
Abstract
Background:
Rats are an important reservoir animal for several zoonotic pathogens worldwide, including hantaviruses and Leptospira spp., which are the causative agents of hemorrhagic fever with renal syndrome, hantavirus cardiopulmonary syndrome, and leptospirosis. Although a previous study indicated a high frequency of antihantaviral antibodies in patients with acute fever in Indonesia, circulating hantaviruses and their reservoir animals in the country remain limited.
Materials and Methods:
The presence of hantavirus in rats captured in the urban area of Bogor, Indonesia, from which Leptospira spp. were isolated using PCR, followed by DNA sequencing. Immunohistochemical analyses were performed to detect hantaviral and leptospiral antigens in rat kidney tissues.
Results:
Seoul of Orthohantavirus seoulense (SEOV) RNA was detected from 24 of 80 Rattus norvegicus (30%). SEOV and Leptospira coinfection was detected in 10 of 80 rats (12.5%). Immunohistochemistry revealed that hantavirus antigens were positively stained in the interstitial capillaries and cells, whereas Leptospira antigens were stained in the luminal side of the renal tubules.
Conclusion:
This study revealed a high prevalence of SEOV and SEOV and Leptospira coinfection among rats in the urban areas of Bogor, Indonesia, indicating a potential risk of rat-borne zoonotic diseases in the area.
Introduction
Rodents carry zoonotic pathogens that pose a risk to public health (Himsworth et al. 2013, Strand & Lundkvist 2019). Rats are highly adapted to almodified and fragmented environments, such as urbanizing cities, resulting in close proximity between rats and humans, increasing the risk of human exposure to zoonotic pathogens carried by rats (Morand et al. 2015). Rattus norvegicus has lived in close association with humans for thousands of years and is one of the most invasive vertebrates, inhabiting both urban and rural habitats (Puckett & Munshi-South 2019).
Indonesia is the fourth most populated country in the world (UNFPA 2022). Urbanized and urbanizing landscapes provide opportunities for harboring rats such as R. norvegicus and Rattus rattus. Both species are well-known reservoir hosts for several zoonotic pathogens, including hantaviruses, Leptospira spp., Yersinia pestis, and rickettsiae, worldwide (Strand & Lundkvist 2019). Recently, several studies have reported the presence of rats carrying hantavirus and Leptospira spp. in Indonesia (Ibrahim et al. 2013; Koizumi et al. 2022; Lukman et al. 2019; Plyusnina et al. 2009; Plyusnina et al. 2004).
Orthohantaviruses are classified in the order Bunyavirales and family Hantaviridae by the International Committee on Taxonomy and often cause two acute febrile diseases in humans: hemorrhagic fever with renal syndrome (HFRS) and hantavirus cardiopulmonary syndrome (Noack et al. 2020). Over 50 hantavirus species have been identified, 24 of which are associated with human pathogenicity (Jiang et al. 2017). Various rodent species can act as reservoirs of hantaviruses, which have apparently coevolved with their hosts (Vaheri et al. 2013). Seoul (SEOV) of Orthohantavirus seoulense species is carried by R. norvegicus and R. rattus and inhabits areas close to human settlements (Truong et al. 2009). SEOV infections in humans often lead to asymptomatic or mild diseases; however, some may develop HFRS, which has a case-fatality rate of 1–2% (Goeijenbier et al. 2015). The signs and symptoms of SEOV infection are not specific and include fever, headache, nausea, joint pain, and cough, which are similar to the clinical symptoms of other tropical diseases such as dengue fever and leptospirosis (Kim et al. 1995). Although only 14 acute hantavirus infections, including one traveler from Germany, have been reported in Indonesia to date (Groen et al. 2002; Hofmann et al. 2018; Kosasih et al. 2011; Lie et al. 2018; Suharti et al. 2009), 11.6% (38/327) of acute febrile patients recruited in a study showed a previous hantavirus infection (positive for hantavirus IgG) during 2013–2016 in Indonesia (Lukman et al. 2019).
