
Abstract
Background:
This study conducted in the departments of Oueme and Plateau aims to assess the presence of the dengue virus and its different serotypes in Aedes aegypti and Aedes albopictus, as well as the epidemic risk incurred by the populations.
Methods:
Collections of adult mosquitoes using human landing catches (HLC) were carried out in six communes, three (Porto-Novo, Adjarra, and Avrankou) in the Oueme department and the rest (Ifangni, Kétou, and Pobè) in the Plateau department. Pools of ten Aedes mosquitoes were formed, and stored at −80°C in RNA later. RT-PCR was used to detect dengue virus, and conventional PCR for the different serotypes. Inspection of water containers and collection of Aedes larvae was performed inside and around each house to calculate the stegomyan indices.
Results:
In the six communes, the dengue virus was present both in Ae. aegypti and Ae. albopictus. Combined data of the two Aedes species at the communes level revealed infection rates ranging from 80.00% (95% CI: 61.43–92.29) to 96.67% (95% CI: 82.78–99.92). In all the communes, the values of stegomyan indices reached the WHO threshold, which indicates the existence of the risk of an arbovirus epidemic. In addition, the infection rates were similar for Ae. aegypti [88.19% (95% CI: 81.27–93.24)] and Ae. albopictus [86.79% (95% CI: 74.66–94.52)]. The three virus serotypes detected in the pools of Aedes were DENV-1, DENV-3, and DENV-4, with a high prevalence for the first two.
Conclusion:
This study revealed that three serotypes (DENV-1, DENV-3, and DENV-4) of dengue virus circulate in Ae. aegypti and Ae. albopictus in the departments of Oueme and Plateau. Moreover, the risk of transmission of arboviruses was globally high and variable from commune to commune. This information is essential for informed decision-making in the preventive control of the disease.
Introduction
Dengue fever is a hemorrhagic and potentially fatal disease transmitted by Aedes mosquitoes (WHO, 2017). Its global incidence has increased over the past four decades (Messina et al., 2014). Increasingly, many countries have started reporting their first outbreaks. The number of dengue cases reported to WHO has increased more than eightfold over the past two decades, from 505,430 cases in 2000 to over 2.4 million cases in 2010 and 4.2 million cases in 2019 (WHO, 2020). The number of reported deaths shifted from 960 in 2000 to 4032 in 2015 (WHO, 2020). In 2019, an epidemiological rebound in dengue regardless of the continent was observed (Aubry and Gaüzère, 2023). This disease, whose occurrence was limited to Latin America and Asia in the past, has now spread to West Africa with outbreaks in Burkina-Faso, Nigeria, Côte d’Ivoire, Niger, and Senegal (Stoler et al., 2014).
The disease is caused by an infection with dengue virus (DENV), which exists as four genetically distinct serotypes (DENV-1 to DENV-4) (Calisher et al., 1989). Although an infection with one of these serotypes confers long-term immunity, the latter is only acquired for this type, and not for the other three (Gubler, 2002). Several studies have shown that an infection with different serotypes can cause a severe and potentially life-threatening form of dengue fever, due to a reinforcement of antibodies (Bhatt et al., 2013; Bravo et al. 1987). The emergence of severe dengue fever in tropical environments can be partly attributed to the co-circulation of different serotypes in the same geographical area (Kraemer et al., 2015). In sub-Saharan Africa, existing information on dengue fever is unreliable due to unreported cases, low level of awareness among health care workers, and confusion of this ailment with other common febrile illnesses such as malaria (Amarasinghe et al., 2011).
