
Abstract
Background:
West Nile virus (WNV), Everglades virus (EVEV), and five species of Orthobunyavirus were isolated from mosquitoes collected in the Everglades in 2016–2017. Prior studies of blood meals of mosquitoes in southern Florida have related findings to acquisition and transmission of EVEV, St. Louis encephalitis virus, and WNV, but not the Orthobunyavirus viruses associated with the subgenus Melanoconion of the genus Culex.
Materials and Methods:
In the present study, blood-fed mosquitoes were collected in the Everglades in 2016, 2017, 2021, and 2022, and from an industrial site in Naples, FL in 2017. Blood meals were identified to host species by PCR assays using mitochondrial cytochrome b gene.
Results:
Blood meals were identified from Anopheles crucians complex and 11 mosquito species captured in the Florida Everglades and from 3 species collected from an industrial site. The largest numbers of blood-fed specimens were from Culex nigripalpus, Culex erraticus, Culex cedecei, and Aedes taeniorhynchus. Cx. erraticus fed on mammals, birds, and reptiles, particularly American alligator. This mosquito species could transmit WNV to American alligator in the wild. Cx. nigripalpus acquired blood meals primarily from birds and mammals and frequently fed on medium-sized mammals and white-tailed deer. Water and wading birds were the primary avian hosts for Cx. nigripalpus and Cx. erraticus in the Everglades. Wading birds are susceptible to WNV and could serve as reservoir hosts. Cx. cedecei fed on five species of rodents, particularly black and hispid cotton rats. EVEV and three different species of Orthobunyavirus have been isolated from the hispid cotton rat and Cx. cedecei in the Everglades. Cx. cedecei is likely acquiring and transmitting these viruses among hispid cotton rats and other rodents. The marsh rabbit was a frequent host for An. crucians complex. An. crucians complex, and other species could acquire Tensaw virus from rabbits.
Conclusions:
Our study contributes to a better understanding of the host and viral associations of mosquito species in southwestern Florida.
Introduction
Knowledge of blood-feeding patterns of mosquito species is essential to understanding acquisition and transmission of zoonotic arboviruses, such as St. Louis encephalitis virus (SLEV; Flaviviridae: Flavivirus), Eastern equine encephalitis virus (EEEV; Togaviridae, Alphavirus), West Nile virus (WNV; Flaviviridae: Flavivirus), and Everglades virus (EVEV; Togaviridae: Alphavirus), which occur in south Florida. Serologic identification of blood sources of Florida mosquitoes was initiated in 1966–1968 using precipitin tests to identify vertebrate hosts of Culex nigripalpus collected in wooded swamp habitats in Tampa and Indian River County (Edman and Taylor, 1968). Subsequent serologic studies reported blood feeding in several species in Florida (Edman, 1979; Edman, 1974; Edman, 1971). Collections of mosquitoes in Florida were from Indian River County on the mid-east coast, Tampa on the west coast, and in the Everglades near the southern tip of the state. More recently, five studies focused on blood feeding by Melanoconion species of Culex and Culiseta melanura using PCR to identify blood meals.
Wading bird species, particularly herons, were most frequently fed upon by Culex erraticus and Cs. melanura in winter in Hillsborough County, FL (Bingham et al., 2014). Culex iolambdis fed more frequently on birds, reptiles, and amphibians than on mammals in southern Florida (Blosser et al., 2016). Culex atratus and Culex pilosus were associated primarily with reptiles in Everglades National Park (Reeves et al., 2019). Rodents, particularly hispid rats, cotton mice, and black rats, were the primary hosts for Culex cedecei in the Florida Everglades (Hoyer et al., 2019; Hoyer et al., 2017; Sloyer et al., 2022). Host use by Cx. cedecei in relation to infection with EVEV and risk of infection in humans was reported in Big Cypress, Fakahatchee Strand, and other locations in southwest Florida (Burkett-Cadena et al., 2021; Sloyer et al., 2022). Burkett-Cadena et al. (2023) reported Cx. cedecei to be the only confirmed vector of EVEV. Additional studies in south Florida reported local, habitat-specific, and seasonal variations in host usage patterns for Cx. nigripalpus and Culex quinquefasciatus in Sarasota County (Hancock and Camp, 2022).
