
Abstract
Background:
Fleas are ectoparasitic insects with holometabolous development. It has a hematophagous habit with mouthparts adapted to sting and suck its hosts. There are about 3000 species in the world, ∼61 in Brazil, and 19 in Rio Grande do Sul state. The objective of the research is to catalog the diversity of fleas recorded in the state, their respective hosts, and endosymbionts.
Materials and Methods:
To this end, a search was carried out in the scientific literature, from articles, books, to abstracts submitted to congresses.
Results:
The 19 species of fleas occurring in Rio Grande do Sul are divided into 7 families and 10 genera. These ectoparasites, in addition to being found in the environment, were associated with 10 different families of hosts in Rio Grande do Sul, and on the endosymbiont, agents found associated with fleas, there were 7 different species. The main agents researched in the state are Rickettsia spp. and Bartonella spp. The relationships between parasites, hosts, environment, and etiological agents present different scenarios, whether anthropized or conserved, but unknown. Sometimes, this overlap, a factor that aggravates the possibility of spillovers, either from cosmopolitan fleas in these conserved areas, or from their endosymbionts.
Conclusion:
Thus, it is important to characterize the environment so that the complexities of each location are known for the adoption of environmental and public health policies in each case. The challenges are extensive, but necessary in view of the One Health perspective.
Introduction
Fleas are wingless insects perfectly adapted to an ectoparasitic existence favored by their laterally compressed bodies, which allow to attach and walk freely through the pelage, bristles, or feathers of its hosts. They are holometabolous with biting–sucking mouthparts adapted to the hematophagous habit (mammals and birds) (Dick and Patterson, 2006; Kerkut et al., 1967). About 3000 species are known worldwide, distributed in 240 genera (Lewis, 1998). In Brazil, 61 species of 20 different genera are recorded (Avelar et al., 2013; Linardi and Guimarães, 2000; Linardi et al., 2005). States, such as Rio Grande do Sul, started to describe the flea fauna late. Until the turn of the century, Linardi and Guimarães (2000) highlighted the occurrence of only eight species in the state. Since then, some studies were carried out contextualizing the diversity of fleas in the southernmost state of the country (Antunes, 2005; Schott et al., 2020a), and, at this moment, there are 19 species of fleas, distributed in 10 genera and 7 families (Table 1). The behavior of fleas toward their hosts may vary in three different ways.
List of Flea Species Occurring in the State of Rio Grande do Sul, Their Hosts, Localities, and Endosymbiont Agents Found
This is an ethnographic study without collecting the ectoparasites for identification in the laboratory.
SFG, spotted fever group.
One of them, the insect remains in the host only for the blood meal, as an example is Pulex irritans Linneaus, 1758 (Pulicidae), and Ctenocephalides felis felis (Bouché, 1835) (Pulicidae). On the other hand, there are those that remain constantly on their host, such as representatives of the genus Polygenis Jordan, 1939 (Rhopalopsyllidae) and Craneopsylla Rothschild, 1911 (Stephanocircidae). Finally, there are the penetrating fleas (Tungidae), which enter the dermis of their host causing an inflammatory skin disease, either partially, as Hectopsylla Frauenfeld, 1860 (Hectopsyllinae); or almost entirely, as Tunga (Linnaeus, 1758) (Tunginae) (Linardi and Guimarães, 2000).
In the process of hematophagy, fleas have the potential to transmit several endosymbiont agents, some of which pose a potential risk to public health, such as in plague outbreaks, caused by the etiological agent Yersinia pestis (Lehmann and Neumann, 1896) (Barbieri et al., 2021; Carvalho, 2010; Eisen et al., 2009). Still, other etiological agents have been detected, such as those that cause bartonellosis and rickettsiosis, an opportunity in which it is still necessary to understand the role of fleas in the epidemiological cycle of these diseases as well as their vector capacity and competence (Brown and Macaluso, 2016; El Hamzaoui et al., 2020).
In Rio Grande do Sul, few studies associate the occurrence of these agents with fleas. In a preliminary analysis, after dissection of the digestive system of Ct. felis, the occurrence of Enterobacter cloacae (Jordan, 1890) and Serratia odorifera (Grimont et al., 1978) was indicated through biochemical tests in the municipalities of São Leopoldo and Novo Hamburgo (Schott et al., 2016). Analysis by PCR, genotypes of Rickettsia sp. (da Rocha Lima, 1916), Rickettsia asembonensis (Maina et al., 2016), Rickettsia felis (Bouyer et al., 2001), Rickettsia sp. Taim spotted fever group (SFG), in addition to different genotypes of Bartonella sp. (Strong et al., 1915) and Bartonella clarridgeiae (Lawson and Collins, 1996) (Table 1) (Schott et al., 2020a; Schott et al., 2019; Souza et al., 2021).
Methodology
A review study was carried out, conducted according to the recommendations of the PRISMA Extension for scoping reviews (PRISMA-ScR): checklist and explanation (Tricco et al., 2018). For this scoping review, searches were carried out in the GoogleScholar, PubMed, and Scielo databases. Scientific studies were included, such as articles, theses, dissertations, and abstracts duly published in the annals of congresses and/or conferences, in Spanish, English, and Portuguese, with a time frame for publications up to the year 2022. This search was carried out using the advanced search feature with descriptors in English (family, flea, genus), according to Medical Subject Headings (MeSH), and keyword (Rio Grande do Sul), combined with the Boolean operators OR and AND. It was considered that the genera and families of fleas occurring in Brazil were used, according to Linardi and Guimarães (2000) plus Rio Grande do Sul, in addition to the vernacular flea in English (flea), and to Portuguese and Spanish (pulga).
