
Abstract
Background:
Chagas disease or American trypanosomiasis, caused by Trypanosoma cruzi and vectored by triatomines, affects millions of people worldwide. In endemic countries including Mexico, infections in domestic animals, such as dogs, may affect the risk of human disease when they serve as a source of infection to vectors that subsequently infect humans.
Materials and Methods:
We conducted a cross-sectional study of 296 dogs from two cities near the northern and southern borders of Mexico: Reynosa, Tamaulipas, and Tuxtla Gutierrez, Chiapas. Infection was measured based on testing of blood using T. cruzi quantitative PCR (qPCR) and up to three antibody detection assays. The StatPak immunochromatographic assay was used to screen samples and the indirect fluorescent antibody (IFA) and multiplex microsphere immunoassay (MIA) tests were used as secondary tests on all samples that screened positive and a subset of negatives. Serologic positivity was defined based on reactivity on at least two independent tests.
Results:
Of the 280 samples tested for parasite DNA, two (0.7%) were positive, one of which (0.4%) was confirmed as T. cruzi discrete typing unit TcIV. Overall, 72 (24.3%) samples were reactive for T. cruzi antibodies via StatPak of which 8 were also positive using MIA and 2 were also positive using IFA (including one of the PCR-positive dogs). Overall, nine dogs (3.4%) met study criteria of positivity based on either/both serology or PCR tests. Positive dogs were found in both regions of Mexico; five (2.7%) from Reynosa and four (3.6%) from Tuxtla Gutierrez. We found no association between infection status and state of origin, sex, age group, breed group, neighborhood, and whether other pets lived in the home.
Conclusion:
Our results re-emphasize dogs’ utility as sentinels for T. cruzi in Mexico and underscore the need for improved veterinary diagnostic tests and parasite surveillance at the household level in endemic countries.
Introduction
Caused by the protozoan parasite Trypanosoma cruzi, Chagas disease is endemic in over 21 countries in the Americas, and there are more than 60 million people at risk of infection (PAHO, 2023; Bern et al., 2011; Montgomery et al., 2014). T. cruzi is vectored by triatomine insects and currently three different cycles of transmission have been established and extensively studied: wild, peridomestic, and domestic cycles throughout the Western hemisphere (Bern et al., 2011; Breniere et al., 2016; Rassi et al., 2010; Velasquez-Ortiz et al., 2022). In North America, there are 40 known species of native triatomines (Ibarra-Cerdena et al., 2009), including 34 species of triatomines found in Mexico where three main triatomines are known to be domiciled: Triatoma longipennis, T. dimidiata, and T. pallidipennis (PAHO, 2022; Rojo-Medina et al., 2018). The most abundant species in the northeastern Tamaulipas region are Triatoma gerstaeckeri and T. protracta whereas T. longipennis and T. dimidiata are the predominant species in the southern Chiapas region (Cruz-Reyes and Pickering-Lopez, 2006; Rojo-Medina et al., 2018).
T. cruzi is divided into seven discrete typing units (DTUs), which include TcI-TcVI and TcBat (Sanchez-Guillen Mdel et al., 2006; Zingales et al., 2012). TcI is the most widely distributed strain found in Mexico where it has been identified in humans, several species of wild and peri/domestic kissing bugs, and multiple species wildlife although other DTUs have been identified in humans as well (Martinez et al., 2013; Monteon et al., 2016; Villanueva-Lizama et al., 2019). Parasites of DTUs TcI-TcVI have also been identified in kissing bugs in multiple parts of Mexico, such as Michoacan, Veracruz, and Yucatan peninsula as well as several rodent species including the west Mexican cotton rat (Sigmodon mascotensis), skunks (Mephitis macroura), and armadillos (Dasypus novemcintus) (Bosseno et al., 2009; Ibanez-Cervantes et al., 2013; Monteon et al., 2016; Ramos-Ligonio et al., 2012).