Several West African countries have experienced epidemics with the presence of different serotypes. Indeed, serology-based dengue surveillance studies conducted in Nigeria showed a high level of infection in the population (Kolawole et al., 2017; Mustapha et al., 2017). Over the past three decades, several dengue epidemics occurred in Côte d'Ivoire (Ano et al., 2020). Also, three serotypes of the virus (DENV2, DENV3, and DENV4) were found to circulate in Burkina-Faso (Amarasinghe et al., 2011; Ridde et al., 2016; Saluzzo et al., 1986). Several confirmed cases of dengue fever were detected in Senegal and attributed to serotypes DENV1 and DENV3 in 2017 and 2018 (Diagne et al., 2019). Dengue was also detected in travelers departing from Benin to France (Gautret et al., 2010; Fourié et al., 2020). In this country, Tchibozo et al., (2022) recently detected the circulation of DENV3 in Aedes aegypti.
The high malaria prevalence in West Africa makes it difficult to detect arboviruses and other nonmalaria febrile illnesses, because they have similar symptoms. About 70% of febrile illnesses diagnosed in sub-Saharan Africa are mistakenly cared as malaria (Aubry and Gaüzère, 2023). The main vectors that transmit these different serotypes of dengue virus are Ae. aegypti and Ae. albopictus, two mosquito species commonly encountered in the urban areas of Benin and that live in sympatry (Yadouleton et al., 2022). Aedes aegypti was found throughout the country, while Ae. albopictus was only collected in the Southern part (Padonou et al., 2023; Yadouleton et al., 2022).
A study by Padonou et al. (2020) showed a strong infestation of the municipality of Abomey-Calavi by Ae. aegypti. In control programs, isolation of dengue virus from patients is key for monitoring the disease. Unfortunately, this approach detects the virus when it already strongly circulates in the community. Detecting viral activity in local mosquito populations is an essential step in dengue prevention and control (Bhatt et al., 2013). So this study was conducted to evaluate the epidemic risk and detect the different serotypes of the dengue virus (DENV) in populations of Ae. aegypti and Ae. albopictus in the departments of Oueme and Plateau.
Method
Study area
Mosquito samples were collected from six communes in the departments of Oueme and Plateau, Southern Benin. These two departments border Nigeria, a dengue-endemic country (Mustapha et al., 2017). They are characterized by a subequatorial climate with two rainy seasons (March-July and September-November), with the rest of the year being the dry seasons. The annual rainfall ranged between 1,300 to 1,500 mm. Except for the commune of Porto-Novo, all the remaining study communes (Adjara, Avrankou, Ifangni, Pobè, and Kétou) are rural, with agricultural activities that include production of cassava, bean, maize, millet, and palm oil, livestock farming, and hunting. Palm oil production requires water to be stored in containers. The lack of drinking water in these communes led the inhabitants to store water in containers for their household requirements.
Data collection was performed in the communes of Porto-Novo, Adjarra, Avrankou, Ifangni, Pobè, and Kétou, as shown on the map in Figure 1.

Map of the study area.
Assessment of the epidemic risk of dengue
About 5% of the houses of 2 villages selected in each study commune were investigated. After greeting the occupants of the houses, to which a brief description of the objectives of the survey was provided, inspection of water containers and collection of Aedes larvae were carried out inside and around each house. Each interviewer used a tablet to record GIS data and general information. A breeding site was considered positive when it housed at least one Aedes larva. The collected data allowed the record of different information about encountered breeding sites. The gîtes encountered fall into several categories: Domestics containers: Any hollow object capable of holding water, in particular for storing or transporting (buckets, jars, canaries, barrels, cans, bottles, flower pots, cisterns, drinking troughs, etc.) Tires Discarded containers: Plastic bags and tins. Natural containers: Tree hollows, sheathing plants (leaves and flowers), and coconut. Other: Abandoned cars, salt mortars, wheelbarrows, and fruit shells.
Epidemiological monitoring indicators were calculated. When the House Index (HI) is greater than 5% and/or the Breteau Index (BI) greater than 20 in a given area, the transmission of arbovirus diseases is likely to occur. These indices (BI and IH) are generally used to define at-risk (priority) areas, to better plan control intervention (WHO, 2003). The stegomyan indices were calculated as follows: Breteau index (BI): Number of positive containers for 100 surveyed houses; House index (HI): Number of positive houses × 100/number of visited houses. Container Index (CI): Number of containers containing water with the presence of larvae or pupae × 100/Number of visited containers.
