán and Kouri, 1996). In addition, isolation and safeguarding of strains have been useful in comparative studies of viral pathogenicity (Gutiérrez-Barbosa et al., 2020; Niu et al., 2020), immunology (Hou et al., 2020), and drug susceptibility (Hitakarun et al., 2020).
Biological models, such as the use of mosquitoes, have also been helpful in DENV isolation (Donald et al., 2020). These models require a small inoculum volume and are sensitive to low virus titers or low infectivity; however, the biological risk posed by hematophagous mosquitoes must be considered. In contrast, Toxorhynchites mosquitoes are considered a safe model for the study of arboviruses, because they are not hematophagous (Medina et al., 2012; Rosen, 1981). However, the establishment of nonhematophagous mosquito colonies has been a challenge in recent years (Medina et al., 2012; Schiller et al., 2019), which could hinder its application as a biological model.
In this study, experimental studies were conducted to determine the replication of DENV-4 in Plodia interpunctella larvae. The advantages of evaluating this lepidopteran as a study model for DENV are that it is easy to handle, and the establishment of its colonies does not require specialized equipment, and also that it is not a natural vector of DENV, nor is it capable of transmitting the virus due to its nonhematophagous condition.
Materials and Methods
This study was approved by the Biosafety and Bioethics Committee Postgraduate in Biological Sciences, Faculty of Biology, Autonomous University of Sinaloa.
Cell culture and viral infection
Aedes albopictus cell line C6/36 and supernatant from mouse brain cells infected with DENV-4 were used, which were kindly donated by Dr. Rosa María del Ángel, Centro de Investigación y de Estudios Avanzados, Instituto Politécnico Nacional, Mexico. In total, 5 × 105 C6/36 cells were seeded in a 25 cm3 culture flask and incubated for 24 h at 34°C without CO2 in minimum essential medium (MEM) culture supplemented with 10% fetal bovine serum (FBS), 1% penicillin–streptomycin, and 1% vitamins (GIBCO), until a confluence of ∼80% was obtained.
The medium was then removed and the cells were incubated with 1.5 mL of DENV-4-infected mouse brain cell supernatant, previously diluted in MEM (1:10). They were incubated for 1 h at 34°C with gentle agitation every 5 min. Then, 10 mL of MEM with 2% FBS, 1% penicillin–streptomycin, and 1% vitamins was added. Cells were incubated for 7 days in maintenance, and the supernatant was stored at −70°C.
Assays for viral infection
Two groups of 20 P. interpunctella larvae (culture established from specimens collected in 2019 in the city of Culiacán, Sinaloa, Mexico) of ∼0.20 g were immobilized (5 min at −20°C) and subsequently injected with 7 μL of supernatant of cells (C6/36) infected with DENV-4 in the dorsal–abdominal part of the segment corresponding to the penultimate pseudoleg (Fig. 1A). The negative control consisted of the inoculation of 7 μL of 1 × PBS buffer with the previously indicated parameters. Larvae were deposited in Petri dishes and kept at 30°C for 1 and 7 days postinoculation (dpi); this assay was replicated three times.

Analysis of DENV-4 replication in Plodia interpunctella larvae based on intra-abdominal inoculations. (
RT-PCR
Ten P. interpunctella larvae were randomly collected from each evaluated time (1 and 7 dpi) and macerated with a plastic micropestle in 1.5 mL tubes containing 150 μL of 1 × PBS buffer. Viral RNA was extracted from the macerates (Viral RNA + DNA Preparation kit; Jena Bioscience, Jena, Thuringia, Germany), and the purity and concentration of the RNA were determined using a NanoDrop One spectrophotometer (Thermo Scientific, Madison, WI, USA) and homogenized at 24 ng/μL in all cases.
RNA was retrotranscribed into cDNA (Script cDNA Synthesis kit; Jena Bioscience), and a PCR was subsequently performed for DENV detection with the consensus oligonucleotides D1 (5′-TCAATATATGCTGAAACGCGCGAG AAACC G-3′) and D2 (5′-TTGCACCAACCAACAGT CAATGTCTTCAGGTTC-3′) proposed by Lanciotti et al. (1992). For the PCR amplification mixture, the commercial GoTaq Green Master Mix kit (Promega, Madison, WI, USA) was used and consisted of a final volume of 12.5 μL containing 6.25 μL PCR mixture, 0.33 μL of each oligonucleotide (10 pmol), 2 μL cDNA, and 3.59 μL H2O.
