
Abstract
Objective:
The study explored larval mosquito breeding sites in urban and rural areas from March 2021 to February 2022 in the North-East region of Algeria to provide information on larval biology and ecology of different mosquito species.
Methodology:
It focused on the effect of the physicochemical characteristics of the breeding sites (pH and water temperature), area, and months on the larval mosquito abundance. Diversity index and equitability were calculated for the registered mosquito species in both areas.
Results:
Months significantly affected the larval mosquito abundances, with a clear abundance increase from March to August 2021, and then, a clear decrease to February 2022. The interactions between rural and urban areas, water pH, and water temperature as well affected the mosquito abundance. Twelve species of Culicidae were identified and distributed in three genera: Culex, Culiseta, and Aedes. Culex pipiens was the most dominant species in rural and urban sites, with a rate of 69% and 48%, respectively. In rural site, Aedes caspius and Aedes vexans occupied the second position with value of 7–8% each. In urban sites, the second position was occupied by Culiseta longiareolata (36%) and then by Culex theileri (10%). In terms of site diversity, only one site was highly diversified in rural sites, El Hchichia (H′ = 1.12 bits). In urban sites, Setif Center, Guellel an Ain Trick, had almost similar diversity with values between 1.03 and 1.04 bits. Equitability values were 0.48 in rural sites and 0.55 in urban sites. Therefore, the Culicidae species presented in these study sites are moderately balanced.
Conclusion:
The present study provides key tools for planning a better vector control through the information on the effect of some factors on the proliferation of mosquitoes in the urban and rural sites of the Setif region and on the Culicidae diversity and abundance.
Introduction
Since mosquitoes represent a serious threat to the public health, it is crucial to acquire knowledge of mosquitoes and their behavior through research on their ecology, reproduction, and feeding. Despite these important advances, mosquito-borne diseases (MBD) still represent a significant part of the global burden of morbidity and mortality (estimated at 17% of infectious diseases) and socioeconomic loss in the world (Patricia et al., 2014; WHO, 2019). The geographic expansion of mosquitoes is associated with a rising frequency of outbreaks of MBD worldwide (Wang et al., 2022). MBD have caused a high disease burden among humans, including dengue fever, chikungunya fever, Zika virus disease, yellow fever, Japanese encephalitis, and malaria, with around 96 million, 693,000, 500,000, 200,000, 68,000, and 212 million cases per year, respectively (Franklinos et al., 2019; Reno et al., 2020).
Many arboviruses have been isolated from mosquito species belonging to the genera Culex, Aedes, and Culiseta (Ayhan et al., 2022; Pfeffer and Dobler, 2010). Through their ecological plasticity, mosquitoes have been able to colonize almost all natural environments and to adapt to a very large number of habitats (Brunhes et al., 2001). They are found in almost all environments where water occurs as a necessity for their larval development (Schaffner et al., 2023). The development period from egg to adult varies among species and is strongly influenced by environmental factors. Temperature, relative humidity, and rainfall have a positive impact on the proliferation of mosquitoes (Aissaoui and Boudjelida, 2017). At the time of oviposition, the female mosquito selects the better habitat for the immature aquatic stages (Carpenter and Walter, 1955).
Since the proliferation and abundance of larval stages in aquatic habitats depend on physicochemical factors such as water temperature, and chemical composition particular to each local conditions (Benhissen et al., 2018; El Ouali et al., 2010; Paksa et al., 2019), it is essential to update the knowledge of the factors affecting the diversity and abundance changes of mosquito vectors species in specific local conditions. The analysis of the relationship between Culicidian species and the physical and chemical parameters of the breeding sites made possible to identify the parameters that are favorable to the development of mosquito species (Hapugoda et al., 2020). This will allow monitoring and assessing the prevalence and risk of their associated diseases (Medeiros-Sousa et al., 2021; Nkumama et al., 2017). Mainly, the microclimates of urban and rural housing differently affect mosquito richness and abundance (Flores Ruiz et al., 2022).
On the contrary, the agricultural activities and urbanization such as rice cultivation, watering, and fish farming have led to spread of various species of mosquitoes (Amusan et al., 2005). Due to major landscape modifications related to the settlement of novel urban structures and to cover human basic needs (e.g., Food, energy, water; McDonnell and Pickett, 1990), urbanization has been identified as a major threat to biodiversity (Alberti, 2008; McKinney, 2002; Shochat et al., 2010). Hence, the present study aims to estimate the composition, abundance, and diversity of Culicidae mosquitoes and to explore whether physicochemical parameters, such as water temperature, and pH, influence the general production in mosquito larvae in urban and rural aquatic habitats along the year in the Setif district (North-Eastern part of Algeria).