Leptospirosis is one of the most prevalent zoonotic diseases worldwide caused by pathogenic spirochetes of the genus Leptospira. This disease has been recognized as a major cause of acute undifferentiated fever in various parts of the world (Crump et al. 2013; Gasem et al. 2009; Mayxay et al. 2013; Mueller et al. 2014; Mørch et al. 2017; Reller et al. 2014; Wangrangsimakul et al. 2018). Rodents are the most significant maintenance hosts of Leptospira spp., which are excreted in the urine of host animals and transmitted to humans mainly through exposure to water or soil contaminated with urine (Bharti et al. 2003; Boey et al. 2019). Outbreaks of leptospirosis occur during flooding due to excessive seasonal rainfall, typhoons, or hurricanes in urban settings of developing countries in the tropics, including Indonesia (Amilasan et al. 2012; Ko et al. 1999; Trevejo et al. 1998; Victoriano et al. 2009). In Indonesia, leptospirosis is considered an underestimated health problem, with a high case-fatality rate of up to 18% (Costa et al. 2015; Indonesia KKR 2020; Victoriano et al. 2009). Recent studies indicated that more than half of patients with leptospirosis hospitalized with acute fever are clinically misdiagnosed with other infectious diseases, most commonly dengue fever. Notably, leptospirosis is the third most common etiology of nonmalarial acute fever in Jayapura (Gasem et al. 2020; Punjabi et al. 2012). Leptospirosis has been demonstrated to be a cause of nonhepatitis A–E viral acute jaundice disease, acute undifferentiated fever, and community-acquired pneumonia in Indonesia (Gasem et al. 2016; Gasem et al. 2009; Laras et al. 2002).
There are several reports of coinfections with hantavirus and Leptospira spp. in rodents: coinfections have been reported in 15.9% (7/44) in Croatia, 2.6% (9/338) in Hungary, and 0.9% (6/660) in Southwest Germany (Kurucz et al. 2018; Obiegala et al. 2017; Tadin et al. 2012). One study reported that 2 of 114 (1.8%) R. norvegicus carried hantavirus and Leptospira spp. on Flores Island, Indonesia (Mulyono et al. 2017). Leptospirosis and hantavirus infections have clinical and epidemiological similarities and represent rodent-borne zoonoses of significant public health concern.
We previously reported that Leptospira spp. were isolated from 25 of 80 R. norvegicus (31.3%) captured from local markets in the urban area of Bogor, Indonesia (Koizumi et al. 2022). In this study, we conducted additional gene detection of hantaviruses and investigated coinfections with Leptospira spp. and hantaviruses in R. norvegicus living in urban habitats. Furthermore, we observed coinfections using immunohistochemistry (IHC).
Materials and Methods
Sample collection
Ninety small mammals, 80 R. norvegicus, and 10 Suncus murinus were trapped and collected from four local markets, Anyar, Bogor, Jambu Dua, and Merdeka, in Bogor, Indonesia, in October 2019, as previously described (Huy et al. 2021;). Rats were humanely euthanized by isoflurane inhalation, as recommended by the American Veterinary Medical Association guidelines. Immediately after euthanization, liver and one kidney from an individual rat were collected and stored at −80°C until use for gene detection and IHC, respectively, whereas another kidney was used for culture of Leptospira spp. (Koizumi et al. 2022).