Host use by Cs. melanura relative to EEEV was reported for central Florida (West et al., 2020a; West et al., 2020b), and the importance of lizards as dilution hosts of flaviviruses by Cx. nigripalpus in Florida was presented by Reeves and Burkett-Cardena (2022).
In 2016 and 2017, we isolated seven different viruses from mosquitoes collected monthly in Centers for Disease Control (CDC) miniature light traps in the Everglades in southwestern Florida (Collier County) (Anderson et al., 2022). In the current study, engorged or partially engorged mosquitoes, which had been separated from nonblooded specimens collected in 2016 and 2017, were analyzed by PCR to determine origin of the blood. Additionally, mosquitoes were collected in an industrial recycling plant in Naples, FL, during 3 weeks in July 2017, and in the Everglades from September 2021 through October 2022. We report the host species' identity of these bloods and relate these findings to viruses infecting specific species of mosquitoes.
Materials and Methods
Mosquito collections
Mosquitoes were trapped in the Everglades, Collier County, FL in 2016, 2017, 2021, and 2022 (Fig. 1). Collections were made with CDC miniature light traps (Model 512; John W. Hock Co., Gainesville, FL) baited with compressed CO2 in 2016 and 2017. Traps were hung from a tree branch at a height of 1.0 meters above the ground in the afternoon or evening and retrieved the following morning. From September 2021 through October 2022, mosquitoes were collected in CDC traps baited with CO2 placed at heights of 1.0 and 5.0 meters above the ground, MMX traps (American Biophysics Corp., East Greenwich, RI) placed 2.5–7.5 cm above the ground, BG Sentinel traps (BioQuip Products, Rancho Dominguez, CA) baited with CO2 and BG lure, and resting shelters (Molaei et al., 2016). In 2016 and 2017, habitats included mangrove, hardwood hammock, sawgrass, and cypress swamps within or slightly east of Naples, FL (Anderson et al., 2022). In 2021 and 2022, mosquitoes were trapped in hardwood hammock (about 0.5 km west of a water and wading bird rookery), cypress swamp, and mangrove habitats, including buttonwood-edge of a coastal lake.
Mosquito collection locations in or near Port of the Islands, the Everglades, Naples, Collier County, Florida.
Buttonwood (Conocarpus erectus) belongs to the same plant family as white mangrove (Laguncularia racemosa) and occurs at the edges of mangrove swamps (Lodge, 2017).
Additionally, mosquitoes were trapped in an industrial area in Naples, FL for 3 weeks in July 2017 (Fig. 1). Traps were placed on the perimeter of the industrial site where trees and shrubbery were present. Both CDC traps baited with CO2 and BG Sentinel traps baited with CO2 and BG lure were used.
Collected mosquitoes were placed in an insulated container with cold packs, immobilized at freezing temperatures, and transferred into glass or plastic containers sealed with water and gas-proof tape. Containers were appropriately labeled and immediately stored on dry ice or in a −80°C freezer until packaged in dry ice within a Styrofoam shipping container and sent by overnight mail to the Connecticut Agricultural Experiment Station (CAES) in New Haven, CT. Mosquitoes were stored at CAES in a −80°C freezer until processing. Mosquitoes were removed from the freezer, placed on a cold platform, and each specimen was identified to species using a dissecting microscope (Burkett-Cadena et al., 2022; Darsie and Morris, 1998; Darsie and Ward, 2005; Knight and Haeger, 1971). Engorged or partially engorged mosquitoes were separated from nonblooded specimens, placed individually into 1.5-mL vials, labeled, and frozen at −80°C.
Blood meal identification
DNA was extracted from abdominal contents of individual blood-fed mosquitoes using DNAzol BD (Molecular Research Center, Cincinnati, OH) according to the manufacturer's recommendation with some modifications. Briefly, individual mosquito abdomens were homogenized with the aid of sterile polypropylene pestles (microtube pestles; USA Scientific, Enfield, CT) in 1.5-mL tubes containing 400 μL DNAzol BD solution. After adding 15 μL of proteinase K (Qiagen, Valencia, CA), the homogenates were incubated at 70°C for 10 min and centrifuged at 20,000 × g for 10 min. This was followed by transferring 400 μL of the supernatants into new microtubes, adding 3 μL Poly Acryl Carrier (Molecular Research Center), and incubating at room temperature for 3 min. DNA precipitation was carried out by the addition of 200 μL of 100% EtOH and centrifugation at 11,000 × g for 10 min. DNA pellets were washed twice with 75% EtOH, air dried briefly, reconstituted in 30 μL TE buffer (10 mM Tris-HCL [pH 8.0], 1 mM EDTA), and stored at −20°C for further analysis.