Materials and Methods
Study area and sampling sites
The study areas are represented by the district of Setif, in the North-Eastern part of Algeria belonging to the highlands (36°11′29″ N, 5°24′34″ E). It spreads over an area of 6549.64 km2 and an altitude of 1100 meters above the sea level (Fig. 1). This region is characterized by a continental Mediterranean climate with cool and rainy winters with 8.5°C average temperature and 933 mm rainfall, and long, hot, and dry summers with 29°C average temperature and 1366 mm rainfall (Added et al., 2017; Zerroug, 2012). The main part of the region tends to be semi-arid.
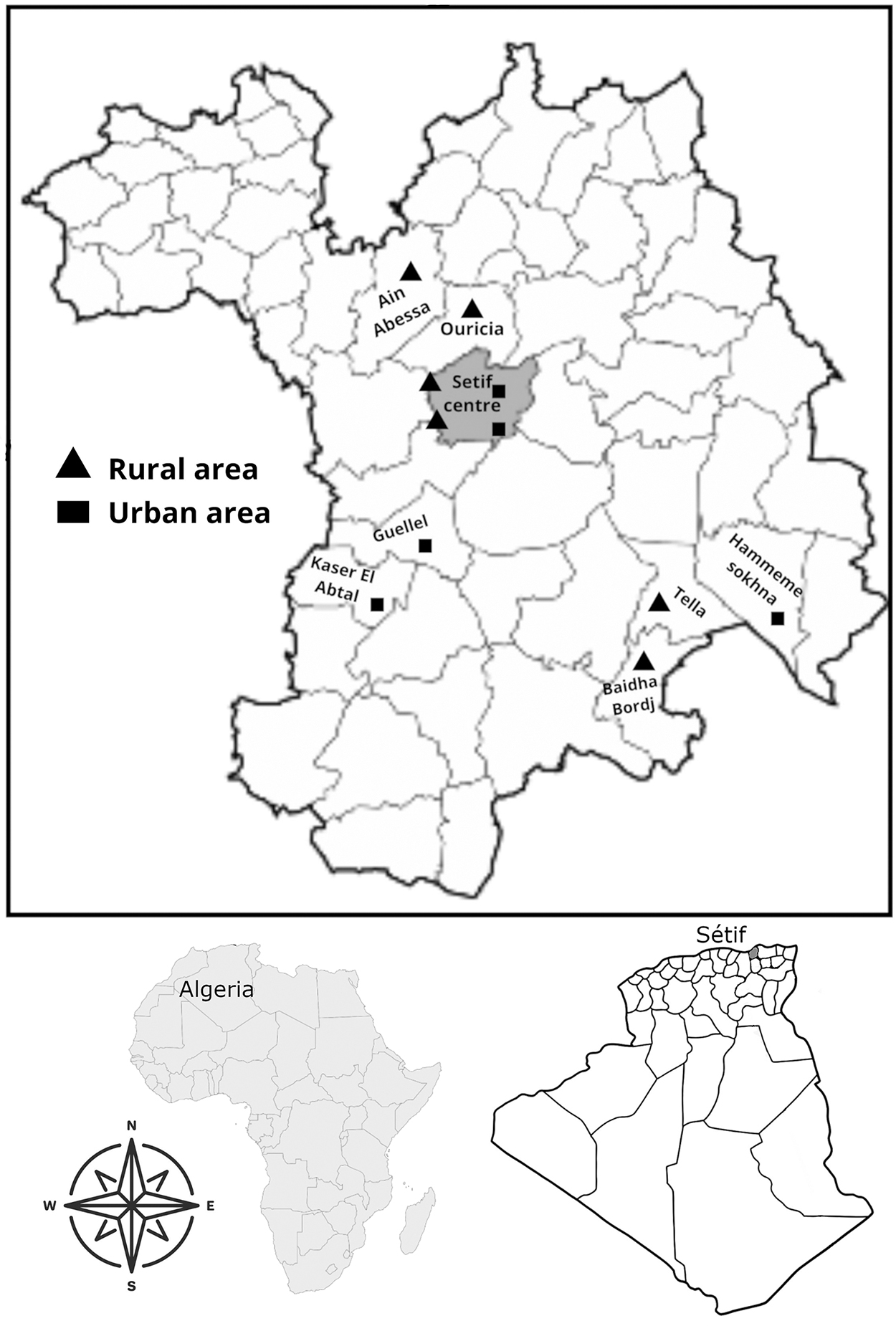
Geographical location of the 11 sites in the study area. DSA of Setif, 2011.