Estimates of the burden of human infection with T. cruzi in Mexico range from just over 1 million cases (PAHO, 2006) to 4 million (Arnal et al., 2019; Ibarra-Cerdena et al., 2009). Case distribution between northern and southern Mexico differs; in 2017–2020, the southern state of Chiapas was one of the states with the highest incidence of acute cases (1.8 acute cases vs. 0.8 chronic cases per 100,000 inhabitants in this period), whereas Tamaulipas had a higher number of chronic cases (4.13 chronic cases vs. 0.83 acute cases per 100,000 inhabitants) (Velazquez-Ramirez et al., 2022). In Chiapas, over 600,000 individuals are estimated to be infected with T. cruzi (Carabarin-Lima et al., 2013). Contributing to the burden of Chagas disease in Chiapas may be infected immigrants from Guatemala who cross the border while traveling north through Mexico (Carabarin-Lima et al., 2013).
Dogs share common space with humans, are a blood source for vectors, and in some epidemiological contexts can serve as T. cruzi reservoirs for transmission to humans (Estrada-Franco et al., 2006; Gurtler et al., 2007). Given that dogs often have greater exposure to vectors than humans do, their utility as sentinels for T. cruzi has been evaluated. T. cruzi infections have been reported in dogs from across the triatomine range from the United States to Argentina (Jaimes-Duenez et al., 2020; Jimenez-Coello et al., 2010; Montenegro et al., 2002; Roegner et al., 2019). The objective of this study is to estimate domestic dog exposure to T. cruzi in settings from northeastern and southern Mexico based on molecular and antibody detection while providing transparency on serologic assay discordance.
Materials and Methods
Sample collection
Dog blood samples were opportunistically collected in two regions of Mexico with known local transmission of vector-borne zoonoses (Fig. 1). Companion dogs were sampled from three residential areas adjacent to a zoological park (ZooMat) in the nature reserve called “El Zapotal” in the city of Tuxtla Gutierrez, state of Chiapas, southern Mexico. The municipality of Tuxtla Gutierrez covers an area of 412.4 km2 and has a population of about 600,000 residents according to the 2020 population census. The ZooMat borders to the South with the colonia Francisco I Madero to the West with the colonia Cerro Hueco and the colonia Ampliación Francisco I Madero. The selection method of the sampling areas for this research was intentional (purposive sampling). Owners of dogs from the residential areas were invited using a combination of periphonium (megaphone advertising) and home visits for inviting them to participate in the sampling.

Map of sampling areas in squares: Reynosa, Tamaulipas, Mexico on northern Mexican border, and Tuxtla Gutierrez, Chiapas, Mexico on southern Mexican border.
In the city of Reynosa, state of Tamaulipas, in northern Mexico, samples were collected in eight neighborhoods. The city of Reynosa, with a population of over 600,000 residents, covers roughly 3,100 km2 that borders the U.S across from Hidalgo and McAllen, Texas. In Reynosa, dog owners were recruited for the study by door-to-door visits in communities. Representative neighborhoods include “La Moderna,” consisting predominantly of small concrete houses with paved streets and basic services (drinking water, electricity, drainage); these homes are typically purchased by workers using social interest credits. The neighborhood “15 de Enero” is located on the outskirts of the city of Reynosa with the perimeter of the neighborhood bordering with wild mountains (natural areas). These houses are located in large plots with large back and front yards. Basic services are provided and most streets are unpaved. The neighborhood “La Nopalera” has houses of variable constructing materials, some of them including cement and metal laminates in walls and roofs. The roads are unpaved with no drainage but the houses have electricity and drinking water.
Given previous studies in northern and southern Mexico which concluded that 4.4–9.5% of dogs were seropositive (Arce-Fonseca et al., 2017; Balan et al., 2011; Jimenez-Coello et al., 2010), we aimed to collect at least 73 dogs per region to estimate infection prevalence with 95% confidence and 5% precision. For both regions, dog owners provided written informed consent and data was gathered on dog age, sex, and breed. Blood was collected via cephalic, jugular, or medial saphenous venipuncture into tubes containing a clot activator for serum and/or tubes with EDTA as an anticoagulant for plasma samples (Becton, Dickinson, and Company, Franklin Lakes, NJ). Rabies vaccinations were offered as an incentive for participation. All samples were kept in a cooler with ice packs until returned to the laboratory for processing. After centrifugation, aliquots of blood components were stored at −80°C for up to three months until shipping to Texas A&M University. Samples from these dogs were initially used to assess the utility of domestic dog surveillance in these regions to infer the risk of transmission of arboviruses to humans (Davila et al., 2022). Animal sampling was approved by the Texas A&M University Institutional Animal Use and Care Committee (IACUC 2021-0124D CA) and El Colegio de la Frontera Sur’s Institutional Review Board.
