
Abstract
Background:
The taxonomic status of the relapsing fever spirochete Borrelia hermsii in western North America was established in 1942 and based solely on its specific association with the soft tick vector Ornithodoros hermsi. Multilocus sequence typing (MLST) of the 16S rRNA, flaB, gyrB, glpQ, and 16S-23S rRNA intergenic spacer of B. hermsii isolates collected over many years from various geographic locations and biological sources identified two distinct clades designated previously as B. hermsii Genomic Group I (GGI) and Genomic Group II (GGII). To better assess the taxonomic relationship of these two genomic groups to each other and other species of Borrelia, DNA sequences of the entire linear chromosome were determined.
Materials and Methods:
Genomic DNA samples were prepared from 11 spirochete isolates grown in Barbour-Stoenner-Kelly-H medium. From these preparations, DNA sequences of the entire linear chromosome of two isolates of B. hermsii belonging to each genomic group and seven additional species were determined.
Results:
Chromosomal sequences of four isolates of B. hermsii contained 919,212 to 922,307 base pairs. DNA sequence identities between the two genomic groups of B. hermsii were 95.86–95.99%, which were more divergent than chromosomal sequences comparing Borrelia parkeri and Borrelia turicatae (97.13%), Borrelia recurrentis and Borrelia duttonii (97.07%), and Borrelia crocidurae and B. duttonii (97.09%). The 3′ end of the chromosome of the two GGII isolates also contained a unique intact oppA gene absent from all other species examined.
Conclusion:
Previous MLST and the chromosomal sequences presented herein support the division of the B. hermsii species complex into two species, B. hermsii sensu stricto ( = GGI) and Borrelia nietonii sp. nov. ( = GGII). We name this unique relapsing fever spirochete in honor of our late friend and colleague Dr. Nathan Nieto for his outstanding contributions to our understanding of tick-borne relapsing fever.
Introduction
The taxonomic status of the three species of relapsing fever spirochetes associated with soft ticks (Argasidae) in North America, Borrelia hermsii, Borrelia turicatae, and Borrelia parkeri, is based historically on the species of Ornithodoros tick that serves as their biological vector (Davis, 1942). Transmission of these Borrelia spp. is executed by only one species of tick, Ornithodoros hermsi, Ornithodoros turicata, and Ornithodoros parkeri, respectively (Brumpt, 1933; Davis, 1942). This unique association may result, in part, from these Borrelia spp. permanently colonizing the salivary glands of only its specific tick vector (Schwan, 2021). The 19 species of Borrelia known worldwide when reviewed in 1974 (Felsenfeld, 1974)—all named and characterized by their biological vector—remain valid species today, although the list of relapsing fever spirochetes has grown to 27 species (Barbour and Schwan, 2018; Talagrand-Reboul et al., 2018).
The inability to grow borreliae in pure culture for many years prevented their further characterization except when these spirochetes were separated from infected host blood, or when morphological features were visible microscopically in infected vertebrate and tick tissues. Kelly's (1971) development of a novel liquid medium that successfully first cultivated B. hermsii to high cell densities with growth maintained during continuous passage was a major breakthrough.
When we began our studies in 1986, only one isolate of B. hermsii was available to us, designated HS1 (Stoenner et al., 1982). This isolate originated from two infected O. hermsi ticks collected at Browne Mountain in eastern Washington at a boy scout camp where an outbreak of relapsing fever occurred in 1968 (Thompson et al., 1969). This isolate was first maintained in ticks and laboratory mice at the Rocky Mountain laboratories before Kelly's medium became available. Therefore, with a paucity of material to work with, our initial efforts were directed at establishing new isolates of tick-borne relapsing fever spirochetes in North America with an emphasis on B. hermsii. We used the culture medium developed by Kelly (1971) and later improved by Stoenner (1974) and Barbour (1984) to grow new isolates of these spirochetes from various natural and clinical sources.
We also directed efforts at developing molecular techniques to identify B. hermsii from other species of tick-borne spirochetes, methods that included DNA hybridization probes (Schwan et al., 1989), a species-specific monoclonal antibody (Schwan et al., 1992), and plasmid profiles (Schwan et al., 1995). As new isolates of B. hermsii were established, two distinctly different plasmid profiles among them were observed by agarose gel electrophoresis (Banerjee et al., 1998; Hinnebusch et al., 1998; Schwan et al., 1995). The segregation of isolates into two groups based on their plasmid composition—estimated number and size—was subsequently validated by multilocus sequence typing (MLST) targeting five chromosomal loci (16S rRNA, flaB, gyrB, glpQ, and the 16S-23S rRNA intergenic spacer) and the bipA gene encoded on the ∼200 kb linear megaplasmid. All isolates fell into one of two distinct clades of spirochetes we designated Genomic Group I (GGI), which included the type strain B. hermsii HS1T, and Genomic Group II (GGII) (Christensen et al., 2015; Fischer et al., 2009; Johnson et al., 2016; Kelly et al., 2014; Lopez et al., 2010; Porcella et al., 2005; Schwan et al., 2009; Schwan et al., 2007).