As an inclusion criterion, the work must verify the occurrence of fleas (Siphonaptera) in the state of Rio Grande do Sul. The research within these requirements included information on the species (or genus) of the flea, location, host, and occurrence of endosymbiont.
The identified studies had their citations grouped and loaded into the Rayyan software (Qatar Computing Research Institute, Doha, Qatar), with duplicates detected and removed by the program. Titles and abstracts were screened and then imported into the EndNote reference manager (Clarivate Analytics, PA). Potentially relevant and available works were retrieved in full and their citation details imported into a spreadsheet created by the authors. The full text of selected citations was organized into folders and assessed in detail against the inclusion criteria by two independent reviewers. Studies that did not meet the inclusion criteria were excluded. Data from the included studies were extracted by the reviewers, using a data extraction instrument developed by the authors. The extraction was presented in table form in line with the objective of this scoping review. From this, quantitative analyzes were carried out, with description and categorization of the key components, describing how the results relate to the objective.
Flea Families Occurring in Rio Grande do Sul, and Their Endosymbionts
Pulicidae
Research on fleas is largely focused on the Pulicidae family, given their abundance in urbanized regions, in addition to the possibility of being vectors of important pathogens (Azrizal-Wahid et al., 2021; Urdapilleta et al., 2021). The flea Ct. felis is the most common associated with dogs and cats and, originally from the African continent, has dispersed globally to all others, except Antarctica (Lawrence et al., 2019). In addition, they are generalist regarding their hosts and present an interface of great significance between anthropized and wild areas. Clark et al. (2018) found 138 wild animals parasitized by it from 685 samples, which corresponds to 20%.
Currently, Pulicidae occurs in 15 locations in Rio Grande do Sul (Table 1 and Fig. 1A). There are records of Ct. felis, Ctenocephalides canis (Curtis, 1826); Pu. irritans Linneaus, 1758, and Xenopsylla brasiliensis (Baker, 1904) parasitizing dogs (Corrêa 1947; Neuwald et al., 2003; Silva et al., 2018); and Ct. felis and Xenopsylla cheopis (Rothschild, 1903) in synanthropic rodents (Ribeiro et al., 2003; Winkel, 2013). There is also occurrence of Ct. felis collected from opossums, in the southern region of the state (Antunes, 2005). They were also captured in the environment, using light traps in areas with companion animals and livestock in São Leopoldo and Novo Hamburgo. In investigation by biochemical tests of the digestive tract of fleas carried out by Schott et al. (2016), found infection by En. cloacae, an emerging pathogen in neonatal intensive care units with progression to sepsis (Dalben et al., 2008),l and Se. odorifera, an opportunistic microorganism potentially infectious in humans (Lee et al., 2006).
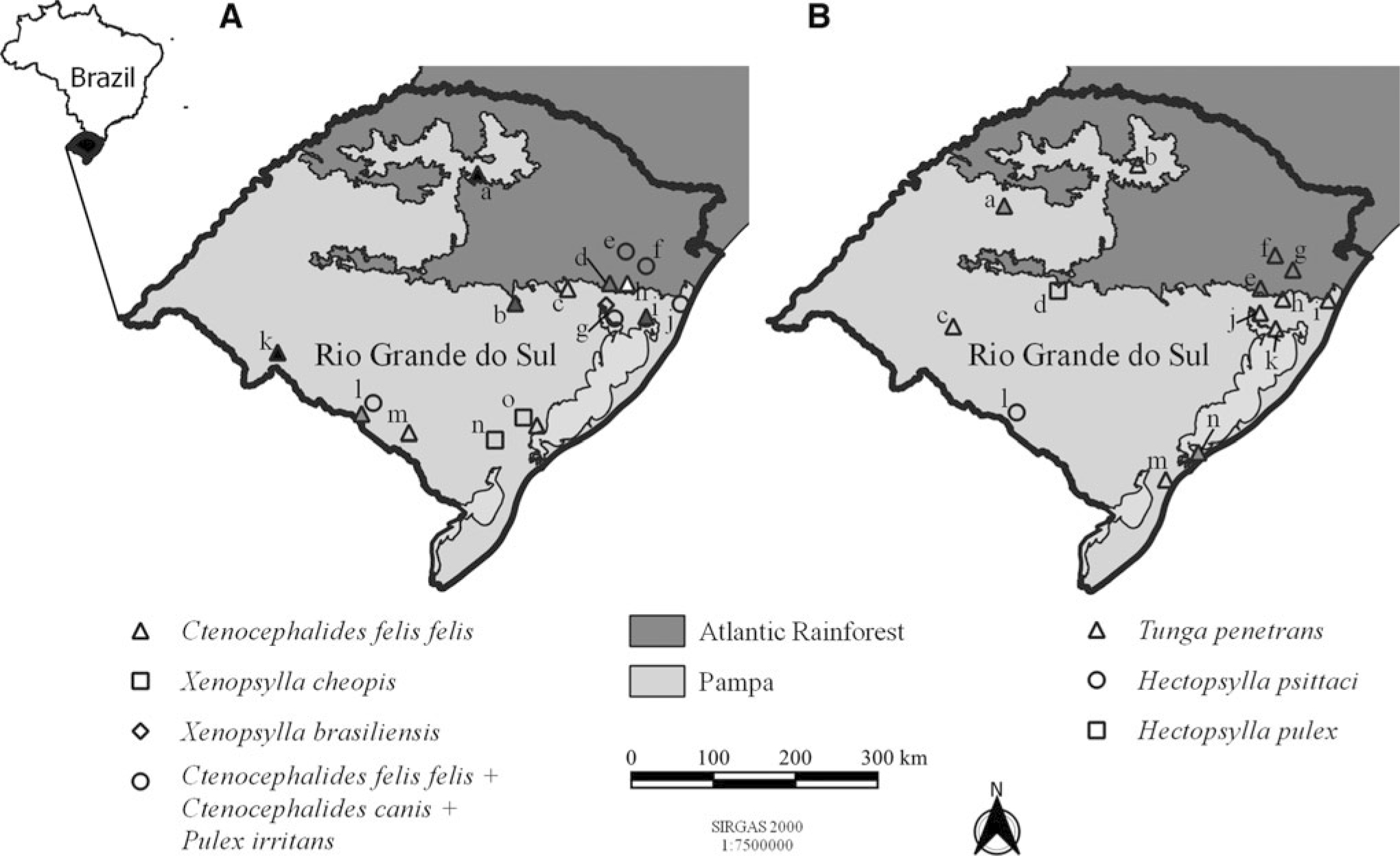
Map of the State of Rio Grande do Sul with the distribution of flea species belonging to the Pulicidae and Tungidae families. For each panel map, the light gray color corresponds to the Pampa biome and, in dark gray, to the Atlantic Rainforest biome. In insight, the map of Brazil, highlighting the state of Rio Grande do Sul in black, and referring to