Molecular testing
Using a volume of 200 uL of whole blood, the DNA was extracted using E.Z.N.A. kits (Omega Bio-Tek, Norcross, GA). Samples were tested for the presence of T. cruzi using up to three PCRs. The first quantitative qPCR (qPCR) amplified a 166-bp region of satellite DNA using primers Cruzi 1 (5′-ASTCGGCTGATCGTTTTCGA-3), Cruzi 2 (5′-AATTCCTCCAAGCAGCGGATA-3′), and a Cruzi 3 probe (5′-Fam-CACACACTGGACACCAA-NFQ-MGB-3) (Duffy et al., 2013; Piron et al., 2007). Primers and probe were utilized at 0.5 µM and 0.75 µM, respectively, along with iTaq Universal Probes Supermix (BioRad Laboratories, Hercules, CA) and 5 µL of DNA for a final volume of 20 µL. For samples that tested positive on the initial assay, the T. cruzi DTU was classified by subsequent analysis of the spliced leader intergenic region (SLIR) via multiplex qPCR with probes specific to the different parasite DTUs. Reactions were run using the QIAGEN Multiplex qPCR Kit (QIAGEN, Valencia, CA) and 2 µL of DNA for a final volume of 20 µL (Cura et al., 2015). If the multiplex qPCR result was negative, a final conventional PCR was run using 121/122 [121: 5′-AAATAATGTACGG(T/G)GAGATGCATGA-3′, 122: 5′-GGTTCGATTGGGGTTGGTGTAATATA-3′] primers to amplify a 330-bp region of kinetoplast DNA to verify the presence of T. cruzi (Virreira et al., 2003; Wincker et al., 1994). A sample was considered positive if it returned a Ct value under 40 using the initial screening qPCR and at least one subsequent qPCR was positive.
Serological testing
Whole blood or plasma/serum was screened for antibodies to T. cruzi using Chagas StatPak rapid immunochromatographic assays (Chembio Diagnostic Systems, Inc., Medford, NY). This assay is validated for humans but has been used for research purposes for detecting antibodies in dogs (Dumonteil et al., 2020; Nieto et al., 2009). For research purposes, we scored the bands of reactive sample as ghost band (indicative of a very faint band) or a numerical score of 1–4 (Fig. 2) in which four is the strongest result. All reactive samples, plus 10% of randomly selected negatives, were then tested for antibodies using an indirect fluorescent antibody assay (IFA). The IFA is a validated test for detection of anti-T. cruzi antibodies in dogs run on fee-for-service basis at the Texas A&M Veterinary Medical Diagnostic Laboratory and uses serum or plasma. Samples were screened via IFA at 1:20 with subsequent twofold serial dilutions performed to determine an endpoint titer; the titer was reported as the reciprocal of the highest reactive dilution (e.g., positive at 1:1,280 = titer of 1,280). Finally, all StatPak samples plus the randomly selected negatives were also tested using a multiplex microsphere immunoassay (MIA) when remaining sample volumes allowed. The MIA used nine T. cruzi antigens (FF10, G10, LE2, Kn 107, FAB4, ATPase, Kn122, Kn80, and a whole-organism lysate) to detect specific antibodies to T. cruzi as previously described (Cooley et al., 2008; Rodriguez et al., 2023). Green fluorescent protein was added to provide a negative recombinant antigen control. The canine parvovirus antigen (VP2, MyBioSource, Inc., San Diego, CA) was included as a positive control for dogs that have been vaccinated for parvovirus. In addition, we used a Leishmania protein (K39, MyBioSource, Inc., San Diego, CA) to detect cross-reactivity. The assay was carried out in 96-well, flat bottom plates containing a mixture of antigen-bound magnetic beads and an assay buffer fixed to magnetic holders. Dog samples were diluted 1:500 with 100 µL used in each test well. After a 1-h incubation at room temperature, wells were washed thrice with 200 µL of assay buffer after which 150 µL of a 1:100 dilution of anticanine IgG/phycoerythrin conjugate (Rockland, Inc., Limerick, PA) was added to each well for additional incubation, washing, and resuspension in beads in 150 µL of assay buffer. Plates were read using a MAGPIX instrument running xPONENT software (Luminex Corp, Austin, TX) and a median fluorescence intensity (MFI) was recorded for each antigen as described (Cooley et al., 2008; Rodriguez et al., 2023). A sample was considered reactive for a given antigen if the MFI was greater than four standard deviations above the mean for that antigen based on a set of true negatives (Rodriguez et al., 2023). The criteria for categorizing a sample as T. cruzi seropositive on the MIA was included reactivity to two more T. cruzi antigens and negative on the Leishmania antigen.