The study was carried out during 12 months, from March 2021 to February 2022, in 11 locations for which the ecological characteristics of mosquito's larvae aquatic habitats allowed us to divide into rural and urban sites. Six rural sites were characterized by dense vegetation, animal stables, and low human population: Ain Abessa, Baidha Bordj, Oued Bousselem, Ouricia, Tella, and El Hchichia. Five urban sites counted with high human population density, building, and factories: Ain Trick, Guellel, Hammeme Sokhna, Kaser El Abtal, and Setif Center (Fig. 1). Satellite images of rural and urban sites are available in the Supplementary Data S1 and S2, respectively.
Air temperature
Monthly minimum and maximum temperature averages were calculated from data obtained from the meteorological service of the Setif region (Ain Sfaha) using Excel 2016.
Water physicochemical parameters and specimens' collection
Preliminary prospections were realized to detect one positive breeding site per site. The breeding sites were classified and registered using the following criteria: nature (temporary or permanent), type (irrigation basin, drainage ditch, mare, valley), as well as their location and altitude were recorded using the Google Maps application for phone. The water parameters of each breeding site, water pH, and water temperature were measured during each sampling visit using digital meters (Hanna instrument, model: HI122).
In the study period, all sites have been visited two times per month, every 15 days, between 13:00 and 16:00 (hotter period of the day) to secure robustness of the results. The sampling took place using a 500 mL dipper to catch larvae during daytime and according to the standard “dipping” method cited by Silver (2008) and Becker et al. (2020). Collection details were recorded (sampling location and date) in the field for each collected specimen. The immature stages were counted and separated to the different larval stages and reared in a controlled environment in the laboratory at a temperature of 26°C ± 3°C and 12/12 h (light:dark) photoperiod. Larvae were daily fed with fresh food composed of a mixture of biscuit (Bifa special digestif blé complet) and dry yeast (75%:25% by weight, respectively) until the pupa stage was reached. Subsequently, the resulting pupae were transferred to jars containing water, with the help of a dipper, and placed in a cubic cage (30 × 30 × 30 cm) covered with a net until adult emergence. The adults were fed with 10% sugar solution (Aissaoui et al., 2022).
Morphological identification of mosquito specimens
Only mosquito larvae that had progressed to the fourth stage were mounted and morphologically identified based on their visible features with the aid of a dissecting microscope (OPTIKA B-350) and identification keys by Brunhes et al. (1999) and Brunhes et al. (2001). These were preserved in 70% ethanol until they were fixed between the slide and the cover glasses in a drop of Faure's liquid. On the contrary, for cryptic species (Aedes dorsalis and Aedes vexans), pupae were reared in emerging boxes (25 × 25 × 30 cm) until the emergence of male adults to confirm identification by the genitalia analyses. In these cases, 12 males were dissected using a stereomicroscope at 20–40 × magnification. These specimens were then mounted between a slide and coverslip and further examined under a microscope at 100–400 × magnification. Species were identified with Himmi et al. (1995) for Moroccan species and Brunhes et al. (2001) and Brunhes et al. (1999).
Data analyses
A regression model with negative binomial response was used to investigate the effect of sites, months, water pH, and water temperature on larval mosquito abundances. These analyses were performed using the R program (version 4.1.2; R Core Team 2021).
The results of the mosquito's population inventory were analyzed through different ecological indices. The total richness of a site is the number of species (S) encountered in the study area. The frequency F or relative abundance studies the distribution of a species in each region and evaluate its commonness: F = (ni/N) × 100, where ni is the number of individuals of the species and N is the total number of individuals. The Shannon–Weaver diversity index (H′) is the amount of information sample about the structure of the site from which the sample was taken and how the sample and how individuals are distributed among various species (Daget, 1976). It is calculated as follows:
Results
Effect of environmental parameters on larval abundances
From March to July, minimum and maximum air temperature increased from 19°C to 22°C, respectively. From July to August, air temperature did not change, while from August to February, minimum and maximum air temperature decreased to 16°C and 22°C, respectively (Supplementary Data S3).