Extracted DNAs were used as templates in PCR assays. Primers were based on avian and mammalian mitochondrial cytochrome b gene sequences using established protocols (Molaei et al., 2006; Ngo and Kramer, 2003). DNAs that did not amplify with mitochondrial cytochrome b gene primers were subjected to additional PCR assays using universal vertebrate primers based on mitochondrial cytochrome oxidase I gene (Hernandez-Triana et al., 2017). PCR amplicons were then purified using the QIAquick PCR Purification Kit (Qiagen), and sequencing of both DNA strands was performed directly in cycle-sequencing reactions (Keck Sequencing Facility, New Haven, CT) with a SEQStudio Genetic Analyzer 3730xl (Applied Biosystems, Foster City, CA). DNA sequences were annotated by ChromasPro 2.1.8 (Technelysium Pty Ltd., South Brisbane, Queensland, Australia) and compared with the GenBank DNA sequence database (https:blast.ncbi.nim.nih.gov/Blast.cgi). This molecular-based assay was verified by isolating the DNA from the blood of several species, including domestic cat, gray catbird, and white-tailed deer (Molaei et al., 2006).
Data analysis
To assess the relationship between mosquito species and host species, we avoided methods that rely on approximations valid only for relatively high observation counts, like chi-squared analysis, because many mosquito species–host species pairs were observed only rarely (Haberman, 1988). First, we calculated the frequency with which each mosquito species and host species occurred in our dataset. We used these base rates to calculate the expected frequency of occurrence of each mosquito species–host species pair under the null hypothesis that these two variables were independent. We calculated the sum of squared standardized residuals for datasets simulated from this model by Monte Carlo sampling. We compared the summed squared residuals from our dataset to this null distribution to assess the significance of the mosquito species–host species relationship. Lastly, we employed Fisher's Exact Test to evaluate the significance of the observed frequency of each mosquito species–host species pair compared with the null expectation for that pair (Shan and Gerstenberger, 2017). We applied Benjamini–Hochberg multiple comparison corrections to reduce the Type I error rate across these post hoc tests.
We applied analogous methods to assess the relationship between other variable pairs of interest (mosquito species vs. host category, mosquito species and host category vs. collection season).
Results
Four hundred fifteen blood-fed specimens were tested to identify the vertebrate hosts. These were obtained from (1) 668,809 mosquitoes collected in mangrove, cypress swamp, hardwood hammock, and sawgrass habitats in the Everglades in 2016 and 2017, (2) 211,044 mosquitoes collected in 2021 and 2022 from mangrove, cypress swamp, and hardwood hammock habitats, and (3) 17,786 mosquitoes collected at the industrial site in Naples, FL in July 2017 (Fig. 1). Hosts were identified from 78.1% (n = 324) of tested blood meals. They were obtained from 7 mosquito genera comprising 13 species and 1 mosquito complex, Anopheles crucians (An. crucians, Anopheles bradleyi, and Anopheles georgianus). The largest number of specimens tested were from Cx. nigripalpus (n = 144), Cx. erraticus (n = 66), Cx. cedecei (n = 34), and Aedes taeniorhynchus (n = 29) (Tables 1 –3 and Fig. 2).

Counts of blood meals from different mosquito species. Species are grouped by genus in order of frequency. Within-genus species are ordered by frequency.