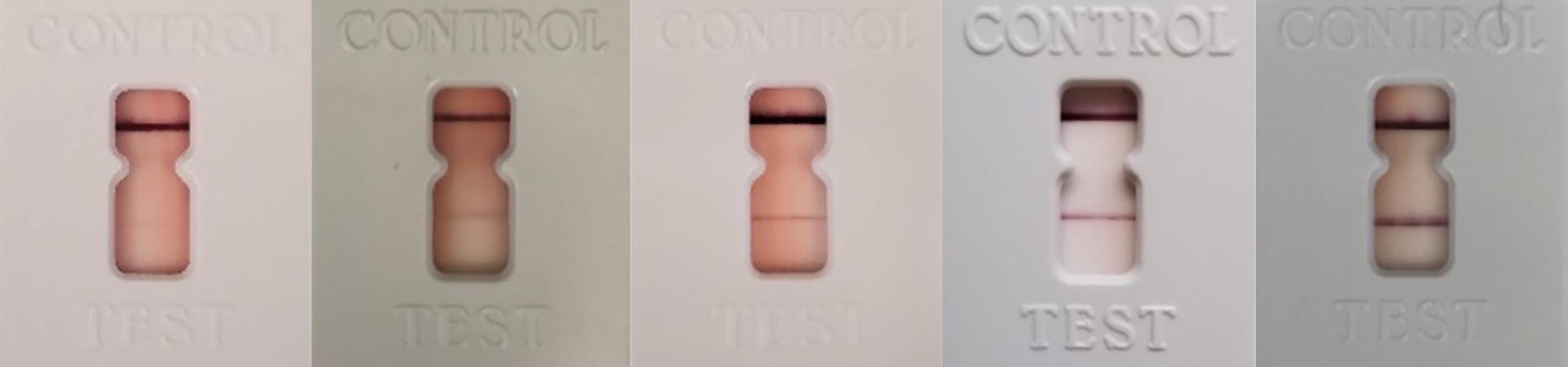
Reactive StatPak immunochromatography test results from dogs sampled between December 2018 and October 2019 in Reynosa, Tamaulipas and Tuxtla Gutierrez, Chiapas, Mexico. Scores from left: GB (ghostband), 1, 2, 3, 4.
Overall, samples were considered seropositive if they tested positive on two or more individual tests (StatPak; IFA; MIA).
Statistical analysis
Fisher’s exact test and chi-square test were used to analyze the relationship between independent variables and serostatus. Descriptive statistics were calculated and independent variables included geographical region and neighborhood, sex, age, breed group, and whether there were multiple animals in the home.
Results
Between December 2018 and October 2019, samples were taken from 296 dogs of which 184 (62.2%) came from Reynosa in Northern Mexico and 112 (37.8%) came from Tuxtla Gutierrez in Southern Mexico. There were 153 (51.9%) males and 142 (42.1%) females. The top three most common breed groups were mixed (52.2%), toy (26.1%), and terrier (8.8%). There were 188 (63.7%) over the age of 2 years, 105 (35.6%) under the age of 2 years, and 2 (0.7%) dogs of unknown ages.