The reaction was performed in a T 100 thermal cycler (Bio-Rad, Chicago, IL, USA) according to the conditions proposed by Apodaca-Medina et al. (2018), described as follows: 94°C for 4 min, 35 cycles (94°C for 1 min, 55°C for 1 min, and 72°C for 1 min), and 72°C for 5 min. The annealed PCR product was diluted 1:50 with H2O, and a second round of amplification (seminested multiplex PCR) was performed with the consensus D1 and specific TS4 oligonucleotides (5′-CTCTCTGTTGTCTTAAACAAGAGAGA-3′) that amplify a region of ∼390 bp referring to DENV-4 (Lanciotti et al., 1992).
The reaction was carried out at a final volume of 12.5 μL (6.25 μL PCR mixture, 0.33 μL of each oligonucleotide [10 pmol], 2 μL PCR, and 3.59 μL H2O) with the following amplification conditions: 95°C for 1 min, 25 cycles (95°C for 30 s, 60°C for 30 s, and 72°C for 30 s), and 71°C for 2 min (Apodaca-Medina et al., 2018).
Agarose gel electrophoresis and densitometry
A 1.5% agarose gel electrophoresis was performed for visualization of PCR products. Wells were loaded with 5 μL of reaction mixture (seminested multiplex PCR) and 0.5 μL of GelRed dye (Biotium, Fremont, CA, USA), and 1 × Tris-Borate-EDTA buffer loading buffer was used. The band size of the amplified products was determined by a 100 bp molecular weight marker (DNA Ladder; Promega). The agarose gel was run for 35 min at 80 V.
Gel visualization and image capture were performed with the DigiDoc-it system (UVP; An Analityk Jena Company, Jena, Thuringia, Germany) and the UV transilluminator, TFM-20 (UVP; Analityk Jena Company). The image was analyzed in ImageJ version 1.8.0 software (Bethesda, MD, USA) at 8-bit grayscale depth (0–255 intensity value), and the optical density (OD) of the bands (peak area) was evaluated as a parameter of viral replication.
qPCR
The products amplified by consensus PCR were used as an amplification template for DENV-4 gene detection by qPCR. The reaction volume was 20 μL consisting of 0.5 μL of each oligonucleotide (D1 and TS4), 5 μL of H2O, 4 μL of amplified product, and 10 μL of SsoFast Evagreen Supermix (Bio-Rad) with the following incubation conditions: 4 min at 94°C to denature the DNA, followed by 40 cycles of 94°C for 1 min, 60°C for 1 min, 72°C for 1 min, as well as a final extension of 72°C for 5 min.
The qPCR analysis was performed in triplicate, and DENV-4 gene expression was assessed with the 2−ΔΔCt (Livak and Schmittgen, 2001); the QuantStudio 12k Flex Real Time PCR System kit (Thermo Fisher Scientific, Waltham, MA, USA) was used. The ribosomal 18s was amplified from P. interpunctella cDNA as an endogenous control with the oligonucleotides 18s-329R (5′-TAATGATCCTTCCGCAGGTT-3′) 18s-AF (5′-CAGCMGCCGCGGGGTAATWC-3′) (Spears et al., 1992).
Statistical analysis
Survival of P. interpunctella with DENV-4 inoculum and PBS was determined by the Klapan–Meier method in 7 dpi assays and compared by the log-rank (Mantel-Cox) test. The t-Student test was used to compare the OD at 1 and 7 dpi; similarly, the 2−ΔΔCt values for both times evaluated (1 and 7 dpi) and for the positive control were compared with the Kruskall–Wallis nonparametric statistical test. The statistical analysis was performed using SPSS statistical package version 25 (IBM SPSS, New York, NY, USA). p < 0.05 was considered to indicate a statistically significant difference.
Results
The total viral inoculation assays in P. interpunctella larvae amplified a fragment of ∼390 bp corresponding to DENV-4 (Fig. 1B). These molecular fragments, when analyzed by photometry and fluorescence, with respect to PCR products, showed prospective OD and 2−ΔΔCt values at 1 and 7 dpi (OD: t-Student, t = −5 to 10.5, df = 4, p = 0.004, 2−ΔΔCt: Kruskal Wallis, H = 13.56, n = 20, df = 3, p = 0.004) (Fig. 1C, D), indicating an increase in viral load mediated by DENV-4 replication.
Larval survival decreased as inoculation time passed; however, this effect was not significantly related to viral infection since no significant differences were observed with intra-abdominal assays with PBS (Replicate 1: 95% confidence intervals [95% CI], 7 days, p < 0.733; replicate 2: 95% CI, 7 days, p < 0.248; replicate 3: 95% CI, 7 days, p < 0.826) (Fig. 2). Similarly, pupae and adults were not observed during the evaluation; therefore, no alterations in the life development of this lepidopteran in response to the virus inoculum were evidenced, at least under the conditions evaluated in the study.