In the wildlife, Schott et al. (2019) collected Ct. felis in free-ranging wild canids belonging to Cerdocyon thous (Linnaeus, 1766). In a PCR procedure, was detected Bartonella spp. genetically compatible to circulating in Ct. felis in Tunisia and Pu. irritans in the Peruvian Amazon, both collected from domestic dogs. Ri. asembonensis, agent with unknown pathogenicity, was also detected, so also Ri. felis, the etiological agent responsible for an emerging disease in the world (Brown and Macaluso, 2016). In a study with wild free-ranging felids, Souza et al. (2021) recovered Ct. felis from four felid species (Table 1), Pu. irritans on Leopardus geoffroyi (d'Orbigny and Gervais, 1844), and Xe. cheopis parasitizing Leopardus wiedii (Schinz, 1821). In molecular analysis, were detected genotypes of Rickettsia spp., Ri. asembonensis, Bartonella sp., in addition to Ba. clarridgeiae, etiologic agent with potential development of endocarditis in humans and animals (Chomel et al., 2001; Logan et al., 2019).
According to Schott et al. (2019) and Souza et al. (2021), there is a disruptive scenario for representatives of Pulicidae, such as the one described by Clark et al. (2018). Cosmopolitan fleas, abundant in anthropic environments, break this barrier and spillover in wild environments. five different species of wild animals Are indicated parasitized by Ct. felis in Rio Grande do Sul carriers of genotypes with wide distribution, thus observing a shared scenario and human influence in the relationship between environmental and animal health.
The information gap regarding the possibility of circulation of these fleas and their endosymbionts is highlighted. It is important to consider that, although it has not occurred in Rio Grande do Sul for a long time, among the representatives of Pulicidae, Ct. felis, Ct. canis, Pu. irritans, and Xe. cheopis were indicated as vectors in the locality in the outbreak of plague that occurred in the early 20th century (Di Primio, 1970). Thus, entomological surveillance with the distribution of these endosymbionts is important for One Health. In contrast, specimens of Pulicidae are registered in only 15 locations in the state, which has 497 municipalities (IBGE, 2021). It amounts to 3.02% of the cities and demonstrates the great gap of the presence of these fleas, in addition to the lack of knowledge of circulating endosymbionts with potential risk to public health.
Tungidae
Another specie that deserves attention is Tunga penetrans (Linnaeus, 1758), the responsible by disease named tungiasis, a neglected tropical disease that occurs in Latin America, the Caribbean, and Sub-Saharan Africa (Adriko, 2022; Feldmeier et al., 2014). It has great relevance in health, given the environmental, animal, and human influences on the cycle of agent, in addition to inequities related to parasitism (Carvalho et al., 2003; Feldmeier et al., 2014; Harvey et al., 2021). The negligence of the disease becomes evident when faced with the ethnographic records brought by Adomilli (2007) in reference to artisanal fishermen from São José do Norte, and Ávila (2005) regarding the parasitosis in Mbyá-Guarani indigenous people, with interethnic contact in the municipality of São Miguel das Missões (Fig. 1B). Such ethnographic records trigger the situation of vulnerability of these groups and of so many others silenced in the face of the disease. There is a lack of robust contemporary epidemiological study, with the household epidemiological survey carried out by Matias (1989), in the cities of Rio Grande and Gravataí.