Mosquito collection
From July 2021 to October 2022, monthly rounds of human landing catches were organized to sample adult mosquitoes. In each of the six (6) study communes, a central site and a peripheral site were selected for human landing catches between 7 a.m. and 6 p.m. During the collections, a first team of 8 people/communes worked from 7 a.m. to 1 p.m., and was replaced by another team of 8 people. In each house, a collector was installed inside and a second outside, which makes a total of 4 collectors/house/day. To reduce bias related to the ability of collectors in capturing mosquitoes and their attractiveness, they were rotated between the different houses. At each capture point, the seated collectors who served as baits collected the mosquitoes that landed on their bare legs or feet using hemolysis tubes before they bite.
The field-collected larvae/pupae were transported to the CREC insectary for rearing until adult stage. After emergence, mosquitoes were morphologically identified per breeding site and per commune using taxonomic keys of Edwards (1941) and Yiau-Min (2004). The main vectors of arboviruses (Ae. aegypti and Ae. albopictus) were kept aside and stored on RNA later at −80°C before the extraction of dengue virus RNA.
Extraction of the viral RNA and synthesis of the complementary DNA
RNA extraction was performed on pools of 10 mosquitoes of Ae. aegypti or Ae. albopictus, using Qiagen RNeasy Mini Kits® (Qiagen, UK). Thus, the grinding of the 10 mosquitoes of each pool was carried out in 150 µL of the RTL lysis buffer, using the Qiagen tissue lyser. Around 150 µL of 70% ethanol was added to the ground material and the supernatant obtained was transferred to RNeasy plates. The RNeasy plates were then sealed with the AirPore tape sheet and centrifuged at ∼5600 g for 2 min. This was followed by a series of three centrifugations at ∼5600 g for 2 min, 2 min, and 4 min after respective addition of 100 µL of buffer RW1, 100 µL of buffer RPE, and an additional 100 µL of buffer RPE.
After this step, 45 µL of RNase-free water was added to each well and incubated for 1 min at room temperature, and then centrifuged at ∼5600 g for 2 min. The concentration of the extracted RNA was measured using the Nanodrop and stored at -80°C. Before cDNA synthesis, 2 mg of extracted RNA will be DNase treated using the Promega RQ1 RNase-Free DNase Kit (#M6101), as described below:
The DNase digestion solution was prepared with a reaction mix containing 2 mg of extracted RNA, 1 µL of Buffer RQ1, 2 µL of RQ1 DNase, and 10 µL of nuclease-free water. The mixture underwent a first incubation at 37°C for 4 min. After this step, 1 μl of the DNase stop solution was added and the mixture was incubated at 65°C for 10 min to inactivate the DNase. After this treatment, the cDNA was synthesized using large capacity cDNA reverse transcription kits (Applied Biosystems #4368814), according to the manufacturer's procedure. The cDNA concentration was measured using the Nanodrop, and stored at -80°C before the PCR, which allowed to detect the dengue virus and its type.
Detection of the dengue virus and its serotypes by PCR
These were performed following the protocol described by Pérez-Castro et al. (2016). This protocol provides for two series of amplifications: A first that detects the presence of the virus, and a second, the serotype. The first PCR was performed using a thermocycler (Eppendorf Nexus) with a total reaction volume of 25 µL containing 10 pmol of each mD1 and D2 primers using a HotStar Taq Master Mix kit (Qiagen). PCR was performed by a denaturation step at 95°C for 15 min, followed by 35 cycles of 94°C for 15 s, 55°C for 15 s, 72°C for 30 s, and an extension of 72°C for 10 min. A fragment of 511 base pairs is expected for this first amplification.