All breeding sites were open and stagnant. In the rural sites, 66% were temporary, and 50% had waste. In urban sites, 60% of the breeding sites were permanent, while 66% had waste. Water temperature varied significantly along the 12 months of the study (F = 8.3253, DF = 1, p = 0.032). The period from March to August corresponds to a 6°C increasing in the water temperature (from 15°C to 21°C). Then, from September to November, it slightly varied from 16°C to 17°C. Finally, from December to February, water temperature slightly varied from 12°C to 13°C. Water pH was significantly different in both areas: 7.7 ± 0.13 in rural sites and 8.14 ± 0.16 in urban sites (F = 4.7019, DF = 1, p = 0.032).
Months significantly affected the larval mosquito abundances (Table 1), showing a bell with a clear abundance increase from March to August and a clear decrease to February (Fig. 2).
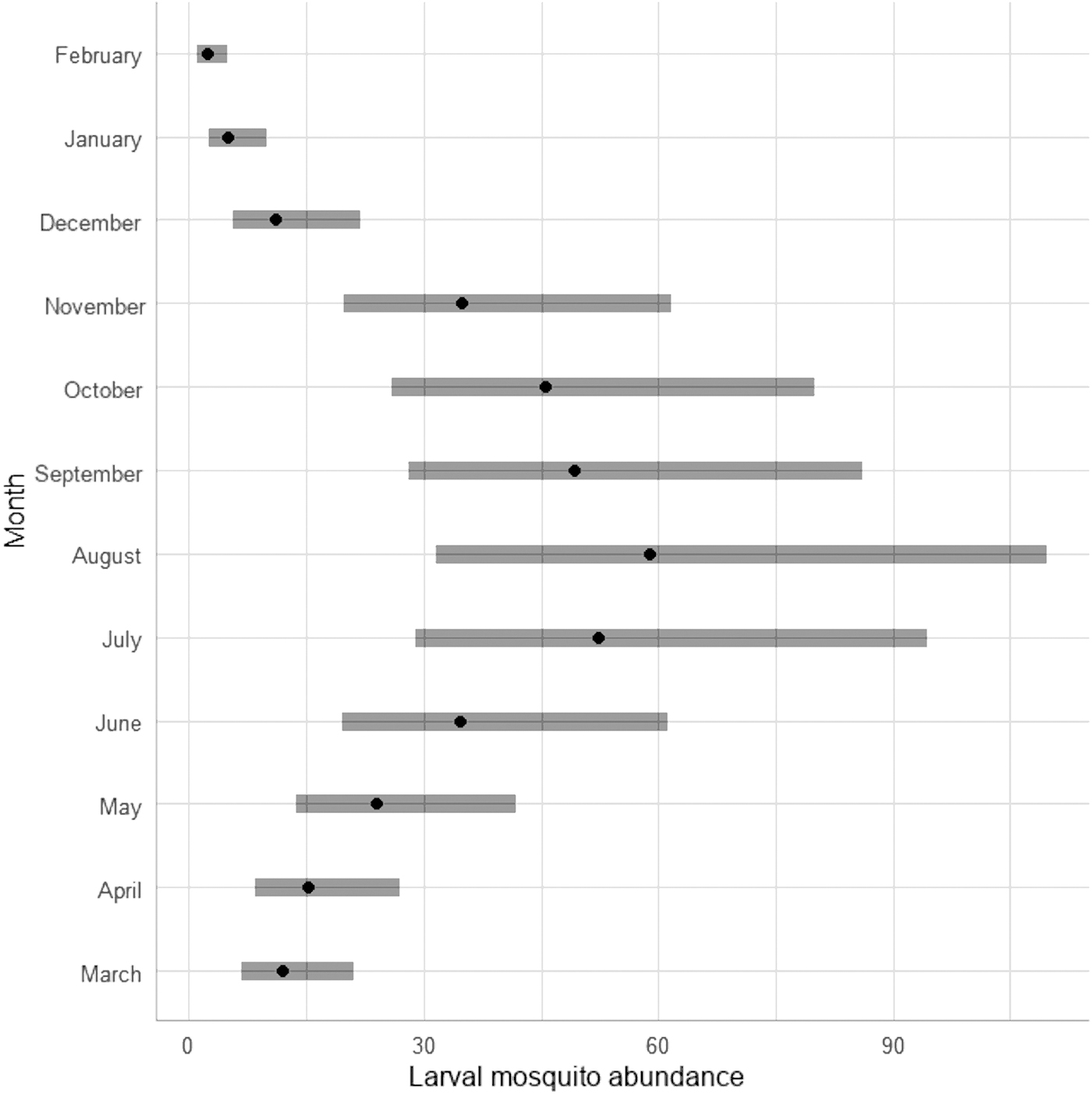
Phenology of the larval mosquito abundance from March 2021 to February 2022.