The infestation by this ectoparasite, although frequent due to the anecdotes that circulate among people and in health agencies, lack scientific records on the disease and flea. The discussion does not only focus on the cycle of the etiological agent of tungiasis found in the state, Tu. penetrans, but also in the identification and location of outbreaks in cities (Fig. 1B), as may be seen in the writings of Rosário do Sul and São Leopoldo (Lopes and Peres, 2011; Serratt et al., 1996), in addition to the aforementioned survey carried out by Matias (1989). As evidenced in the ethnographic studies of São José do Norte (Adomilli, 2007) and São Miguel das Missões (Ávila, 2005), there is a tendency of severe omission of tungiasis in the territory of Rio Grande do Sul. Domestic and farm animals are considered important hosts and have the potential to disperse the disease agent (Linardi and Guimarães, 2000), and there are records of dogs affected by the disease in Gramado, Osório, and Três Coroas (Neuwald et al., 2003), and swine in the locality of Coqueiros do Sul (Pedroso-de-Paiva et al., 1997).
Cases of tungiasis are common in the various environments of the state, affecting, in addition to humans, urban and wild animals. Recently two cases of parasitism on New World primates, Alouatta guariba clamitans Cabrera, 1940, have been described from Porto Alegre and Viamão (Schott et al., 2020b). These primates exhibit arboreal behavior (Verona and Pissinatti, 2006) and were influenced by human environment, allowing the greater contact with the soil, where the ectoparasite develops their immature stages. One of the cases had a severe degree of cachexia, dehydration, and died fatally. After necropsy only lesions characterized as Tu. penetrans infestation were found and all of them were localized on its members, changing the habits and natural feeding practices of the species. This is an example of agent spillover from urbanized areas to preserved areas, creating a sanitary challenge to the local veterinarian authority (Schott et al., 2020b). In Brazil, there are other records of these spills into wild environments as described by Harvey et al. (2021). Even so, it is assumed that there is a large gap in reporting similar episodes.
Undoubtedly this ectoparasitosis deserves better attention by the health authority, especially with regard to social, educational, and environmental aspects, allowing and stimulating new studies to clarify the real epidemiological situation of the state, enabling better discussion about the conditions of the exposed, the inequities of access to basic services so that it is possible to design action measures that may intervene in the local scenario. In this context, it is to attribute an important social dimension that directly affects the principle of human dignity, protected by the Brazilian Constitution, article 1, III (Brasil, 1988) and, according to Feldmeier et al. (2014), is a problem with an intrinsic relationship with poverty, and transdisciplinary efforts are necessary to create effective protection measures to control the problem that afflicts marginalized and disadvantaged communities for a long time.
Another representative of the family, Hectopsylla pulex (Haller, 1880) have a Neotropical distribution and are semi-penetrating ectoparasites of bats. In Rio Grande do Sul, the only record of the species was on molocid, Nyctinomops laticaudatus (Geoffroy, 1805) in the city of Santa Maria (Fig. 1B) (Monteiro et al., 2005). In Brazil, in Atlantic Rainforest biome, there are records of He. pulex in two species of bats, Molossus rufus (Geoffroy, 1805) and Molossus molossus (Pallas, 1766), with a prevalence of 71.4% and 90%, respectively, in the restinga region of “Praia das Neves,” Presidente Kennedy, Espírito Santo state (Luz et al., 2009).
The Latin American Southern Cone has an occurrence of the species in northwest Argentina in Tadarida brasiliensis (Geoffroy, 1824) and Eumops perotis (Schinz, 1821), collected in the Yungas Forest and Dry Chaco ecoregions (López-Berrizbeitia et al., 2020). Considering that they are species of bats distributed throughout the region (Quintela et al., 2011), these findings are important for the characterization of the distribution of He. pulex in the surrounding regions and indicates the need for investigations into the siphonaptera fauna of bats in the region and possible endosymbionts.
Of the same genus, semipenetrating habit, and exclusive ectoparasite of birds, there is a record of Hectopsylla psittaci (Frauenfeld, 1860) for the locality of Bagé in undetermined host (Linardi and Guimarães, 2000). Considering other locations, notes on the occurrence of the species are scarce. In Brazil, there is a record in Columba livia Gmelin, 1789 (Columbidae), a bird from an anthropic environment. It has also been found parasitizing Progne chalybea (Gmelin, 1789) (Hirundinidae) and Turdus leucomelas (Vieillot, 1818) (Turdidae), both birds distributed in Latin America (Linardi and Guimarães, 2000). It also occurs in migratory hosts, as recorded in California, United States of America, in Petrochelidon pyrrhonota (Vieillot, 1817) (Hirundinidae), a bird native to the region that migrates to South America in the winter period (Nelson et al., 1979); and seabirds, such as in the Arica Desert, Chile, in Oceanodroma markhami (Salvin, 1883) (Hydrobatidae) (Cerpa et al., 2018). The diversity of habitats and host behavior matters in attention to cases of parasitism by He. psittaci, as it may have facilitated its dispersion and establishment in new locations and hosts.
Also, attention should be paid to the lack of investigation of possible pathogens that can be transmitted by these fleas associated with birds.
Ischnopsyllidae
The fleas of this family are ectoparasites exclusive to bats and, in Rio Grande do Sul, Sternopsylla distincta distincta (Rothschild, 1903) is the only representative, but without the designation of the locality and host (Table 1). In Brazil they were found in bats of the Molossidae family (Linardi and Guimarães, 2000).