Our most recent work in western Montana identified several isolates representing a slightly divergent type of GGI spirochetes based on MLST. This led us to designate these new isolates as B. hermsii GGIB, separated from the other members of the genomic group now designated GGIA (Fischer et al., 2009; Johnson et al., 2016). Collectively, these spirochetes represent a species complex herein designated B. hermsii sensu lato. These findings demonstrate a genetic diversity of spirochetes that infect the same species of tick, O. hermsi, for their maintenance and transmission in nature. Here, we further characterize the B. hermsii sensu lato group with other species of relapsing fever spirochetes based on the DNA composition of the entire chromosome and establish Borrelia nietonii sp. nov. for those isolates assigned previously to B. hermsii GGII.
Materials and Methods
Bacterial isolates and DNA extraction
Eleven isolates of spirochetes (Table 1) were grown in Barbour-Stoenner-Kelly (BSK)-H medium (Sigma-Aldrich, St. Louis, MO, USA) and total genomic DNA samples were prepared as described (Simpson et al., 1990). Spirochetes belonging to the B. hermsii complex were identified previously by MLST (Christensen et al., 2015; Fischer et al., 2009; Johnson et al., 2016; Kelly et al., 2014; Porcella et al., 2005; Schwan et al., 2009; Schwan et al., 2007).
Species, Isolate Designation, Locality, Host, and Reference for Original Source of the Spirochetes Sequenced in this Study
San Luis Obispo County. Locations for other isolates provided in reference cited.
Location of origin unknown.
Apodemus argenteus.
We chose two isolates from each genomic group in the B. hermsii species complex and seven isolates of other species (Table 1) to compare their relatedness based on sequences of the entire linear chromosome. The two isolates representing B. hermsii GGII (MTW-2 and YOR) are herein designated B. nietonii sp. nov., while those spirochetes belonging to GGIA and GGIB (DAH and YBT-12, respectively) remain B. hermsii sensu stricto.
DNA sequencing and analysis
Chromosomal and plasmid sequences for B. hermsii DAH and Borrelia turicatae 91E135 were obtained by shotgun sequencing on an ABI 3730XL DNA sequencer base using Phred and assembled using Phrap. Finishing was performed using Consed, PCR amplification, manual editing, and a minimum consensus base quality of Q40. Sequences for the remaining nine isolates were determined with the Roche 454 FLX titanium system, 3 kb mate pair, and fragment libraries and assembled with Newbler v. 2.5, 100 × coverage or GS Assembler v. 2.5, 100 × coverage. Putative open reading frames (ORFs) in the chromosomal DNA sequences were identified with the ERGOT Genome Analysis and Discovery System (Overbeejk et al., 2003) licensed to the Rocky Mountain Laboratories.
The present analysis utilized only chromosomal sequences, which for B. hermsii HS1, the type strain belonging to GGI, represents ∼68% of the total genome (Barbour, 2016). Sequences of the remaining segmented genome that includes the linear megaplasmid and eight smaller linear and circular plasmids were not included. The chromosomal sequences were aligned with CLUSTAL W (Thompson et al., 1994), and a phylogenetic tree was constructed with the neighbor-joining method (Saitou and Nei, 1987).
Results
Sequences of the B. nietonii sp. nov. and B. hermsii sensu stricto chromosome contained 919,212 to 922,307 bp, which were similar in size to the other New and Old World species of relapsing fever spirochetes (Table 2). The % G + C contents of the B. nietonii sp. nov. and B. hermsii sensu stricto chromosome were 29.6% and 29.8%, respectively, which were slightly higher than the other species (27.5–29.5%) (Table 2). Multiple alignments of the chromosomal sequences and DNA identity values showed a clear delineation of B. nietonii sp. nov. from B. hermsii sensu stricto, as well as these two species from the others (Fig. 1; Supplementary Table S1). Identity values for the B. nietonii sp. nov. and B. hermsii chromosome were 95.86–95.99%, which were more divergent than B. parkeri SLO compared to Borrelia turicatae 91E135 (97.13%) (Table 3), Borrelia recurrentis A1 compared to Borrelia duttonii Ly (97.07%), and Borrelia crocidurae DOU compared to Borrelia duttonii CR2A (97.09%) (Supplementary Table S1).