Effect of the Type of Site (Rural or Urban), Months (March 2021 to February 2022), pH, Temperature, and Their Interactions on Larval Mosquito Abundance Values
Regression model with negative binomial response.
0.05 > P > 0.01, ***0.001 > P.
NS, not significant.
The interactions between sites (rural and urban), water pH, and water temperature as well affected the mosquito abundance. In rural sites, the abundance increased with the water temperature in different slope depending on the water pH. As the pH increased from 6 to 10, the slope increased. The decrease in the abundance of larvae is inversely related to the increase in the temperature of the water and pH 6, while the abundance increased with water temperature for pH 8 and 10, with a higher slope for basic pH (Fig. 3).

Effect of interactions between sites (rural and urban) and water parameters (pH and temperature) on the larval mosquito abundance from March 2021 to February 2022.
Specimens' collections
We collected and identified a total of 4066 mosquito specimens in 11 sites of Setif. The data allowed us to record 12 mosquito species belonging to a single subfamily: Culicinae, subdivided into 3 genera: Culex, Culiseta, and Aedes (Table 2). Eleven species were collected in rural sites and eight in urban sites.
Number of the Collected Culicidae Species in Urban (U) and Rural (R) Sites of the Study Area per Month from March 2021 to February 2022
Relative abundance of mosquito species
The number of each species varied as a function of month and sites (Table 2). In rural sites, Culex pipiens was clearly the most abundant species, with a proportion of more than 69% of the total species. Aedes caspius and Ae. vexans followed far behind with abundances of 7–8%, whereas the other species were present at proportions lower than 5%. In urban sites, Cx. pipiens was also the most abundant species, with a proportion of more than 48%. Culiseta longiareolata occupied the second position with a rate of 36%, followed by Culex theileri with 10%. The other species had abundance percentages lower than 2%.
Shannon–Weaver diversity index
For rural sites, sites showed clear differences between them. The site of El Hchichia was the most diversified with the highest H′ value (1.12 bits), followed by Tella and Ouricia (0.9 and 0.72 bits, respectively), and Oued Bousselem (0.38 bits). Finally, Ain Abessa and Baidha Bordj (0.3 and 0.27 bits, respectively) were the less diversified rural sites (Fig. 4). For urban sites, diversity varied very few between four sites from 1.04 to 0.98 bits, with Setif Center and Guellel as the most diversified sites, followed by Ain trick and Kaser El Abtal, and the fifth site, Hammeme Sokhna, had no diversity at all (0 bits).
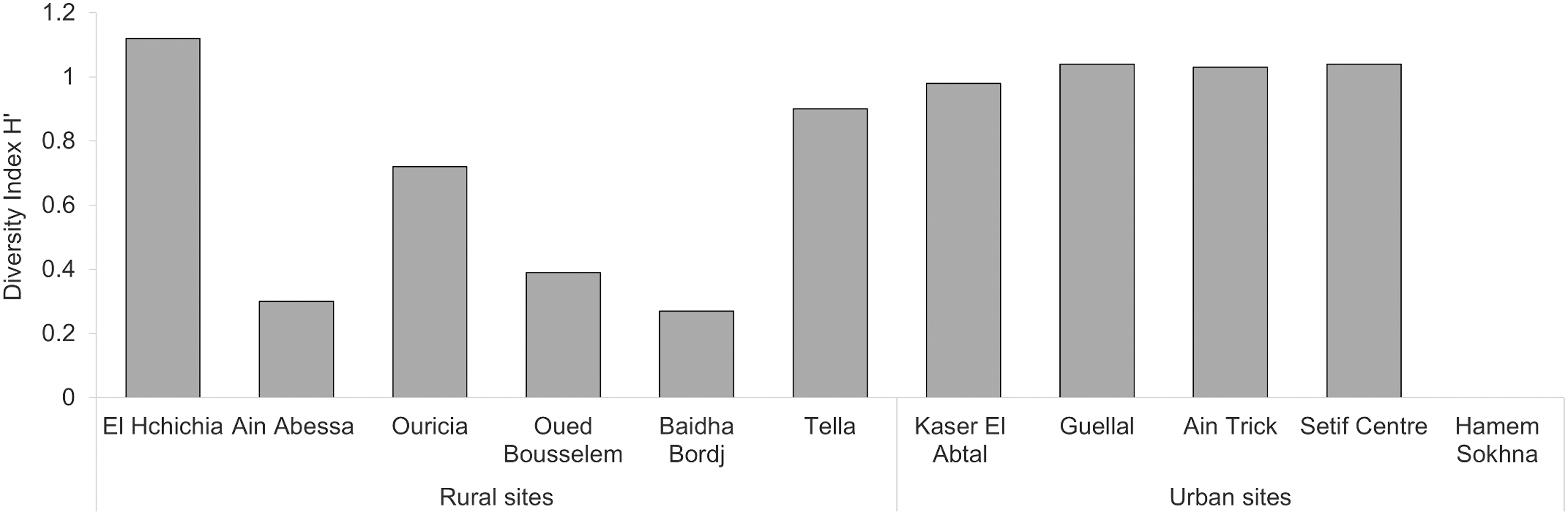
Diversity index (H′) of mosquito species in the study areas.