Number (Percent) of Blood Meals Identified from Nine Species or Complexes of Mosquitoes Collected in the Florida Everglades, 2016 and 2017
Fourteen mosquito specimens fed on two hosts (numbers are included in the table): three Ae. taeniorhynchus fed on Virginia opossum and on human; one Cx. erraticus fed on common box turtle and on green heron; one Cx. erraticus fed on Virginia opossum and on human; one Cx. nigripalpus fed on Virginia opossum and on cedar waxwing; one Cx. nigripalpus fed on Virginia opossum and on green heron; one Cx. nigripalpus fed on Virginia opossum and on blue-headed vireo; two Cx. nigripalpus fed on Virginia opossum and on blue-gray gnatcatcher; one Cx. nigripalpus fed on Virginia opossum and on a least bittern; one Cx. nigripalpus fed on domestic cat and on green heron; two Cx iolambdis fed on Virginia opossum and on green heron.
Number (Percentage) of Blood Meals Identified from 10 Species or Complexes of Mosquitoes Collected in the Florida Everglades, 2021 and 2022
Number (Percentage) of Mammalian, Avian, and Reptilian-Derived Blood Meals Identified from Three Species of Mosquitoes Collected in Naples FL, 2017
Mosquitoes fed on mammals, birds, reptiles, and amphibians (Tables 1–3 and Fig. 3). Five species fed almost exclusively on mammals. Ae. taeniorhynchus (n = 29), which fed on one bird, fed most extensively on humans, Virginia opossum, and white-tailed deer. Cx. cedecei (n = 34) fed most frequently on black rat, hispid cotton rat, and Virginia opossum. Mansonia dyari (n = 2) fed on Virginia opossum. Psorophora columbiae (n = 1) and Aedes aegypti (n = 1) fed on domestic cat.

Counts of blood meals from different host species. Species are grouped by major category (mammal, bird, reptile, amphibian) in order of frequency count. Within-group species are ordered by frequency. Inset shows frequency of blood meals by major host category.
Other species fed on one, two, or three different orders of vertebrates. Cx. nigripalpus (n = 144) fed on 10 different mammals and 16 species of birds, including Virginia opossum (n = 28), domestic cat (n = 23), white-tailed deer (n = 19), raccoon (n = 15), human (n = 9), and green heron (n = 11). It also fed on one American alligator. Of the 66 partially or fully fed Cx. erraticus feeding on mammals, birds, and reptiles, the most frequent hosts were white-tailed deer (n = 13), Virginia opossum (n = 10), American alligator (n = 10), and great blue heron (n = 7). Cx. iolambdis (n = 13) fed most frequently on Virginia opossum (n = 5) and green heron (n = 5). An. crucians complex (n = 21) fed on mammals, birds, and an amphibian. They fed most frequently on marsh rabbit (n = 10) and white-tailed deer (n = 6). Anopheles atropos (n = 3) fed on three species of birds. Uranotaenia lowii (n = 5) fed on three species of amphibians, particularly Cuban tree frog (n = 3). Fourteen mosquito specimens (8.7%) possessed blood taken from two different hosts in 2016 and 2017 (Table 1).
Mosquito species were correlated with hosts by Monte Carlo sampling. After calculating adjusted residuals for the frequency of each mosquito–host category pair, significant post hoc Fisher's exact tests are shown in the heat map (Fig. 4). U. lowii was highly correlated with feeding on amphibians, Cx. erraticus fed significantly on reptiles, and Cx. cedecei and Ae. taeniorhynchus on mammals. Lower observed frequencies were noted for Cx. erraticus imbibing blood from mammals, Cx. nigripalpus from reptiles, and Cx. cedecei and Ae. taeniorhynchus feeding on birds. For frequent feedings (n ≥ 10) on specific hosts, Cx. cedecei was significantly associated with black rat and with four other species of mammals. Ae. taeniorhynchus was significantly correlated with feeding on humans, Cx. erraticus on American alligator, and Cx. nigripalpus on raccoons. Lower observed frequency of feeding on American alligator was observed for Cx. nigripalpus.