The second PCR, which allowed the detection of dengue serotypes, was carried out with the same Hotstartaque mastermix kit, with 5 μl of PCR product from the previous reaction and 10 pmol of each primer (mD1, rTS1, mTS2, TS3, and rTS4) in a reaction volume of 25 µL. The amplification was composed of one cycle at 15 min, followed by 25 cycles at 95°C for 15 s, 55°C for 15 s, and 72°C for 30 s, and a final extension of 7 min at 72°C. The PCR product was analyzed by electrophoresis on a 2% agarose gel stained with ethidium bromide. The serotypes were determined according to the size of the amplicons. Indeed, fragments of 208 bp, 119 bp, 288 bp, and 260 bp are expected, respectively, for DENV-1-, DENV-2-, DENV-3-, and DENV-4-positive mosquitoes.
The sequences used for the detection and typing of dengue serotypes are as follows:
mD1: 5′-TCAATATGCTGAAACGCGAGAAACCG-3′;
D2: 5′-TTGCACCAACAGTCAATGTCTTCAGGTTC-3′;
rTS1: 5′-CCCGTAACACTTTGATCGCT-3′;
mTS2: 5′-CGCACAAGGGCATGAACAGTTT-3′;
TS3: 5′-TAACATCATCATGAGACAGAGC-3′;
rTS4: 5′-TTCTCCCGTTCAGGATGTC-3′.
Statistical analysis
The Chi 2 test of comparison of multiple proportions was used to compare the proportions of the different breeding sites within the same village.
The infection rate was calculated using the following formula: nP*100/N, where nP is the number of positive mosquitoes and N is the number of mosquitoes tested.
The binomial test was used to determine confidence intervals for both the stegomyan indices and the prevalence of infection to the dengue virus in Ae. aegypti and Ae. albopictus. All statistical analyses were performed with the R software, version 4.13.
Ethical considerations
The protocol of this study was reviewed and approved by the Institutional Ethics Committee for Health Research of the Center for Research in Entomology of Cotonou (N°06–22/CREC/CIERS-CREC/SG). Before the involvement of mosquito collectors in the study, they gave their consent. The risk that they suffer from Aedes or other vector borne diseases was minimized as they were trained to catch the mosquitoes before receiving any bite. They were also vaccinated against yellow fever and regularly monitored. In case of confirmed fever, they were immediately cared in the closest health facility.
Results
Type of Aedes breeding sites
The different Aedes breeding sites encountered, as well as their relative proportions indoors and outdoors in the department of Plateau and Oueme, were mentioned in Figures 2 and 3, respectively. In the domestic environment (Indoor) of the Plateau department, four types of breeding sites were encountered: domestic containers (94.54%), tires (1.64%), discarded containers (3.28%), and others (0.55%). Positive breeding sites were mostly composed of domestic containers (97.14%), followed by tires.

Typology of breeding sites in Plateau region.

Typology of breeding sites in Oueme region.
In the peridomestic environment (outdoor), domestic containers (87.59%), tires (10.95%), discarded containers (0.73%), and other breeding sites (0.73%) were encountered. Furthermore, domestic containers and tires accounted for 83.33%, and 16.47% of positive breeding sites, respectively.
In the department of Oueme, domestic containers (91.64%), tires (7.72%), discarded containers (0.32%), and other breeding sites (0.32%) were found indoors. The positive breeding sites included domestic containers (90%), followed by tires (5%) and other containers (5%). The same trend was observed outdoors.
Assessment of Epidemic Risk with the Container Index (CI), Breteau Index (BI), and House Index (HI) in the Departments of Oueme and Plateau
CI, confidence interval; C Ind, Container Index; BI, Breteau Index; HI, House Index.
In the other three rural communes (Avrankou, Ifangni, and Kétou), our results show a high epidemic risk in all three communes: Avrankou (C Ind: 71.43%; BI: 153.85; HI: 84.62%), Ifangni (C Ind: 23.24%; BI: 179.55; HI: 59.09%), and Kétou (C Ind: 83.33%; BI: 41.67; HI: 33.33%).