Equitability
The values of equitability in rural and urban sites vary between 0.48 and 0.55, respectively, meaning that the populations in these sites were not homogeneous and did not have the same relative abundance (Table 2).
Discussion
Our results indicated that two factors—months, and the interaction between water pH, water temperature, and nature of sites—significantly affected the number of Culicidae mosquito larvae in the Setif region from March 2021 to February 2022.
The abundance of larval mosquitoes showed significant variation across different months, with a notable surge from March through August, followed by a decline leading to February. The months needs to be considered a compounded factor, incorporating various elements such as air and water temperature, rainfall patterns, water pH, luminosity intensity, and photoperiod. So, maximum species density was observed in conditions where the air temperature ranged from 26°C to 36°C, water temperature from 16°C to 24.5°C, and the water exhibited a slightly alkaline pH varying from 7 to 8.5. These results are similar to the findings obtained by Sevarkodiyone and Wilson (2014), which reported comparable temperature and pH conditions (air temperature from 20°C to 27.7°C and alkaline water pH from 8 to 9) affirming our observations.
Our study demonstrated that a more basic water pH conditions result in increased abundance of larvae in both rural and urban sites, whereas acidic pH generally caused a decrease (except for urban sites at low water temperature). These observations are consistent with other authors indicating that Cx. pipiens larvae develop in water with a higher organic content and basic pH (Berchi et al., 2013), whereas other mosquito species larvae prefer neutral or slightly alkaline water (Pelizza et al., 2007; Yewole et al., 2009). On the contrary, the response of the mosquito abundances varied based on the type of area. The discrepancy in water pH between rural (7.7) and urban (8.14) sites might have influenced this variation in conjunction with water temperature. For instance, mosquito abundance increased in urban sites with acidic water pH at low temperatures. Various factors, such as differing geology between rural and urban areas (Nienie et al., 2017), rainfall, or leachate draining, could have contributed to the differences in water pH levels between these areas (Van Ha et al., 2011).
The role of water temperature is pivotal in regulating various abiotic factors and the functioning of aquatic ecosystems. It profoundly impacts the growth, reproduction, and immune responses of aquatic organisms (Hutchinson, 1957; Kataria et al., 1995). Macan (1985) noted the close relationship between atmospheric and water temperatures. In our study, it appears that atmospheric temperature influenced water temperature from March to August, as both increased in tandem. However, during autumn and winter, despite a decrease in air temperature, the water temperature remained relatively stable within a range of 2°C. This observation suggests that the soil's capacity to retain heat potentially stored energy from the summer and facilitates its transfer to the water reservoir, thereby maintaining a relatively higher water temperature (Selker and Or, 2019).
The impact of the interaction between sites (rural–urban), pH, and water temperature on mosquito abundance is validated. These findings align with the data put forth by Gaertner et al. (2017), suggesting that urban settings such as parks, roadside green areas, built surfaces, or refuse heaps might evolve into focal points for invasive species in urban environments. On the contrary, for most species, the abundance and diversity of species are generally lower in urban sites compared with rural ones. This discrepancy is largely attributed to anthropogenic activities that generate significant ecological disruptions capable of affecting animal life (McKinney, 2008).