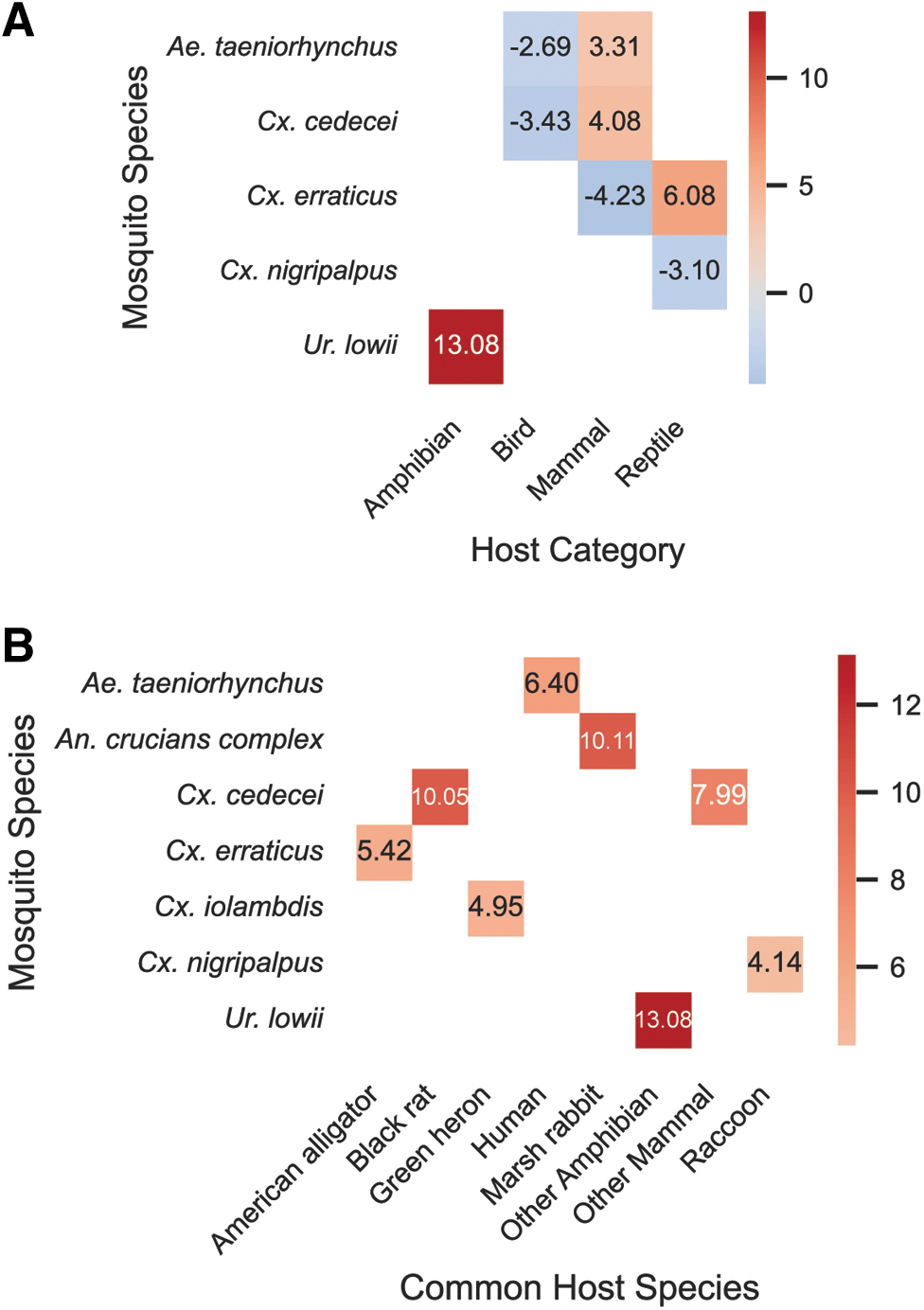
Mosquito species and host blood meal type are correlated.
Host blood meals during the dry season (November–March) and the wet season (April–October) were compared by Monte Carlo sampling (Fig. 5). Blood meals from mammals were especially common in the wet season and uncommon in the dry season. Blood meals from birds and reptiles were especially common in the dry season and uncommon in the wet season. Cx. erraticus fed more commonly in the dry season, whereas Cx. nigripalpus fed more frequently in the wetter months.