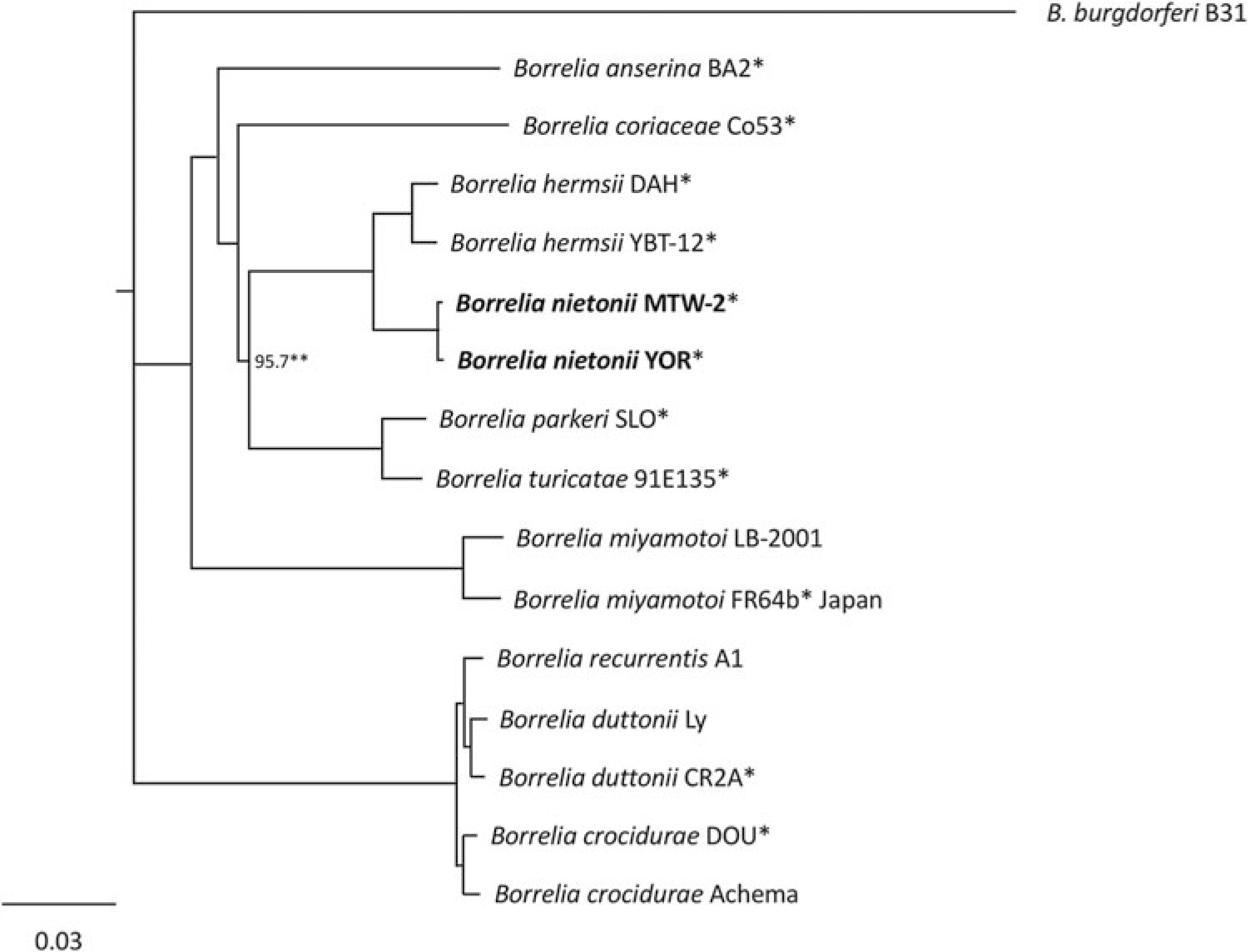
Phylogram based on alignment of chromosomal DNA sequences showing the separation of Borrelia nietonii sp. nov. (in bold) from Borrelia hermsii sensu stricto. Scale bar represents number of nucleotide substitutions per site. *Indicates sequences determined in this study. **All other branches supported by 100 bootstrap analyses.
Species, Isolate Designation, Size of Chromosome, % G + C Content, and Reference Source of the Spirochetes Compared
Some of the sequences determined by other investigators are not included in the phylogram (Fig. 1) but sequence identity values are included in the Supplementary Table S1.