Robert et al. (2019) reported ∼83 mosquito species in North Africa, encompassing 59 Culicinae and 24 Anophelinae. Among these, 26 species were identified as potential vectors for transmitting pathogens to both humans and animals. In Algeria, the same study cataloged 61 mosquito species, including 18 considered as potential pathogen vectors (such as Aedes albopictus, Ae. caspius, Aedes detritus, Ae. vexans, Cx. pipiens, Culex antennatus, Culex perexiguus, Cx. theileri, Culex univittatus, Culex modestus, Culex pusillus, Culiseta annulata, Anopheles labranchiae, Anopheles algeriensis, Anopheles claviger, Anopheles multicolor, Anopheles coluzzi [uncertain], and Anopheles sergentii). Our systematic investigation revealed the presence of 12 mosquito species from 3 genera: Culex, Culiseta, and Aedes. The Culex genus included eight species, with Cx. pipiens being the most abundant in both urban and rural regions, followed by Cx. theileri in lower abundance. Other species had smaller populations: Culex laticinctus, Cx. perexiguus, Culex univitattus in rural sites, and Cx. modestus, Culex torrentium, and Culex deserticola in urban sites.
In the Aedes genus, two species were recorded, primarily Ae. caspius, followed by Ae. vexans in both areas. The Aedes genus encompasses some of the most concerning vectors responsible for transmitting arboviruses such as yellow fever, dengue, chikungunya, Zika, Rift Valley Fever, and West Nile (Becker et al., 2010). In North Africa, 23 Aedes species have been reported, with 5 of them recognized as carriers of viral and parasitic diseases. These include Ae. (Stegomyia) albopictus, Ae. (Stegomyia) aegypti, Ae. (Ochlerotatus) caspius, Ae. (Ochlerotatus) detritus, and Ae. (Aedimorphus) vexans. Recent reports from Saudi Arabia, Iran, and the Middle East region of Emirate have well documented the presence of Ae. caspius (Camp et al., 2019). In addition, the Culiseta genus is represented by two species, namely Cs. longiareolata in both areas, and Cs. annulata found solely in the rural site.
Eight species identified in our study are recognized as potential vectors according to Robert et al. (2019), whereas Cx. laticinctus, Cx. torrentium, Cx. deserticola, and Cs. longiareolata were not listed in this classification. The species Cx. pipiens, Cx. theileri, Cs. longiareolata, and Ae. caspius reported in our study were also identified by Nabti and Bounechada (2019) in the same region. However, the authors additionally registered five species Culex hortensis, Culex simpsoni, and Coquillettidia richiardii that were not observed in our study. These species are considered to have no known human health importance. Two of the collected Anopheles species (in low density), An. labranchiae and Anopheles cinereus hispaniola, are known to be a malaria vector in Algeria (Boubidi et al., 2010; Snow, 2012; WHO, 2014), and in Eritrea, only An. cinereus hispaniola (Shililu et al., 2003). The absence of these species in our collection data could be explained by certain methodological differences.
While our study involved larval collection over 12 months, Nabti and Bounechada's (2019) study spanned a broader time frame from 2016 to 2019, encompassing both larvae and adult mosquitoes. Their methodology employed a standard dipper (1L) for larvae sampling and simple CDC miniature light traps (handmade: yellow light lamp and fan “12VDC”) for adult sampling in 20 sites, while our study used smaller standard dippers (500 mL) and did not involve adult collection. As a partial conclusion, our traditional taxonomic study, although not detecting all the species identified in a more comprehensive and expensive study conducted by Nabti and Bounechada (2019) (combining traditional and molecular taxonomy), successfully identified the most abundant species with fewer sampling efforts and reduced financial costs.
In this way, similar findings were observed in the Bousaada region of Algeria, where Cx. pipiens and Cs. longiareolata were documented in both rural and urban areas (Benhissen et al., 2018). The authors noted a notable correlation between the environmental site type and the abundance of mosquito larvae. Our species richness analysis indicated that rural areas exhibited higher diversity compared with urban areas, aligning with the findings of Dahchar et al. (2017) and Camp et al. (2019), who also highlighted greater species diversity in natural rural environments.
Moreover, our study reveals that the most prevalent species, Cx. pipiens, thrives in diverse breeding sites, regardless of pollution levels. This aligns with the observations of Hutchinson (1957) and Patil and Shivakumar (2021), who noted that Cx. pipiens is more abundant in heavily contaminated water sources, including those tainted by domestic and industrial sewage. While our investigation did not specifically measure chemical pollution in the breeding sites, it is plausible that rural areas such as Guellal and Kaser El Abtal and the urban site such as Oued Bousselem may have been affected by waste materials, plastic bags, and polluted water.