Prevalence of dengue virus infection in Ae. aegypti and Ae. albopictus
A total of 180 pools, including 127 for Ae. aegypti and 53 for Ae. albopictus, were analyzed. Our results showed the presence of the dengue virus in the two Aedes species in the six study communes. The combination of data from the two species (Ae. aegypti and Ae. albopictus) showed an infection rate varying between 80% (95% CI: 61.43–92.29) in Kétou and 96.67% (95% CI: 82.78–99.92) in Ifangni, with an average of 87.78% (95% CI: 82.08–92.18) for the study area (Table 2).
Dengue Virus Infection Rate in Ae. aegypti and Ae. albopictus
N, number; CI, confidence interval.
In Ae. aegypti, the infection rate varied from 76.19% (95% CI: 52.83–91.78) in Adjarra to 95.24% (95% CI: 76.18–99.88) in Ifangni and Pobè, with an average of 88.19% (95% CI: 81.27–93.24) for the study area (Table 2). In Ae. albopictus, the lowest infection rate was 66.67% (95% CI: 29.93–92.51) in Kétou and Pobè and the highest of 100% (95% CI: 66.37–100) in Ifangni, Porto –Novo, and Adjarra, with an average of 86.79% (95% CI: 74.66–94.52) for the study area. No difference was observed between the infection rate of Ae. aegypti and Ae. albopictus.
Distribution of dengue virus serotypes in the study communes
Three dengue virus serotypes have been detected. These were the DENV-1, DENV-3, and DENV-4 serotypes (Table 3 and Fig. 4). DENV-1 and DENV-3 serotypes were present throughout the study area, while the DENV-4 serotype was found only in Adjarra and Ifangni. Overall, high frequencies of the DENV-1 serotype were observed, with prevalence varying from 66.67% (47.19–82.71) in Kétou to 83.33% (65.28–94.36) in Porto-Novo and Avrankou, and an average of 77.22% (70.39–83.13) for the study area (Table 3). DENV-3 serotype was present in all the study communes with relatively low prevalence ranging from 10% (2.11–26.53) in Adjarra and Ifangni to 30% (14.73–49.40) in Porto-Novo, and an average of 17.22% (12.01–23.55) for all the study communes (Table 2).