DNA Sequence Identity Values (%) for the Chromosome of North American Relapsing Fever-Causing Spirochetes Associated with Soft Ticks
Analysis of the ORFs of the B. nietonii sp. nov. and B. hermsii sensu stricto chromosome showed nearly identical content and order of the ∼860–870 ORFs, although the number of small ORFs of unknown function varied. All relapsing fever spirochetes examined contained two genes, oppA1 and oppA2, that compose part of the oligopeptide transport system (Monnet, 2003), in synteny with homologous ORFs on the chromosome of B. burgdorferi (Bono et al., 1998; Fraser et al., 1997). However, both isolates of B. nietonii sp. nov. were unique from B. hermsii sensu stricto and all other relapsing fever spirochetes by having a third oligopeptide binding gene, designated here as oppA3 (1641 bp encoding a predicted 547 amino acid protein), at the 3′ terminus of the chromosome. The oppA3 gene sequences were identical in the two B. nietonii sp. nov. isolates, but only pseudogenes containing portions of oppA3 sequences were found at the end of chromosome of B. hermsii DAH, YBT-12, and Borrelia turicatae 91E135. The oppA3 of B. nietonii sp. nov. MTW-2 shared 63.2% and 73.2% DNA identity with its oppA1 and oppA2, respectively.
The annotated chromosomal sequences are deposited in the National Center for Biotechnology and Information and available in GenBank. The GenBank/EMBL/DDBJ accession numbers for the chromosomal sequences generated for this analysis are Borrelia hermsii DAH CP000048; Borrelia hermsii YBT-12 CP005706; Borrelia hermsii MTW-2 CP005680; Borrelia hermsii YOR CP004146; Borrelia turicatae 91E135 CP000049; Borrelia parkeri SLO CP005851; Borrelia coriaceae Co53 CP005745; Borrelia anserina BA2 CP005829; Borrelia miyamotoi FR64b CP004217; Borrelia crocidurae DOU CP004267; Borrelia duttonii CR2A AZIT01000001.
Discussion
Since its naming in 1942, the identity of B. hermsii has been based on the specific association of this spirochete with its tick vector O. hermsi. In our previous analyses (Christensen et al., 2015; Fischer et al., 2009; Johnson et al., 2016; Kelly et al., 2014; Porcella et al., 2005; Schwan et al., 2009; Schwan et al., 2007), spirochetes identified as B. hermsii were collected over a 50-year period (early 1960s to 2013), and included 72 isolates established in culture and eight infected samples without isolation, all characterized by PCR and MLST. Five of the 72 isolates were established by other investigators and available to us for further analysis: HS1 (Stoenner et al., 1982; Thompson et al., 1969), MAN, CON, YOR (Kurishige et al., 1990), and EST-7 (Trevejo et al., 1998). In all previous analyses cited above, these 80 samples segregated into either B. hermsii sensu stricto (N = 42) or B. nietonii sp. nov. (N = 38) established herein.
Along with the unique intact oppA3 at the 3′ end of the B. nietonii sp. nov. chromosome identified in this report, our earlier work on the purine salvage pathways of Borrelia spp. identified a 1644 bp (548 amino acid) linear plasmid-borne orf coding for adenine deaminase (adeC) in GGII spirochetes—B. nietonii sp. nov—that was a fragmented pseudogene in GGI isolates—B. hermsii sensu stricto (Pettersson et al., 2007). This additional pathway for purine metabolism by converting adenine to hypoxanthine (Canale-Parola and Kidder, 1982), along with the third OppA binding protein to potentially utilize more nutrients from the host, may give B. nietonii sp. nov. a competitive growth advantage compared to B. hermsii sensu stricto. This speculation is supported by previous attempts to isolate spirochetes from three wild-caught pine squirrels (Tamiasciurus hudsonicus) that were dually infected with both species when blood samples were collected and inoculated into BSK-H medium; only B. nietonii sp. nov. survived after one or two passages in culture (Johnson et al., 2016).
B. nietonii sp. nov. and B. hermsii sensu stricto overlap in their geographic distribution throughout much of their range in western North America (Schwan et al., 2009; Schwan et al., 2007), showing that these species are not geographically isolated variants. Both species have been found to occur sympatrically, including a tick-infested cabin where three individuals that slept in the same bed were infected, one with B. hermsii sensu stricto and the other two with B. nietonii sp. nov. (Schwan et al., 2007). O. hermsi ticks collected from a woodpile where another patient was exposed included two ticks infected with B. hermsii sensu stricto, one tick infected with B. nietonii sp. nov., and one tick dually infected with both species (Christensen et al., 2015). As mentioned above, three pine squirrels (T. hudsonicus) trapped in western Montana were also dually infected with both species at the time of their capture (Johnson et al., 2016).