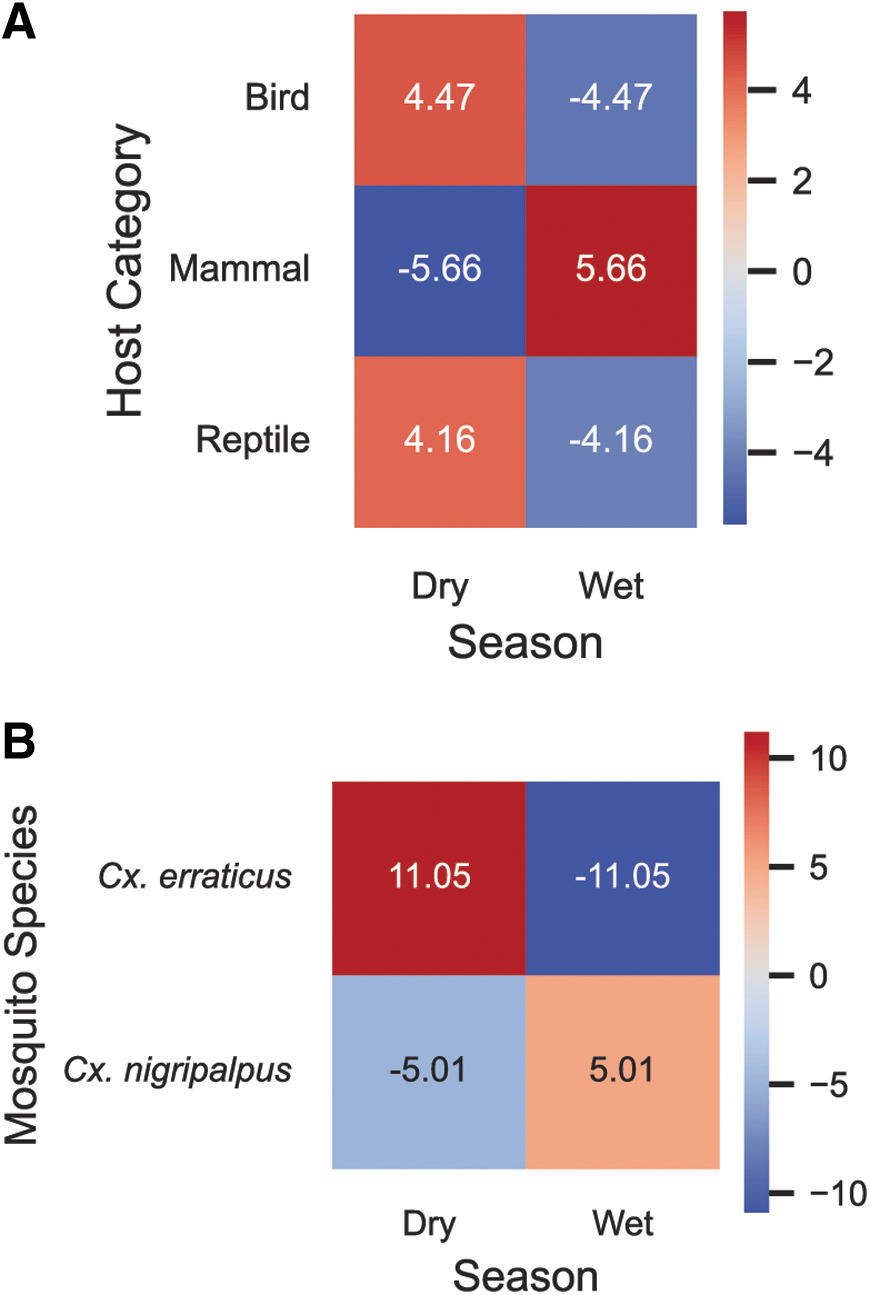
Seasonal variation in mosquito and host frequency.
Discussion
Cx. nigripalpus, which is the principal vector of SLEV and WNV in Florida (Day, 2001; Day et al., 2015) is an opportunistic feeder and imbibed blood from 10 different mammals and 16 different species of birds in our study, feeding most frequently on Virginia opossum, white-tailed deer, domestic cat, raccoon, and green heron. More specimens fed on mammals than birds during August–October in the Everglades. This larger proportion of feedings on mammals in southwestern Florida during the wet season supports the earlier findings of Edman and Taylor (1968), although Hancock and Camp (2022) did not find a seasonal shift in feeding. Reeves and Burkett-Cardena (2022) documented the extensive feeding on reptiles, particularly Anolis species.