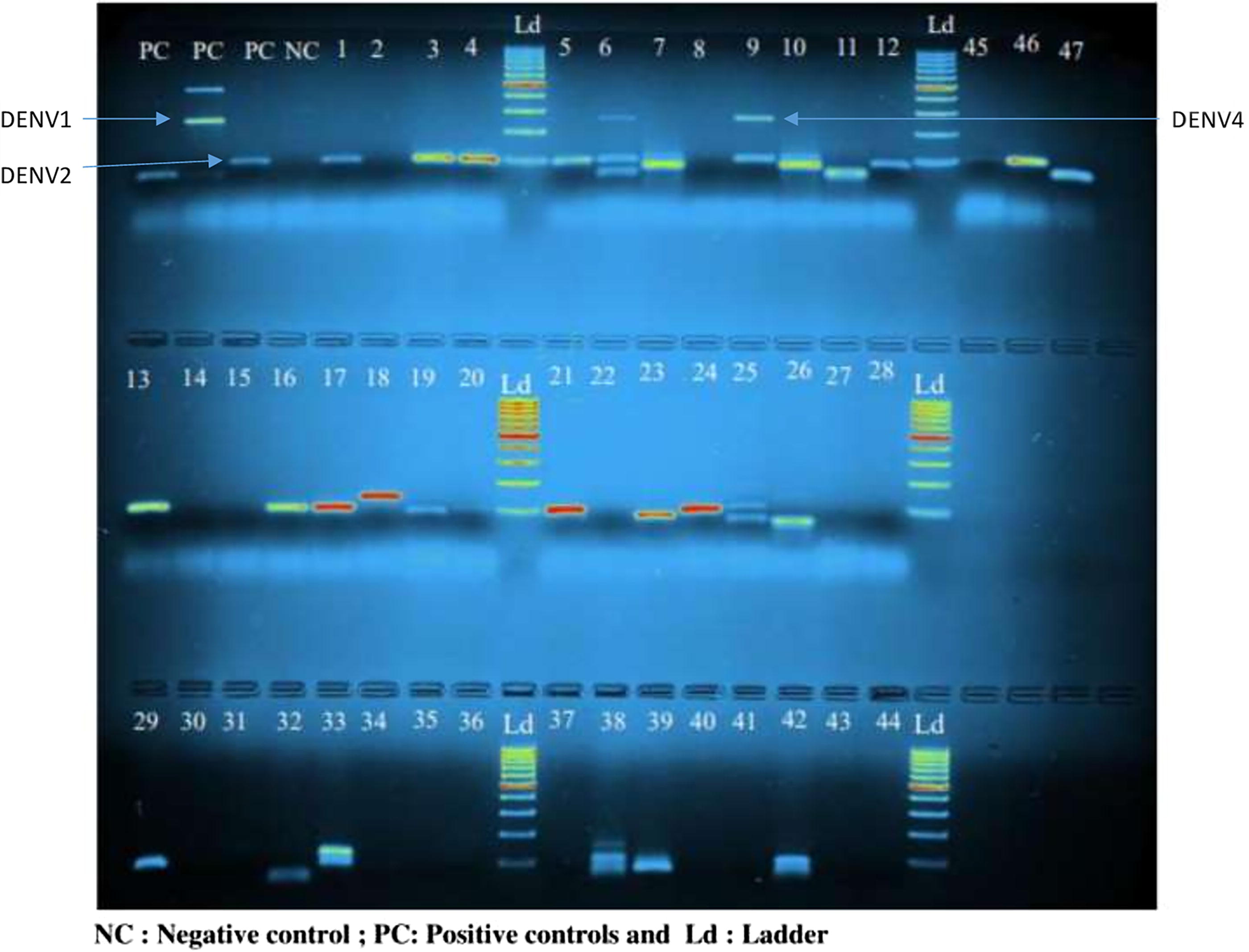
Agarose gel showing amplicons derived from RT-PCR with RNA isolated from pools of Aedes aegypti and Aedes albopictus mosquitoes.
Distribution of Dengue Serotypes in the Surveyed Districts
N_Total, number of positive pools; N_DENV 1, number of positive pools for DENV1; N_DENV 2, number of positive pools for DENV2; N_DENV 3, number of positive pools for DENV3.
Furthermore, the DENV-4 serotype was only present in Adjarra and Ifangni with a low prevalence of 3.33% (0.08–17.22), and an average of 1.11% (0.13–3.96) for all the study area (Table 3).
Distribution of dengue fever serotypes by mosquito species and study commune
The dengue serotypes were diversely distributed among the main Aedes species present in the different study communes (Table 4). In Aedes aegypti, the prevalence of the DENV-1 serotype varied from 71.43% (95% CI: 47.82–88.72) in Adjarra and Ifangni to 86.36% (95% CI: 65.09–97.09) in Avrankou, with an average of 77.95% (69.74–84.82) for all 6 study communes. DENV-3 was represented in all the study communes, with a relatively low prevalence ranging from 10% (2.11–26.53) in Adjarra and Ifangni to 30% (14.73–49.40) in Porto-Novo, with an average of 17.22% (12.01–23.55) for all the communes surveyed.
Distribution of Dengue Serotypes within Species and by Study site
N_Total, number of positive pools; N_DENV-1: number of positive pools for DENV-1; N_DENV-2: number of positive pools for DENV-2; N_DENV-3: number of positive pools for DENV-3.