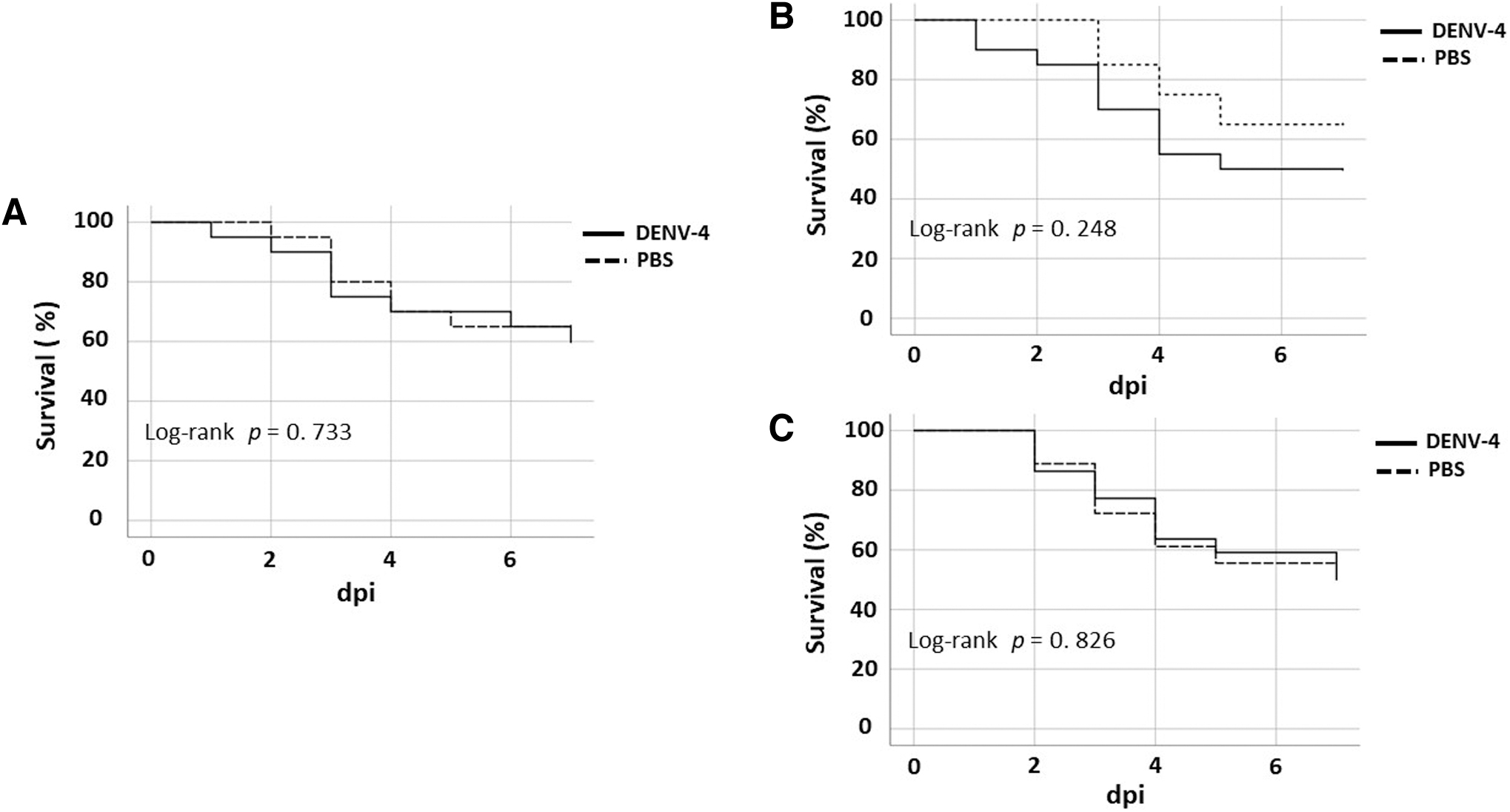
Kaplan–Meyer survival curves for the different replicates of viral inoculation in P. interpunctella larvae. (
Discussion
Evidence of susceptibility to flavivirus infection in biological systems of taxa other than culicids was documented by Suitor (1966) and Converse and Nagle (1967). The authors determined that Japanese encephalitis virus (JEV) and yellow fever virus (YFV) establish a productive and persistent infection in a cell line of Antheraea eucalypti moth tissue. These results were later corroborated, including susceptibility to infection with other viruses such as St. Louis encephalitis virus (SLV) (Yunker and Cory, 1968).