WNV was widely distributed in the Everglades in southwest Florida in 2016 and was isolated from Cx. nigripalpus collected in four different habitats (Anderson et al., 2022). Birds, particularly passerines, are the primary vertebrate reservoirs in northeastern United States (Apperson et al., 2002; Molaei et al., 2006). Avian reservoirs for WNV in the Everglades are not known. Wading birds were the primary avian hosts in the Everglades. Twenty specimens fed on heron, ibis, and bittern; eight fed on owls; and six Cx. nigripalpus fed on passerine species. WNV is known to infect wading birds in Florida (Seixas et al., 2022), and Ciconiiformes species such as green herons may be vertebrate reservoirs for this virus elsewhere (Reisen et al., 2009).
Cx. erraticus is a generalist feeder, obtaining blood from mammals, birds, reptiles, and amphibians (Bingham et al., 2014; Egizi et al., 2018). This species has been reported to be infected with EEEV and WNV (Bingham et al., 2014; Cohen et al., 2009; Cupp et al., 2003; Mitchell et al., 1993; Unlu et al., 2010), and EVEV (Anderson et al., 2022). Wading birds have been reported as primary hosts for Cx. erraticus during winter in Hillsborough County, FL (Bingham et al., 2014). In our study in the Everglades, wading and water birds comprised 27.3% of the feedings. Mammals, reptiles, and nonwading birds comprised 48.5%, 19.7%, and 4.5% of the feedings, respectively. Blood meals from Cx. erraticus were much more common during the dry season and may be an indication of their seasonal abundance (Anderson et al., 2022).
We detected American alligator blood in 15.2% of engorged Cx. erraticus collected in the Everglades. Previously, this mosquito species was reported feeding on alligators near farms in Florida and Louisiana where these crocodilian reptiles were infected with WNV (Rodrigues and Maruniak, 2006; Unlu et al., 2010). WNV titers in alligators have been shown to be high enough to infect mosquitoes (Klenk et al., 2004), and this virus has been detected in Cx. erraticus (Bolling et al., 2005; Cupp et al., 2007; Hribar et al., 2004; Rodrigues and Maruniak, 2006). These data suggest that mosquitoes could acquire and later transmit the virus in commercial alligator farms. Our detection of alligator blood in Cx. erraticus collected in the Everglades suggests that this species could transmit WNV to alligators in the wild. However, vector competency of this mosquito species has not been documented.
Ae. taeniorhynchus, An. crucians complex, Cx. cedecei, and Ps. columbiae fed on mammals as previously reported (Edman, 1979; Edman, 1971; Hoyer et al., 2019). Ae. taeniorhynchus obtained blood meals most extensively from humans, Virginia opossum, and white-tailed deer. Cx. cedecei acquired blood meals most frequently from three species of rodents and Virginia opossum. Hoyer et al. (2019) reported that 77–100% of the blood feedings of Cx. cedecei were on rodents, particularly cotton rats. Their collections were in Everglades National Park ∼89 miles east and south of our study site. Edman (1979) recorded Cx. cedecei feeding most extensively on rodents and to lesser extents on white-tailed deer, Virginia opossum, and raccoon. Cx. cedecei is considered the primary vector of EVEV (Weaver et al., 1986), and rodents are likely the principal mammalian reservoir (Coffey et al., 2004). However, EVEV was isolated from a field-caught Virginia opossum, and the duration of viremia in experimentally infected opossums was reported to last 3–5 days (Lord et al., 1973).
Recently, multiple isolations of EVEV were made or detected from Cx. cedecei collected in the Everglades (Anderson et al., 2022; Burkett-Cadena et al., 2021; Fish et al., 2021; Hoyer et al., 2019). Our recording of Cx. cedecei feeding extensively on Virginia opossum, along with the reports of Lord et al. (1973) and Edman (1979), suggest this marsupial may also be important in the epizootiology of EVEV.