This is the only clipping for the occurrence of St. distincta distincta in the state. Knowledge about this family is scarce in Brazil, as the only records come from, Ouro Preto and Curitiba, districts of Minas Gerais and Paraná states, respectively (Linardi and Guimarães, 2000). Data on endosymbionts are non-existent and the possibility of circulation of pathogens of interest to environmental and health is present. New records have been reported in neighboring countries, such as Uruguay in city of Piriápolis located in Pampa Biome, in an unidentified bat specimen (Lareschi et al., 2006); and in Argentina in bats of the genus Myotis Kaup, 1839 and Tadarida Rafinesque, 1814 (Autino and Lareschi, 1998). These data are important tools for the development of research on this family, its hosts and the possibility of detecting these endosymbionts as a way of evaluating the possible risk to environmental and animal health.
Leptopsyllidae
The record of Leptopsylla (Leptopsylla) segnis (Schönherr, 1811) in Pelotas is unique for the state (Fig. 2A and Table 1) and fleas were found in rodents of species Mus musculus (Linneaus, 1758) and Rattus norvegicus (Berkenhout, 1769) (Winkel, 2013). In Brazil, its main hosts are synanthropic rodents. However, there are records in wild rodents, such as Cerradomys subflavus (Wagner, 1842) and Oxymycterus roberti Thomas, 1901 (Linardi and Guimarães, 2000).
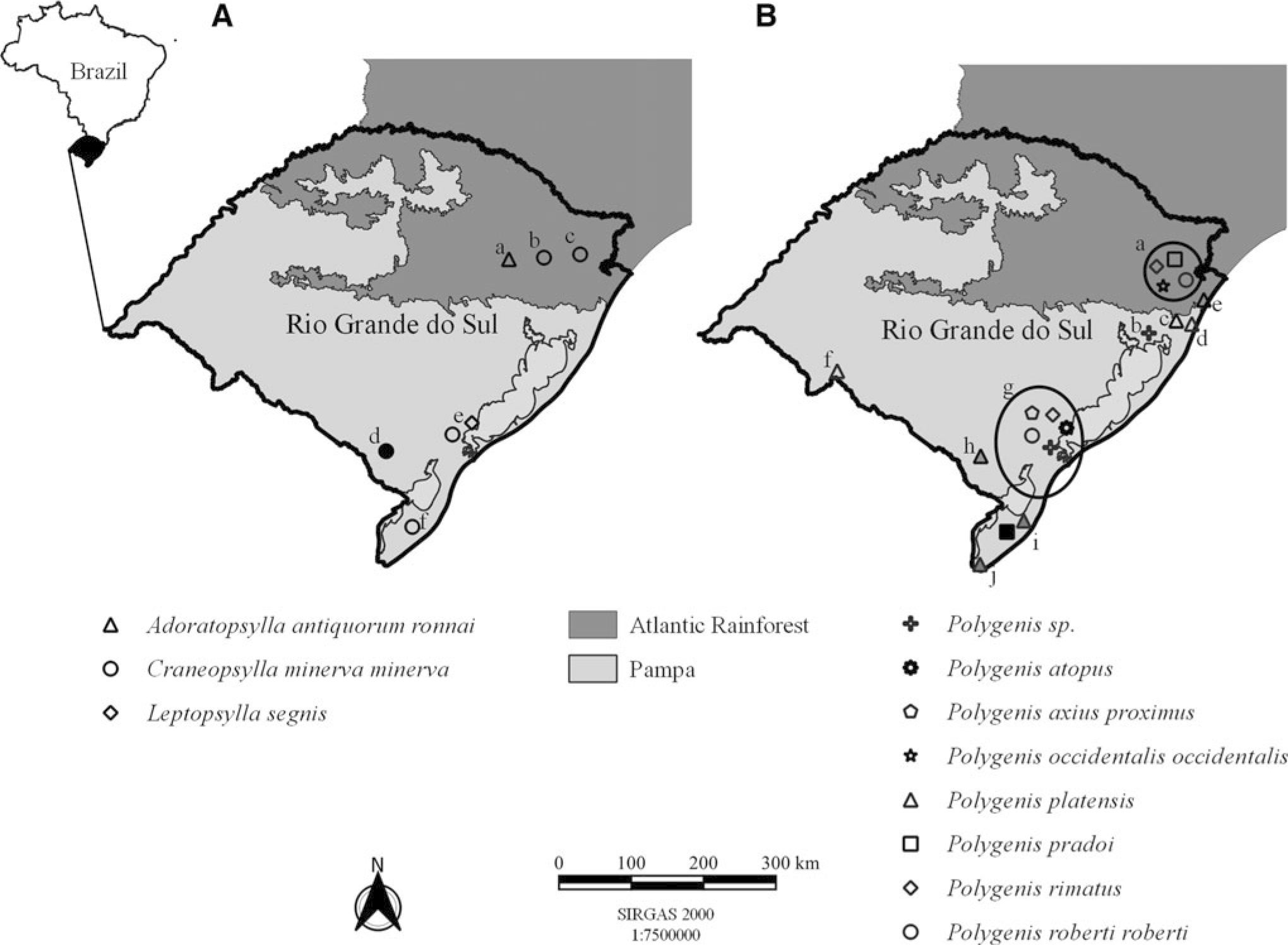
Map of the State of Rio Grande do Sul with the distribution of flea species from the Ctenophthalmidae, Leptopsyllidae, Rhopalopsyllidae, and Stephanocircidae families. For each panel, the light gray color corresponds to the Pampa biome and, in dark gray, the Atlantic Rainforest biome. In the insight, the map of Brazil with emphasis on the State of Rio Grande do Sul filled in black, referring to
In South America, there are records on Rattus sp. (Fischer de Waldheim, 1803) and Rattus rattus (Linneaus, 1758), in the province of Buenos Aires, Argentina (Castro et al., 1985). Although there is no detection of endosymbionts for Brazil, South American studies indicate the circulation of Bartonella spp. in Le. segnis collected on Ra. rattus for Sotaquí, Chile (Salas et al., 2019). Also in Chile, specimens of Le. segnis were captured in Cricetidae family, such as Abrothrix olivacea (Waterhouse, 1837) and Oligoryzomys longicaudatus (Bennett, 1832); and in Muridae family, such as Mu. musculus and Ra. rattus, being, in the latter case, positive specimen to Rickettsia sp. from the locality of Lolol, O'Higgins' region (Moreno-Salas et al., 2020).