We observed the highest abundance of Cx. pipiens in the rural area, potentially influenced by the degree of urbanization, as suggested by Gangoso et al. (2020). Their study indicates that highly urbanized regions negatively impact the distribution and abundance of Cx. pipiens, whereas less urbanized areas are more conducive to this species. However, distribution patterns within urban ecosystems, particularly in developing regions, can vary significantly (Abella-Medrano et al., 2018; Abella-Medrano et al., 2015).
The mosquito fauna observed in the urban and rural areas of the Setif district exhibited notable variations in species diversity, as indicated by the Shannon–Weaver index (H′) and equitability (E), which displayed relatively high values in both environments. This suggests an environment conducive to species coexistence (Aissaoui and Boudjelida, 2017). Dahchar et al. (2017) and Arroussi et al. (2021) also affirmed the differences in species diversity between urban and rural sites. Specifically, the absence of diversity (as indicated by a null H′ value) in the Hammem Sokhna breeding site was characterized by the presence of a single species, Ae. caspius, typically found in stagnant water with clear characteristics, reduced dimensions, and surrounding vegetation (Brunhes et al., 1999).
A distinctive fluctuation in H′ was observed, particularly in rural areas, indicating a coexistence of species in these sites. Previous research in the Setif region established a significant correlation between the presence of Cx. theileri and the malaria vector An. labranchiae. This suggests that Cx. theileri can potentially act as an indicator species for predicting the presence of An. labranchiae in the environment (Nabti and Bounechada, 2019). This underscores the importance of monitoring Cx. theileri, serving as a potential indicator of An. labranchiae in both rural and urban areas, although further research is required to confirm this relationship. Aaroussi et al. (2021) identified several mosquito species, including Cx. theileri, Cx. modestus, An. labranchiae, and An. claviger, predominantly in peri-urban environments near streams with diverse water qualities and sunlight exposure.
In these peri-urban habitats, the predominance of Cx. perexiguus was evident (Camp et al., 2019). The level of urbanization within our sites significantly impacted the diversity and distribution of culicid mosquitoes, with Cx. pipiens being the most abundant species and serving as a prime example. Our findings align with other observations that species diversity diminishes with increased urbanization and anthropization (Ferraguti et al., 2016; Martínez-de la Puente et al., 2016).
Equitability index shows that the abundances of mosquito species were heterogeneous, indicating that some species were more abundant than others. The rural areas were slightly more heterogeneous than urban areas. A study conducted in the same region (Collo in the North-East Algeria) showed similar results, with a balance between populations in urban sites, whereas rural sites were characterized by an unbalanced population (Dahchar et al., 2017).
Conclusions
This study is an actualization of the Culicidae list in the Setif district. Now, the region counts with 12 mosquito species, 8 of them susceptible to transmit MBD to the human population. Even though Ae. albopictus was not detected in our study, it has been introduced in Algeria and detected in Alger (Benallal et al., 2019) and Annaba (Arroussi et al., 2021) the last few years, representing a risk in the emergence of autochthonous cases of chikungunya and Zika fever in important urban zones of the country (Failloux et al., 2017). The possibility of presence of the genus Anopheles is also a preoccupation for its role as malaria vector (Nabti and Bounechada, 2019). The suppression of theses vectors must be carried out with success to protect the human population.
Our study showed that even though the months are not a single factor, they help to predict the larvae mosquito abundances in the district. Then, it is recommendable to realize prevention measures campaigns, such as environmental education of the population and elimination of disposable breeding sites, from December to April, when the mosquito abundances are low. It will help to cut the life cycle of the mosquitoes and to decrease the subsequent abundances. Massive elimination measures, through insecticides nebulization of the adults, are more effective when abundances are high. Assuming that the larvae abundance is a proxy of the adult abundance, the better period to apply these control measures should be from July to October.
Footnotes
Acknowledgment
Thanks to Mr. Javier Francisco Valle Mora (El Colegio de la Frontera Sur—Unidad Tapachula, Chiapas, Mexico) who contributed to the statistical analysis.
Author Disclosure Statement
No conflicting financial interests exist.
Funding Information
This work was supported by the National Fund for Scientific Research and the Ministry of High Education and Scientific Research of Algeria (PRFU: D01N01UN190120220001 Project to Pr. Lynda Aissaoui).
Supplementary Material
Supplementary Data S1
Supplementary Data S2
Supplementary Data S3
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.