The sf9 cell line of the lepidopteran Spodoptera frugiperda has also been evaluated and found to be susceptible to infection by different flaviviruses including YFV and DENV-1 and -2 (Zhang et al., 1994). Although cell lines may have variations in susceptibility compared with multicellular biological models, it is predictable that, under anthropogenic manipulations, some arboviruses, including DENV, infect insects that do not naturally interact with these pathogens and are in phylogenetic clades distinct from natural vectors, as observed with P. interpunctella.
Wax moth larvae Galleria mellonella (Lepidoptera: Pyralidea) are infected by intra-abdominal inoculations with a wide variety of pathogens that affect humans and animals, including fungi (e.g., Cryptococcus neoformans and Candida albicans) (Ali et al., 2020; Ottaviano et al., 2021), bacteria, (e.g., Escherichia coli and Mycobacterium tuberculosis) (Jønsson et al., 2017; Li et al., 2018), and Bovine herpes simplex virus-1 (Büyükgüzel et al., 2007). However, studies of susceptibility of infection to DENV and other flaviviruses in this lepidopteran have not been reported in the literature.
The mosquito inoculation technique, for example, Aedes aegypti, A. Albopictus, and Toxorhynchites spp, is useful for the detection and amplification of the four serotypes of DENV and other flaviviruses such as Zika, YFV, SLV, and JEV. Likewise, the implementation of mosquitoes as hosts of DENV has greater potential for sensitivity with respect to the inoculation technique in suckling mice and mammalian cell lines, such as Vero (Rosen, 1981; Watts et al., 1982; Yamamoto et al., 1987). In particular, Toxorhynchites splendens could be more efficient than A. aegypti, because it tolerates a greater volume of viral inoculum and its survival is greater after intrathoracic inoculation (Rosen and Shroyer, 1985).
In reference to biosecurity, the use of mosquitoes may require specialized facilities, although these measures decrease with the use of nonhematophagous mosquitoes such as male A. aegypti and Toxorhynchites species (Rosen and Gubler, 1974). However, laboratory errors leading to the release of males could have an impact on DENV transmission mediated by sexual transmission, without ruling out the possibility of horizontal larvae–larvae transmission by Toxorhynchites species cohabiting with A. aegypti (Ortega-Morales et al., 2010). Likewise, the complexity in the establishment of Toxorhynchites colonies could be an obstacle for them to be carried out in extensive or routine studies where high densities of larvae are needed.
Regarding P. interpunctella, it completes its metamorphosis in the terrestrial environment and interaction with mosquito larvae is null. As a result, biosecurity measures for its hypothetical exploitation as a host for DENV would be less stringent. In addition, it is a species of easy and economical reproduction and does not require a specialized insectary for its reproduction under controlled conditions.
In relation to DENV-4 infection in P. interpunctella larvae, no association with survival and alterations in the life cycle of this species was determined. In response to these observations, DENV replication is probably under immune control to such a degree that the life of this lepidopteran is not compromised, as has been cited with mosquitoes (Mukherjee et al., 2019; Sim and Dimopoulos, 2010). Likewise, this mechanism probably modulates the biological development times of P. interpunctella, and avoids morphological alterations in T. splendens larvae and A. albopictus adults after inoculation with DENV-2 (Sriurairatna and Bhamarapravati, 1977; Yamamoto et al., 1987). However, studies are needed to elucidate the involvement of molecular and immunological mechanisms related to the arguments previously raised.
Studies related to the sensitivity of flavivirus infection in mosquitoes have determined important variations in the virulence of DENV serotypes, with a marked increase in DENV-1 and DENV-2; these variations do not prevent successful infection of the serotypes in mosquito models (Ekwudu et al., 2020; Rosen and Gubler, 1974) and probably do not limit their infectious potential in P. interpunctella.
Conclusions
The results demonstrate that P. interpunctella is a host, at least, of DENV-4 with the parameters established in this study. However, considerations such as elucidating the degree of susceptibility efficiency through comparisons with other biological models, for instance Toxorhynchites spp. and evaluations of replication kinetics with strains and serotypes isolated in different geographical areas, should be considered for implementation as an efficient biological model in the study of DENV.
Footnotes
Acknowledgment
The authors knowledge to Universidad Autónoma de Sinaloa for supporting this research.
Author Disclosure Statement
No competing financial interests exist.
Funding Statement
This research was funded by Programa de Fortalecimiento y Apoyo a Proyectos de Investigación (PROFAPI 2022) of the Universidad Autónoma de Sinaloa, projects: PRO_A2_001, PRO_A2_002, PRO_A2_003.