Of 280 dogs tested for T. cruzi DNA, two (0.7%) were confirmed positive: a 3-year-old mix breed dog from the 15 de Enero neighborhood of Reynosa (Cruzi 123 Ct = 26.63) and a 6-year-old schnauzer from the Francisco I. Madero neighborhood of Tuxtla Gutierrez (Cruzi 123 Ct = 32.96). The dog from Reynosa tested positive on all three serological tests; it produced a StatPak band score of four, an IFA endpoint titer of 2,560, and reacted with seven recombinant T. cruzi antigens on MIA (Fig. 3). After SLIR analysis it was determined the sample was infected with discrete typing unit (DTU) TcIV (Ct = 34.7). The schnauzer sample was negative for anti-T. cruzi antibodies on StatPak, IFA, and MIA, and the DTU was not determined.

Heat map showing reactivity of serum to T. cruzi antigens from dogs in Reynosa and Tuxtla Gutierrez, Mexico between December 2018 and October.
Of 296 dogs screened for T. cruzi antibodies by StatPak, 224 were negative and 72 (24.3%) were reactive, defined as the development of a weak “ghost band” or stronger. The majority of StatPak reactive samples were scored as “GB” or 1 with only five samples scoring a two, three, or four (Table 1).
Distribution of T. cruzi StatPak Band Scores among Dogs from Chiapas, Tuxtla Gutierrez and Reynosa, Tamaulipas, Mexico Sampled between December 2018 and October 2019
GB, Ghost band.
Of the 72 StatPak positive dogs, 70 were tested on the IFA and 2 were positive (both with endpoint titers of 1:2,560). In addition, 26 StatPak negative dogs were tested on the IFA and none were positive. Of the 69 StatPak positive dogs tested on the MIA, 7 were positive. In addition, 16 StatPak negative dogs were tested on the MIA and 1 tested positive (Table 2).
Demographic and Diagnostic Data for the Nine Dogs of the Study That Met Positivity Criteria by Either Testing Positive on Trypanosoma cruzi qPCR and/or Testing Positive for Antibodies on at Least Two Independent Serologic Assays: StatPak, IFA, and MIA. Dogs Were Sampled from Reynosa and Tuxtla Gutierrez, Mexico between December 2018 and October 2019
GB, Ghost band; IFA, indirect fluorescent antibody; MIA, multiplex microsphere immunoassay; ND = not detected; F = female; M = male.
Of the 9 (3.4%) dogs that met the positivity criteria based on either/both molecular or serologic testing, 5 (2.7%) dogs were from Reynosa and 4 (3.6%) from Tuxtla Gutierrez. A single Reynosa dog tested positive on all three serological assays. Four of the neighborhoods in Reynosa contained seropositive dogs with La Moderna being the only one with multiple positive animals. All positive dogs lived in single animal homes. In Tuxtla Gutierrez, all positive dogs came from the Francisco I. Madero neighborhood. Bivariate analysis showed that there was no significant relation between seropositivity and state of origin, sex, age group, breed group, neighborhood, or whether multiple animals lived in the home.
One dog was reactive to the Leishmania (K39) antigen in the MIA (Fig. 4). This 2-year-old male mixed breed dog lived in the Francisco I. Madero neighborhood of Tuxtla Gutierrez. Overall, 79 (92.9%) of the tested dogs were reactive to the canine parvovirus (VP2) antigen (Fig. 4).