The proximity of these fleas to urban environments, considering their tropism to synanthropic rodent hosts, presents a potential risk to One Health. Le. segnis was also indicated as a vector of plague in Rio Grande do Sul (Di Primio, 1970). Before, Jellison (1959) and Jenkins (1964) had also imputed the species as a vector of the etiological agent of plague, Ye. pestis. Thus, entomological surveillance and of agents with potential transmission by them is important to control the local status in the face of possible outbreaks.
Ctenophthalmidae
The species Adoratopsylla (Adoratopsylla) antiquorum ronnai (Guimarães, 1954) has a unique record in the state on Didelphis albiventris (Lund, 1840) (Didelphidae) for Bento Gonçalves (Fig. 2A and Table 1). This specimen was the type-species for its description. They are exclusive parasites of marsupials, in Brazil; occurring on Di. albiventris, Philander frenatus (Olfers, 1818), and Marmosa (Micoureus) demerarae (Thomas, 1905) (Linardi, 2012; Linardi and Guimarães, 2000); and Monodelphis americana (Müller, 1776) in Pedra Branca State Park, Rio de Janeiro (Oliveira 2008).
Records for the species are scarce. In Latin America, there is an occurrence in Argentina to Ad. antiquorum ronnai on Di. albiventris in Puerto Iguazú, province of Missiones (Urdapilleta, 2021). There are no records of symbionts related to the species, although Di Primio (1970) has indicated the genus Adoratopsylla (Ewing, 1925) as one of the vectors of the plague outbreak in Rio Grande do Sul. The characterization of the species ecology and possible endosymbionts are necessary steps to understand its role within the biological community and possible health risks.
Stephanocircidae
The occurrence of Craneopsylla minerva minerva (Rothschild, 1903), the only species of the family in the state, was recorded by Antunes (2005) parasitizing Di. albiventris in the region of Pelotas. Afterward, it was detected in another marsupial, Ph. frenatus, in Caxias do Sul (Schott et al., 2018). In rodents (Cricetidae), there is an occurrence in Akodon reigi (González, Langguth, and Oliveira, 1998), Brucepattersonius iheringi (Thomas, 1896), Delomys dorsalis (Hensel, 1872), Oligoryzomys sp. (Bangs, 1900), and Oligoryzomis nigripes (Olfers, 1818), in the localities of Herval, Santa Vitória do Palmar, and São Francisco de Paula (Fig. 2A and Table 1), being detected in Ol. nigripes Neotropical genotype of Bartonella sp. and Rickettsia sp. Taim SFG in the municipality of Herval (Schott et al., 2020a). In Brazil, it also occurs in both rodents and marsupials (Linardi and Guimarães, 2000).
The genotype of Bartonella sp. detected is characteristic of those circulating in fleas and rodents in Brazil. Likewise, Rickettsia sp. Taim SFG is a new agent that, in a phylogenetic analysis of the sequencing of PCR products (2050 nucleotides), integrate the spotted fever clade, according to the classification by Gillespie et al. (2008) and is positioned as a sister-sequence to Rickettsia parkeri (Lackman et al., 1965) (Schott et al., 2020a). Representatives of Rickettsia spp. SFG are traditionally associated with ticks, but new records indicate the circulation in fleas, reinforcing their vectorial potential in the dispersion of these etiological agents and in the result of diseases, as indicated by Schott et al. (2020a) and, previously, by Sekeyová et al. (2012), with the detection of Rickettsia africae (Kelly et al., 1996) in Ceratophyllus garei (Rothschild, 1902) (Ceratophyllidae) collected near Velké Blahovo, Slovakia, parasitizing the migratory bird Acrocephalus scirpaceus (Hermann, 1804) (Acrocephalidae), coming from the African continent.
South American records indicate the presence of Cr. minerva minerva in rodents of the Cricetidae family in Uruguay (Lareschi et al., 2006) and in the province of Missiones, Argentina (Urdapilleta et al., 2019). Afterward, in Missiones, there was the detection of Rickettsia sp. and Bartonella sp. of fleas collected on Ol. nigripes (Urdapilleta, 2021), parasite–host–endosymbionts relationship like to that detected in Herval, by Schott et al. (2020a). These are the first detections of endosymbiont agents associated with Cr. minerva minerva. Other surveys about the area of occurrence of the species as well as the associated microorganisms are necessary for effective surveillance of the possibility of transmission of potential diseases to communities, given the lack of information on the pathogenicity of these new etiological agents.