Some species of mammals involved in the enzootic maintenance cycles of B. nietonii sp. nov. and B. hermsii sensu stricto may also differ, but the true diversity of hosts susceptible to infection with these two species of spirochetes requires further investigation. Burgdorfer and Mavros (1970) showed that B. hermsii sensu stricto HS1T ( = GGI) produced spirochetemias in pine squirrels (T. hudsonicus), yellow-pine chipmunks (Tamias amoenus), and meadow voles (Microtus pennsylvanicus) but not in Columbian ground squirrels (Urocitellus columbianus), bushy-tailed woodrats (Neotoma cinerea), flying squirrels (Glaucomys sabrinus), or deer mice (Peromyscus maniculatus).
Additionally, all B. hermsii sensu lato infections reported to date in wild-caught chipmunks—five species in four locations—have been genetically typed as B. hermsii sensu stricto (Christensen et al., 2015; Jones et al., 2016; Nieto and Teglas, 2014; Schwan et al., 2007; Trevejo et al., 1998). These results suggest that chipmunks may not serve as natural hosts for B. nietonii sp. nov. In contrast, as Burgdorfer and Mavros (1970) first demonstrated, deer mice (P. maniculatus) are not susceptible to infection with B. hermsii sensu stricto but, as recently shown, these mice are susceptible to infection with B. nietonii sp. nov. and exhibit high spirochetemias when infected by needle inoculation or tick bite (Johnson et al., 2016).
Conclusions
In closing, our data do not contradict the longstanding paradigm established by Davis (1942) regarding spirochete—tick specificity, but demonstrate a genetic diversity of spirochetes represented by two species, B. hermsii and B. nietonii sp. nov., both of which utilize the same tick vector O. hermsi for their enzootic maintenance in natural foci of infection. We are unaware of any other species of Ornithodoros tick serving as the vector for more than one species of relapsing fever spirochete (Barbour and Schwan, 2018; Talagrand-Reboul et al., 2018; Trape et al., 2013).
Description of B. nietonii sp. nov.
B. nietonii sp. nov. (ni.et.o'ni.i. N.L. gen. n. nietonii in honor of our late colleague and friend Dr. Nathan Christopher Nieto for his outstanding contributions to the ecology of tick-borne relapsing fever in the western United States).
This species is characterized by MLST of 38 isolates originating from Montana, Washington, Idaho, California, and British Columbia, Canada. DNA sequences of 16S rRNA, flaB, gyrB, glpQ, and IGS all distinguish this species from B. hermsii sensu stricto as do the chromosomal sequences presented here. The oppA3 gene at the 3′ terminus of the chromosome is unique and absent in all other species of relapsing fever spirochetes examined. This spirochete's size and shape are consistent with other species of the genus (Barbour and Schwan, 2018) and it binds monoclonal antibody H9724 (Barbour et al., 1986). Our samples originated from humans with relapsing fever, a domestic dog, pine squirrels (T. hudsonicus), a deer mouse (P. maniculatus), and its tick vector O. hermsi. This species is infectious in deer mice (P. maniculatus), while B. hermsii sensu stricto is not (Burgdorfer and Mavros, 1970; Johnson et al., 2016).
The type strain MTW-2T was isolated in BSK-H medium from the blood of a laboratory mouse infected when fed upon by one O. hermsi nymph collected at Mt. Wilson Observatory, at 1737 m (5700 feet) elevation, San Gabriel Mountains, Los Angeles County, California, on June 14, 2007 (Schwan et al., 2009). This isolate has been deposited in the American Type Culture Collection (number TSD-355) and the Leibniz Institute—DSMZ German Collection of Microorganisms and Cell Cultures (number DSM 115472).
Footnotes
Acknowledgments
We thank Stephen Porcella for assistance with genome sequencing during the early stages of this work, Sven Bergström for reviewing the manuscript, and Daniel Dulebohn and Denise Severson for help with shipping the type strain.
Authors' Contributions
Conceptualization: T.G.S. Data curation: T.G.S., S.J.R., and C.M. Investigation: T.G.S., S.J.R., S.M.R., D.P.B., and C.M. Resources: T.G.S. and S.J.R. Supervision: T.G.S. Writing original draft: T.G.S. Writing, editing and review: T.G.S., S.J.R., S.M.R., D.P.B., and C.M.
Author Disclosure Statement
No conflicting financial interests exist.
Funding Information
This work was supported by the Division of Intramural Research, NIAID, NIH.
Supplementary Material
Supplementary Table S1
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.