Five species of Orthobunyavirus, not known to be associated with human disease, are relatively common in the Everglades in southwestern Florida (Anderson et al., 2022). These are Gumbo Limbo (Peribunyaviridae: Orthobunyavirus), Pahayokee (Peribunyaviridae: Orthobunyavirus), Shark River (SRV; Peribunyaviridae: Orthobunyavirus), Mahogany Hammock (MHV; Peribunyaviridae: Orthobunyavirus), and Tensaw (TENV; Peribunyaviridae: Orthobunyavirus). Tensaw virus is associated with An. crucians complex (Calisher et al., 1986; Chamberlain et al., 1969), and 17 isolations were made from this complex in the Everglades in southwestern Florida (Anderson et al., 2022). In our study, Anopheles complex fed extensively on marsh rabbits, a likely reservoir for this virus (Calisher et al., 1986). Marsh rabbit and eastern cottontail rabbit were also fed upon by Ae. taeniorhynchus and Cx. nigripalpus, mosquito species from which TENV has been isolated (Wellings et al., 1972).
Gumbo Limbo virus, SRV, and MHV have been isolated from Melanoconion species (Coleman et al., 1969; Fields et al., 1969; Henderson et al., 1969) and specifically from Cx. cedecei (Anderson et al., 2022; Fish et al., 2021) and from hispid cotton rat (Coleman et al., 1969; Fields et al., 1969; Henderson et al., 1969). In our study in southwestern Florida, Cx. cedecei fed on five species of rodents, hispid cotton rat, black rat, house mouse, marsh rice rat, cotton mouse, and other mammals. Hispid cotton rats are likely viremic with these three viruses and infect Cx. cedecei in southwestern Florida. The role of the other four rodents as reservoirs is unknown.
Twenty-five mosquitoes were identified with human blood. It is possible some Ae. taeniorhynchus, Cx. erraticus, and Cx. nigripalpus acquired blood from individuals setting and retrieving traps in the field. One of the field personnel did not take personal protection precautions and reported multiple mosquito bites on several occasions. These three species have all been reported to feed on humans (Edman, 1979; Edman, 1974; Edman, 1971).
Nineteen percent of the identified bloods were from invasive vertebrates, namely black rat, house mouse, domestic cat, Eurasian collard dove, and Cuban tree frog. Not only are these non-native hosts providing sustenance for the egg production of mosquitoes, but they could also serve as possible reservoirs for arboviruses. Cigarroa-Toledo et al. (2016) reported antibodies of flavivirus in peridomestic rodents, and rats were recorded as essential for maintaining an Aedes albopictus population on Palmyra Atoll (Lafferty et al., 2018). Alternatively, some invasive species may not be competent to support development of viruses and may be dead-end hosts.
Our study contributes to better understanding of the host associations of several mosquito species in the Everglades in southwestern Florida, and in conjunction with the abundance of mosquitoes, viral infection in field-collected specimens, and vector competence, may clarify their roles in transmission of arboviruses in the region.
Footnotes
Acknowledgments
Collection of mosquitoes was carried out in Fakahatchee Strand Preserve State Park under Permit 07221414 to Durland Fish from the Florida Department of Environmental Protection and at Picayune Strand State Forest by a letter of permission from the Florida Department of Agriculture and Consumer Services. Collection of mosquitoes also was carried out in Fakahatchee Strand Preserve State Park under collecting permit 09022114A to J.F.A. from the Florida Department of Environmental Protection. The authors are grateful to Kelsey Fisher for the study sites' map. They thank Mike Owen, Charles Seither, John Shepard, Mark Kartzinel, Cora Ottaviani, Aneta Strumilowska, Tanya Petruff, and Michael Thomas for their invaluable support. Collier Mosquito Control District provided needed assistance. Garden Street Iron and Metal allowed them to collect mosquitoes. Lyle Vanwert and J. Hovey Dodge permitted the authors to trap mosquitoes on their properties.
Disclaimer
All materials reported are original. The data have not been previously published and will not be submitted to another journal.
Authors' Contributions
Designed research—J.F.A., G.M., and D.F. Performed research—J.F.A., G.M., D.F., N.K., S.B., and M.O. Reviewed and approved the article—J.F.A., G.M., D.F., P.A., N.K., S.B., M.J.M., A.B., M.O., and T.G.A. Wrote the article—J.F.A., G.M., and D.F.
Author Disclosure Statement
No conflicting financial interests exist.
Funding Information
No funding was received for this article.