Rhopalopsyllidae
Among the representatives of the Rhopalopsyllidae family in Rio Grande do Sul, only representatives of the genus Polygenis are recorded (Fig. 2B and Table 1). In two studies, the species was not determined, according to findings by Antunes (2005) in Di. albiventris from Pelotas region, and Stein et al. (2010) in Ctenomys lami Freitas, 2001 (Caviidae), from Viamão. Regarding the subgenus Neopolygenis Linardi and Guimarães, 1993, there are records of Polygenis (Neopolygenis) atopus (Jordan & Rothschild, 1922) on Cavia aperea Erxleben, 1777 (Caviidae) in the State (Krüger et al. 2010), and Di. albiventris (Antunes 2005) without detection/research of associated endosymbionts. In addition, to Polygenis (Neopolygenis) pradoi (Wagner, 1937) parasitizing Akodon sp. Meyen, 1833, De. dorsalis, and Thaptomys nigrita (Lichtenstein, 1830) in São Francisco de Paula; and in Santa Vitória do Palmar parasitizing Ak. reigi and Scapteromys tumidus (Waterhouse, 1837), the latter being the PCR-positive flea for Rickettsia sp. Taim SFG (Schott et al., 2020a).
Regarding the subgenus Polygenis (Jordan, 1939), representatives of Polygenis (Polygenis) axius proximus (Guimarães, 1948) were found parasitizing Ca. aperea in Rio Grande do Sul (Krüger et al., 2010); Polygenis (Polygenis) occidentalis occidentalis (Jordan & Rothschild, 1923) on De. dorsalis in São Francisco de Paula (Schott et al., 2020a); Polygenis (Polygenis) rimatus (Jordan, 1932) on Di. albiventris in the region of Pelotas (Antunes, 2005); and Akodon sp. in the municipality of São Francisco de Paula (Schott et al., 2020a). In addition, Polygenis (Polygenis) roberti roberti (Rothschild, 1905) on Di. albiventris from the region of Pelotas (Antunes, 2005) and, in São Francisco de Paula, associated with Akodon sp., De. dorsalis, and Th. nigrita (Schott et al., 2020a). In relation to these fleas, the search for endosymbionts was not carried out or, when performed, the results were negative.
About Polygenis (Polygenis) platensis (Jordan & Rothschild, 1908), they were collected on Ctenomys flamarioni (Travi, 1981), and Ctenomys minutus (Nehring, 1887) in the localities of Capão da Canoa, Osório and Tramandaí (Linardi et al., 2005). They were also found on Akodon sp. in the localities of Herval and Santa Vitória do Palmar, Oligoryzomys sp. in Herval and Santana do Livramento, in addition to Oxymycterus nasutus (Waterhouse, 1837) in Chuí, all of which were negative for endosymbionts. Furthermore, there was a collection in Ol. nigripes from Herval positive for Bartonella sp., and in Santa Vitória do Palmar positive to Rickettsia sp. Taim SFG. In addition, Akodon azarae (Fischer, 1829) and Sc. tumidus from Santa Vitória do Palmar, and Ox. nasutus from Chuí, were positive for both microorganisms (Schott et al., 2020a).
The genus has an important distribution throughout the State and is found in both biomes, Pampa and Atlantic Rainforest. In Brazil, Po. atopus, Po. roberti roberti, and Po. rimatus, together with Rhopalopsyllus lutzi lutzi (Baker, 1904) and Rhopalopsyllus lugubris (Jordan & Rothschild, 1908) were caught in marsupials from areas of São Paulo state (Horta et al., 2007). In the state of Rio de Janeiro, Po. atopus, Po. rimatus, and Po. roberti, in addition to Polygenis (Neopolygenis) pygaerus (Wagner, 1937), and Polygenis (Polygenis) tripus (Jordan, 1933) were collected in rodents in the Serra dos Órgãos region (Carvalho et al., 2001). In Argentina, Misiones region, representatives of Po. platensis, Po. rimatus, Po. roberti, and Po. pradoi were collected associated with rodents (Cricetidae) and marsupials (Urdapilleta et al., 2019). There was also collection of Po. platensis on armadillos Chaetophractus vellerosus (Gray, 1865) (Xenarthra) in the province of Buenos Aires, Argentina (Ezquiaga et al., 2017). In Uruguay, the occurrence of Po. atopus shared common occurrence hosts in Rio Grande do Sul (Lareschi et al., 2006).
Concerning the endosymbionts associated with representatives of Rhopalopsyllidae in other locations in Brazil, five specimens of Po. atopus (5/99) were positive for Rickettsia spp. in the state of São Paulo (Horta et al., 2007). Specimens of Polygenis (Polygenis) bohlsi bohlsi (Wagner, 1901) fleas were also collected on Thrichomys fosteri (Thomas, 1903) (Rodentia, Echimyidae) positive for Bartonella spp., and the same microorganism in its host in Corumbá, Pantanal region, in the state of Mato Grosso do Sul (Sousa et al., 2018). In Argentina, La Plata region, Ri. felis was detected in Polygenis (Polygenis) axius axius (Jordan & Rothschild, 1923) (Melis et al., 2020).
Another important factor concerning to plague, although not determined in Rio Grande do Sul, the genus Polygenis had a high number of naturally infected specimens from 1966 to 1982 in northeastern Brazil. Of 238 infected fleas, 211 specimens belong to the genus Polygenis, corresponding to 88.65% (Almeida et al., 1985). Also, it is important to study the diversity of the family and the possibility of transmission of etiological agents of various diseases, because, in addition to the occurrence of bacterial endosymbionts, there are other groups of organisms associated with fleas to complete their biological cycle, according to findings of Allantonematidae, Hymenolepididae, and Trypanosomatidae on Po. tripus collected from Ne. lasiurus, and Ce. subflavus, in Belo Horizonte, Minas Gerais State (Botelho and Linardi, 1992).