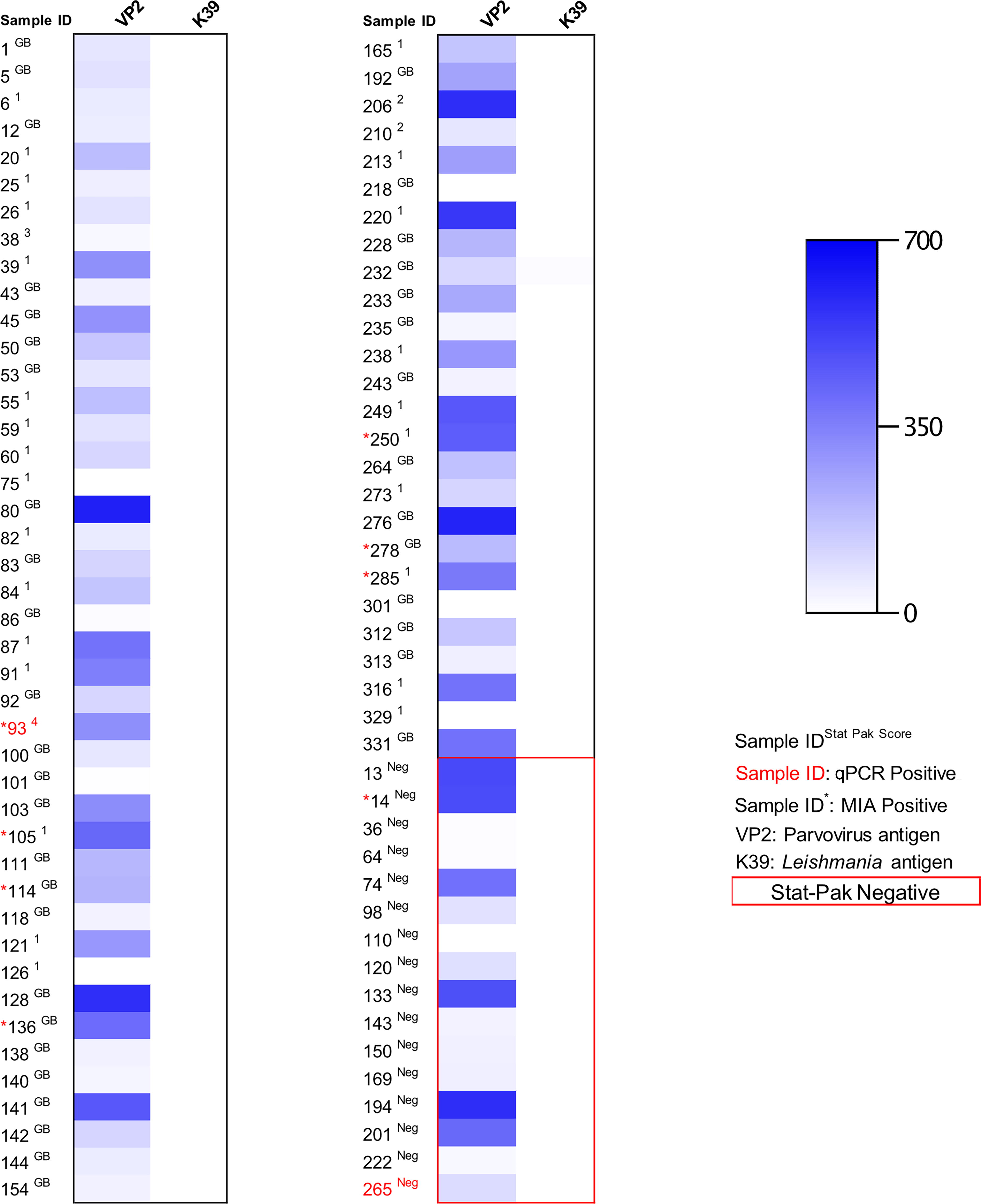
Heat map showing reactivity of serum to Leishmania control, and canine parvovirus control from dogs in Reynosa and Tuxtla Gutierrez, Mexico between December 2018 and October 2019.
Discussion
Our study provides additional support that dogs are involved in the T. cruzi transmission cycle across Mexico and may be a useful species for surveillance given their often-shared proximity to their human owners and their increased environmental exposure to parasite-transmitting triatomines. Given that parasite contamination of bite wounds—a relatively inefficient mode of transmission—accounts for nearly all human Chagas cases in Mexico, whereas dogs can become infected by direct ingestion of infected triatomines (Carabarin-Lima et al., 2013; Montenegro et al., 2002), we expected to find an overall higher level of canine exposure than found in humans within those geographic regions.