On the other hand, DENV_4 was only present in Adjarra and Ifangni, with a prevalence of 3.33% (0.08–17.22) and an average of 1.11% (0.13–3.96) for all surveyed sites (Table 3). The DENV-1 serotype was the most frequent in the infected pools compared to the DENV_3 serotype (16.54%; 95% CI: 10.54–24.16) and the DENV-4 serotype (0.79%; 95% CI: 0.02–4.31) (Table 4).
Similarly, in Ae. albopictus, the DENV-1 serotype presented the higher prevalence (75.47%; 95% CI: 61.72–86.24), followed by the DENV-3 serotype (18.87%; 95% CI: 9.44–31.97), and the DENV-4 serotype (1.89%; 95% CI: 0.05–10.07) in the whole study area. Overall, no significant difference was observed between the distribution of the different serotypes within the main dengue vectors (Ae. aegypti and Ae. albopictus) (Table 4).
Discussion
Longitudinal entomological surveillance of dengue fever is an important mean for detecting and preventing epidemic outbreaks, understanding the spatiotemporal dynamics of diseases, and evaluating control interventions (WHO, 2009). In Benin, the clinical diagnosis of dengue fever is made difficult by the existence of malaria, which presents the same clinical picture as the latter. Dengue fever is therefore mistakenly confused with malaria. Furthermore, there is no effective early warning system for possible dengue epidemics. This study investigated the epidemic risk and detected the different serotypes of the dengue virus (DENV) in the populations of Ae. aegypti and Ae. albopictus.
Our results show a high infestation of domestic containers by immature stages of Aedes in all the study communes. Anthropogenic practices such as storage of drinking water, production of alcohol, and palm oil requiring the conservation of water in many domestic containers could justify the proliferation of Aedes mosquitoes (Nilsson et al., 2018). Similar results were obtained in Benin in 2020 where Aedes larvae were found in different categories of breeding sites in both domestic and peridomestic environments (Padonou et al., 2020). These results suggest that it is important to involve populations in the control of Aedes spp. vectors.
Indeed, some habits and practices of populations create breeding sites for these vectors that multiply very rapidly. These habits that favor the creation of breeding sites for Aedes spp. vectors in the domestic or peridomestic environment could be avoided if this population is aware of the risk that these vectors constitute. Overgaard et al. (2017) associated infestation of domestic containers with low frequency of cleaning. Good housekeeping would contribute to the decrease of household infestation rate by arbovirus vectors. This emphasizes the importance to involve population in implementation of vector control strategies.
Our results have showed a high epidemic risk for dengue in both departments (Oueme and Plateau). The risk was particularly high for dengue, because Aedes mosquitoes have been already found to carry the dengue virus in this area of the country, which deserve regular surveillance to prevent it from being a reservoir for dengue emergence in Benin.
Similar results showing a high epidemic risk for dengue were observed in Abomey-Calavi and Hêvié, Southern Benin (Padonou et al., 2020).
Our results showed high mosquito infection in all the study communes. This finding could be explained by the closeness of these districts to Nigeria, a country endemic to dengue fever. No difference is observed between the infection rate of Ae. aegypti and Ae. albopictus, these two mosquitoes vectors being reported as highly competent in the transmission of arboviral pathogens (Lozano-Fuentes et al., 2019). This could increase the likelihood of dengue epidemics in the study sites, especially as all the conditions (breeding sites) are available for the proliferation of these vector populations.
Similarly, several studies showed the involvement of Ae. albopictus in dengue epidemic cases in Africa (Ahmed et al., 2022; Paupy et al., 2010).
Our results revealed a high prevalence of DENV-1 followed by DENV-3, while DENV-4 was weakly represented. A similar trend was previously observed in Nigeria in the Aedes mosquito population (Isa et al., 2021) and in febrile patients (Ayolabi et al., 2019). The presence of the three dengue serotypes in the departments of Oueme and Plateau in Benin is worrying. Indeed, it has been shown that infections with different serotypes can cause a severe and potentially fatal form of dengue, due to a reinforcement of antibodies (Bhatt et al., 2013; Bravo et al., 1987). The emergence of severe dengue in tropical environments can therefore be partly attributed to the cocirculation of different serotypes (Gubler, 2002).