Endosymbionts and Their One Health Approaches
The One Health approach advocated by the World Health Organization is complemented with the epidemiological cycle of disease; and, in this case, associated with a vector (Johnson et al., 2018). The environment, in a natural way, has the set of biotic and abiotic elements balanced and the break of this paradigm causes damage to the health of the environment, as well as to animals and humans inserted in that context (McMichael and Beaglehole, 2000). Recently dispersed etiologic agents are not necessarily new. These, in a preserved biological community, are self-sustaining in the ecological relationships established in that environment. A disruption of any One Health triad element is likely to spill over from any given pathogen (Lafferty and Kuris, 2005).
The research on flea endosymbionts in Rio Grande do Sul is about the circulation of Bartonella spp. and Rickettsia spp., according to Souza et al. (2021), Schott et al. (2020a), and Schott et al. (2019). Different scenarios of the dispersion and origin of these bacterial endosymbionts were verified. The detection of Bartonella spp., Ba. clarridgeiae, Ri. assembonensis, and Ri. felis in Ct. felis (Schott et al., 2019, Souza et al., 2021) configures a scenario of anthropic influence even if dispersed in environments far from cities. According to Clark et al. (2018), these fleas are interacting with the wild environment in a more intense way. There is an intervention in the balance of these communities, bringing non-natural elements that start to settle in these environments, given the cosmopolitan condition of Ct. felis (Clark et al., 2018). Observing with the One Health approach, there is a different behavior to environmental health with possible reflexes to animal health and human health. They are pathogenic or potentially pathogenic agents and their establishment in these new places may generate future changes within the biological complexity (Lafferty and Kuris, 2005).
In contrast, the detection of Bartonella sp. in Neotropical fleas from wild rodent hosts presented a different scenario (Schott et al., 2020a). Molecular analysis of endosymbiont agents formed a particular clade between detections of the agent in these fleas and their rodents in Rio Grande do Sul (Schott et al., 2020a), along with other detections in Brazil, such as those performed by Sousa et al. (2018) in Po. bohlsi bohlsi, and Gonçalves et al. (2016), directly in wild and synantropic rodents. In these cases, although heterogeneous environments, there is a particular configuration within the previously restricted Neotropical biogeographic region.
In the same sense, we have the detection of Rickettsia sp. Taim SFG in Cr. minerva, Po. pradoi, and Po. platensis, Neotropical fleas on native wild rodents (Schott et al. 2020a). This scenario is quite specific as it is restricted to the localities of Santa Vitória do Palmar and Chuí, in the extreme south of Rio Grande do Sul and associated with the area of the Taim Ecological Station. This agent is placed in the SFG, according to the classification by Gillespie et al. (2008), thus showing zoonotic potential (Schott et al., 2020a), although its pathogenesis is unknown so far.
The configurations of the One Health approach are important for the region, given the very specific delimitation of the area of occurrence and being linked to a federal conservation unit. Knowledge of the species, its determination, and risks are imperative for the establishment of future sanitary measures in case of proof of pathogenicity and the maintenance of the prevention status of the conservation unit in which the agent is found in biological balance. On the other hand, the scarcity of research with the scope of identifying endosymbionts of ectoparasites in the region, mainly in fleas, is highlighted, making possible the coverage of the area to be greater than that determined at present.
Observing the reality of Rio Grande do Sul, although there is the configuration of different scenarios, there is no way to fail to associate each geographic space, its environmental configurations, and conditions in common. The circulation of the agents in these fleas is actually occurring. There is an anthropized scenario related to some diseases and others to be characterized and, sometimes, these spaces are overlapped. Surveillance within the One Health approach is essential and acts in partnership with the conservation of environments, the healthiness of cities as a way of maintaining biological balance and ecosystem services. It is known that disturbances are essential to break these paradigms and affect One Health, usually showing the first signs in the sphere of environmental health (Johnson et al., 2018; Lafferty and Kuris, 2005; McMichael and Beaglehole, 2000). Parsimony between the environment and human occupations are essential for health and reducing potential (re)emergence of disease.
Final Considerations and Perspectives
Gap is the term that exemplifies the study of fleas and endosymbionts in Rio Grande do Sul. This is not a particularity of the region. Although studies on the diversity of fleas and their endosymbionts have grown in recent years, there are vast regions that are silenced. For that, surveys are needed to have a more realistic picture of the state's condition. As next steps, given the circulation of important etiological agents, in addition to the description of these species, verification of pathogenicity, the real vectorial capacity of fleas in the dissemination of these diseases should be analyzed. The challenges are large and extensive, but essential within the One Health approach.
Footnotes
Authors' Contributions
D.S.: conceptualization, methodology, investigation, writing original draft, review and edition, and project administration. F.L.R.: methodology and investigation. F.N.S.: formal analysis, writing review and edition, and supervision. R.W.d.C.: formal analysis, resources, writing review and edition, supervision, and funding acquisition.
Author Disclosure Statement
No conflicting financial interests exist.
Funding Information
This study was financed in part by the Coordenação de Aperfeiçoamento de Pessoal de Nível Superior—Brasil (CAPES). Finance Code 001.