We performed the same molecular and serological testing pipeline on dog serum/plasma from northern and southern Mexico to identify infection and exposure to the parasite T. cruzi. In total, nine dogs (3.4%) were considered T. cruzi positive: one dog was positive on qPCR only, one was positive on qPCR and serology, and seven dog samples were positive on serology only (the latter requiring positivity on two independent serologic assays). However, there was discordance among serologic assay results, which hinders the interpretation of population-level prevalence studies. We used the rapid immunochromatographic assay to screen samples, subjecting only reactive samples (and a subset of negatives) to the secondary assays of IFA and MIA. Here we learned that 64 samples were only reactive on the initial StatPak assay (and thereby considered negative in our analyses), and that 0/26 and 1/16 (6.25%) of samples that screened as negative ultimately tested positive using IFA or MIA, respectively. Extrapolating these results to the broader group of STAT-pak negatives that were not subjected to the MIA (n = 206), we may have missed up to 13 MIA positive results. Taken together, the prevalence estimates reported herein are likely conservative.
Although using different detection methods in previously published studies prevents direct comparison to this study, the level of dog exposure we found is comparable to similar studies conducted in the same or nearby states of Mexico. For example, in 2010 in Tuxtla Gutierrez, researchers reported a 4.5% seropositivity rate (n = 224) among the stray dog population (Jimenez-Coello et al., 2010). In the neighboring state of Campeche, researchers found seropositivity rates among stray and owned dogs to be 5.3% (n = 148) and 9.5% (n = 114), respectively (Balan et al., 2011). Similarly, further west along the border, 4.4% of dogs from Sonora were found to be seropositive for antibodies against T. cruzi (Arce-Fonseca et al., 2017). However, the seroprevalence we report from dogs in Mexico is lower than what we previously reported from dogs north of the border in the Rio Grande Valley of south Texas, where 19.6% of dogs were seropositive on two or more serological tests and the T. cruzi qPCR positivity rate was 2.7% (Curtis-Robles et al., 2017). Further, in the north of Mexico, 9.5% of dogs from Nuevo Leon were found to have active T. cruzi infections (n = 136) (Galaviz-Silva et al., 2017) compared with the single qPCR-positive dog in our study in northern Mexico. Other studies similarly show higher levels of infections in dogs; dogs from rural communities in Veracruz had a seroprevalence of 50% (n = 34) (Benitez-Villa et al., 2022; Castillo-Neyra et al., 2015). In Costa Rica, dogs across several rural villages had a seropositivity rate of 27.7% (n = 54) which boasts a similar ecological context to both Mexico and Guatemala in terms of T. cruzi transmission (Bustamante et al., 2014; Montenegro et al., 2002). In the State of Mexico (Tejupilco municipality), the T. cruzi seroprevalence in dogs was of 21% and 7.1% in the human population. In the municipality of Toluca, historically free of T. cruzi, there was a seroprevalence of 17.5% in dogs, supporting that dogs may be sentinels for T. cruzi in these regions (Estrada-Franco et al., 2006). In comparison to some of these studies, we acknowledge the lower seroprevalence in dogs detected in our study as a limitation that may be a reflection of the more conservative diagnostic approach (e.g., requiring seropositivity on at least two independent tests). In addition, the exclusive sampling of owned dogs from urban zones where home colonization by triatomines may be less common relative to rural regions and where dogs may be kept indoors at night more commonly may have also affected seroprevalence.
Our study reinforces that the results of independent serologic tests for canine T. cruzi infections are highly discordant (Meyers et al., 2017; Tenney et al., 2014), a problem which is also evident in T. cruzi infection detection in humans. Current Pan American Health Organization and Centers for Disease Control and Prevention recommendations suggest that diagnostic testing for humans be conducted using at least two different tests, especially in countries where Chagas disease is endemic in the human population, such as Mexico (Forsyth et al., 2022). In a study from Veracruz, Mexico, 196 human samples were tested for antibodies to T. cruzi using five enzyme-linked immunosorbent assays. Positivity rate with a single test was 31.6% and decreased by more than 30% when based off the two-test approach (Guzman-Gomez et al., 2015). Using a reference cohort of 481T. cruzi-infected women (based on having at least one positive qPCR test) from Argentina, Honduras, and Mexico, Truyens et al. (2021) reported that 12%, 21%, and 72%, respectively, of T. cruzi infections failed to be detected when requiring at least two serological tests to confirm seropositivity. Further, it has been speculated that the validity of rapid diagnostic testing may differ between countries owing to genetic differences in host populations and differences between parasites (Truyens et al., 2021); this may be expected to be most apparent when using a single antigen test.