Unlike this study, which detected no DENV-2 infection, this serotype was found in several other West African countries, either alone (Mali and Ghana) or in cocirculation with the other serotypes (Nigeria, Senegal, Burkina, and Cote d’Ivoire) (Fagbami and Onoja, 2018; Phoutrides et al., 2011; Ridde et al., 2016). This study could have reported the variations in the seasonal circulation of the different dengue virus serotypes, which was not done and constitutes a perspective for future trials. A previous study conducted by Altaras et al. (2016) revealed that some malaria rapid diagnostic tests (RDTs) were negative, while the patient had a fever. This result highlights the need to monitor dengue infection in patients without malarial fever in our study areas, due to the high infectivity of dengue virus detected in local Aedes populations.
Overall, with the findings of this study, there is need to pay particular attention to Aedes mosquito populations by implementing an effective surveillance and alert system to prevent outbreaks of dengue epidemics.
Conclusion
The stegomyan indices have shown that there is a high risk of transmission of dengue and yellow fever in the region. These results indicate the need to pay particular attention to Aedes vectors by implementing an effective surveillance and alert system to prevent outbreaks of dengue epidemics. In addition, three dengue virus serotypes (DENV-1, DENV-3, and DENV-4) are actively circulating among the two Aedes species (Ae. aegypti and Ae. albopictus) in the departments of Oueme and Plateau. This information will enable us to have a better understanding of the epidemiology of the disease, and the risk incurred by local populations to implement effective control measures.
Footnotes
Acknowledgments
The authors thank the Beninese Government through its Ministry of Health for funding this study. The authors also thank the researchers of the Center de Recherche Entomologique de Cotonou (CREC), who supported the study through our critical discussions. Finally, the authors thank the laboratory technicians of the CREC. The authors further thank Research Square for sharing the work by making it available online as a preprint as of May 30, 2023 (![]() ). The authors thank Researchgate on which this preprint was distributed [(PDF) Detection of DENV-1, DENV-3, and DENV-4 serotypes in Ae. aegypti and Ae. albopictus, and epidemic risk in the departments of Oueme and Plateau, South-Eastern Benin].
). The authors thank Researchgate on which this preprint was distributed [(PDF) Detection of DENV-1, DENV-3, and DENV-4 serotypes in Ae. aegypti and Ae. albopictus, and epidemic risk in the departments of Oueme and Plateau, South-Eastern Benin].
Authors’ Contributions
The study was designed and its protocol written by G.G.P., A.K.K., A.S.S., and M.C.A. Data collection was performed by A.K.K., D.M.Z., A.S.S., A.S., O.O., and J.A. Molecular analyses were performed by A.K.K., A.S., J.A., L.T., and D.M.Z. A.K.K., A.S.S., A.S., G.G.P., and M.C.A. wrote the article. K.A.K., D.M.Z., and A.S.S. performed the statistical analysis of data. G.G.P., A.S., R.O., L.B., and M.C.A. provided intellectual input for the scientific quality of the content. The final version was approved by all authors.
Availability of Data and Materials
The datasets that were analyzed in this study are available from the corresponding author and the second author.
Declarations: Ethics Approval and Consent to Participate
The protocol of this study was reviewed and approved by the Institutional Ethics Committee for Health Research of the Center for Research in Entomology of Cotonou (N°06–22/CREC/CIERS-CREC/SG). All the mosquito collectors were vaccinated against yellow fever and are regularly monitored. In case of confirmed fever, they are immediately taken care of by a medical doctor.
Consent for Publication
Not applicable.
Competing Interests
The authors declare that they have no competing interests.
Author Disclosure Statement
No conflicts of interest.
Funding Information
Not applicable.