Molecular analysis of the SLIR yielded one dog sample positive for T. cruzi DNA (Ct = 26.63) that was subsequently confirmed to be DTU TcIV (Ct = 34.7). Several studies have demonstrated that TcI is the predominant strain in Mexico (Monteon et al., 2013; Ruiz-Sanchez et al., 2005), however TcIV has been found in neighboring Veracruz and in Quintana Roo at the tip of the Yucatan Peninsula (Dorn et al., 2017; Ramos-Ligonio et al., 2012). Recently, a mixed T. cruzi infection by TcI and TcII was found in a triatomine specimen). (Domínguez-Cruz et al., 2024). TcIV has been associated with all three phases of disease as well as cardiac manifestations in humans living in the Western hemisphere (Freitas et al., 2023; Magalhaes et al., 2022; Ramirez et al., 2010). The clinical status of the TcIV-infected dog in our study is unknown, but this DTU has previously been implicated in a disseminated infection leading to the death of a dog in Texas (Curtis-Robles et al., 2018).
The MIA serologic analysis revealed a very high prevalence (>90%) of dogs that had antibodies to the canine parvovirus antigen VP2. In the initial MIA multiplex assay design, the inclusion of the VP2 antigen was intended to serve as a positive control for sample integrity in regions where the canine parvovirus vaccine is widely used (e.g., in the United States, the parvovirus vaccine is recommended for all dogs regardless of geographical location). However, in Mexico, there is no parvovirus vaccine in the canine vaccination schedule, and it is used only by consultation with private veterinarians. Canine parvovirus (CPV2) infection is still seen frequently in Latin American countries (Day et al., 2020) with reported cases in Mexico (Ortega et al., 2017). Accordingly, the high level of reaction with the VP2 antigen detected may reflect a combination of some dogs that were vaccinated and/or unvaccinated dogs that have natural immunity from previous nonlethal exposure to CPV2.
Conclusion
Our study demonstrated exposure to T. cruzi among owned dogs in both the northern and southern regions of Mexico and advances our understanding of the application of diverse serodiagnostic approaches for canine surveillance programs. Given the status of Chagas disease as a neglected tropical disease and its high case burden in Latin America, adaptation of the One Health approach will increase the impact of epidemiological studies to understand the involvement of infected domestic animals in peridomestic and domestic cycles of T. cruzi transmission and emphasize the need for surveillance under the One Health approach implementation at the local, state, and/or federal level.
Footnotes
Acknowledgments
The authors appreciate field sampling assistance in Tamaulipas from Sofia Rodríguez, Irma Cobos, Cristian Delgado, Mónica Duarte, Diana Navarrate, Elisa Rodarte, Luis Sánchez, Ricardo Palacios, Adebiyi Adeniran, Jesús A. Aguilar-Durán, and María de J. López-López. The authors also appreciate field sampling assistance in Chiapas from Ivar Jasiel Torres Romero, Susana Arellano, Paola Ruiz, Daniela Mendoza, Ali Fajardo, Azucena, Katia Hernandez, Ma. Fernanda Escobar, Emiliano Escobar, Nathan Penagos, and Cristel Nandayapa.
Author Disclosure Statement
No competing financial interests exist.
Funding Information
The support came from Texas A&M University (TAMU)-CONACyT grant (No. 2018-041-1) and Texas A&M AgriLife Insect Vector Seed Grant. J.G.E.-F. was supported by grants: SIP-IPN grants 20231063, 20221576, 20226932, and 20230712.