Detection of the hantavirus gene
RNA was extracted from the liver tissues using the ISOSPIN Cell and Tissue RNA Kit (Nippon Gene), following the manufacturer’s instructions. cDNA was synthesized from total RNA using the PrimeScript RT reagent kit (Takara, Japan). The cDNA samples were screened by PCR using Go Taq and primers targeting a conserved domain of the L genome segment of hantaviruses (Muthusinghe et al. 2021). The HAN-L-F2 (5'-TGCWGATGCHACIAARTGGTC-3') and HAN-L-R1 (5'-AACCADTCWGTYCCRTCATC-3') primers were used for the first reaction, followed by seminested amplification using the HAN-L-F2 and HAN-L-R2 (5'-GCRTCRTCWGARTGRTGDGCAA-3') primers. Both PCRs included 10 min of initial denaturation at 95°C, followed by 35 cycles of 95°C for 30 s, 55°C for 30 s, and 72°C for 30 s, and a final extension at 72°C for 7 min. Amplicons of the correct size were purified using NucleoSpin Gel and PCR Clean-up (Takara, Japan), followed by Sanger sequencing to confirm the viral species. The nucleotide sequences were deposited in a DDBJ database (accession no. LC792235∼LC792258).
Immunohistochemistry
We selected two kidney samples each from hantavirus gene positive and/or Leptospira positive samples. Kidney tissues stored at −80°C were transferred to 10% neutral buffered formalin for fixation and embedded in paraffin. Subsequently, 2-µm consecutive sections were stained with hematoxylin and eosin and subjected to IHC for Leptospira and hantavirus antigens. For IHC, the sections were de-paraffinized and incubated in methanol containing 10% hydrogen peroxide for 5 min to block endogenous peroxidases. Antigen retrieval was performed by heating the sections in citrate buffer (pH 6.0) at 121°C for 10 min. The sections were blocked with 8% skimmed milk in Tris-buffered saline and subsequently incubated with mouse monoclonal anti-Leptospira antibody (1:10) (Shiokawa et al. 2016) or rabbit polyclonal antihantavirus antibody (1:500) (Pattamadilok et al. 2006) at 4°C overnight. Immunolabelled antigens were visualized using the EnVision+ system (Agilent, Santa Clara, CA) and reacted with 0.05% 3′,3-diaminobenzidine and 0.03% hydrogen peroxide in Tris–hydrochloric acid buffer. Sections were then counterstained with hematoxylin.
Statistical analysis
The association between the prevalence of Leptospira spp. and SEOV infection was analyzed using χ2 test.
Ethical clearance
All experimental animal procedures herein performed were in accordance with the guidelines and approval of the Institutional Animal Care and Use Committee, Graduate School of Agricultural and Life Sciences, The University of Tokyo (approval no. P22-098).
Results
Coinfection with Leptospira spp. and SEOV
Among the 80 R. norvegicus, 24 (30%) were positive for SEOV RNA (Table 1), whereas L. borgpetersenii and L. interrogans were isolated from 4 and 21 rats, respectively (Koizumi et al. 2022). Four and 6 of the 21 rats captured at the Bogor market were positive for Leptospira spp. and SEOV, respectively. In the Anyar market, 18 and 15 of 46 rats tested positive for Leptospira spp. and SEOV, respectively. In the Merdeka market, two of six rats were positive for SEOV, and there were no positive results for Leptospira spp. In the Jambu Dua market, three and one isolates were positive for Leptospira spp. and SEOV, respectively. Coinfections were detected in 10 of 80 rats (12.5%). There was no significant association between sole infection with Leptospira spp. and SEOV or coinfection with Leptospira spp. and SEOV in rats. No hantavirus RNA was detected in S. murinus liver samples.
Results of Leptospira Isolation and Seoul Orthohantavirus (SEOV) RT-PCR Analyses
Koizumi et al. (2022).
Number of positives (positivity rate).
Li, Leptospira interrogans; Lb, Leptospira borgpetersenii.
Histopathology in kidney
IHC revealed Leptospira and hantavirus antigens in consecutive kidney sections (Fig. 1B and C). The kidney tissues did not show marked inflammatory lesions in Leptospira and hantavirus IHC-positive tissues (Fig. 1A, D, and G). The luminal side of the renal tubules was positively stained for Leptospira antigens (Fig. 1B and E). Interstitial capillaries and cells were positively stained for hantavirus antigens (Fig. 1C and I).
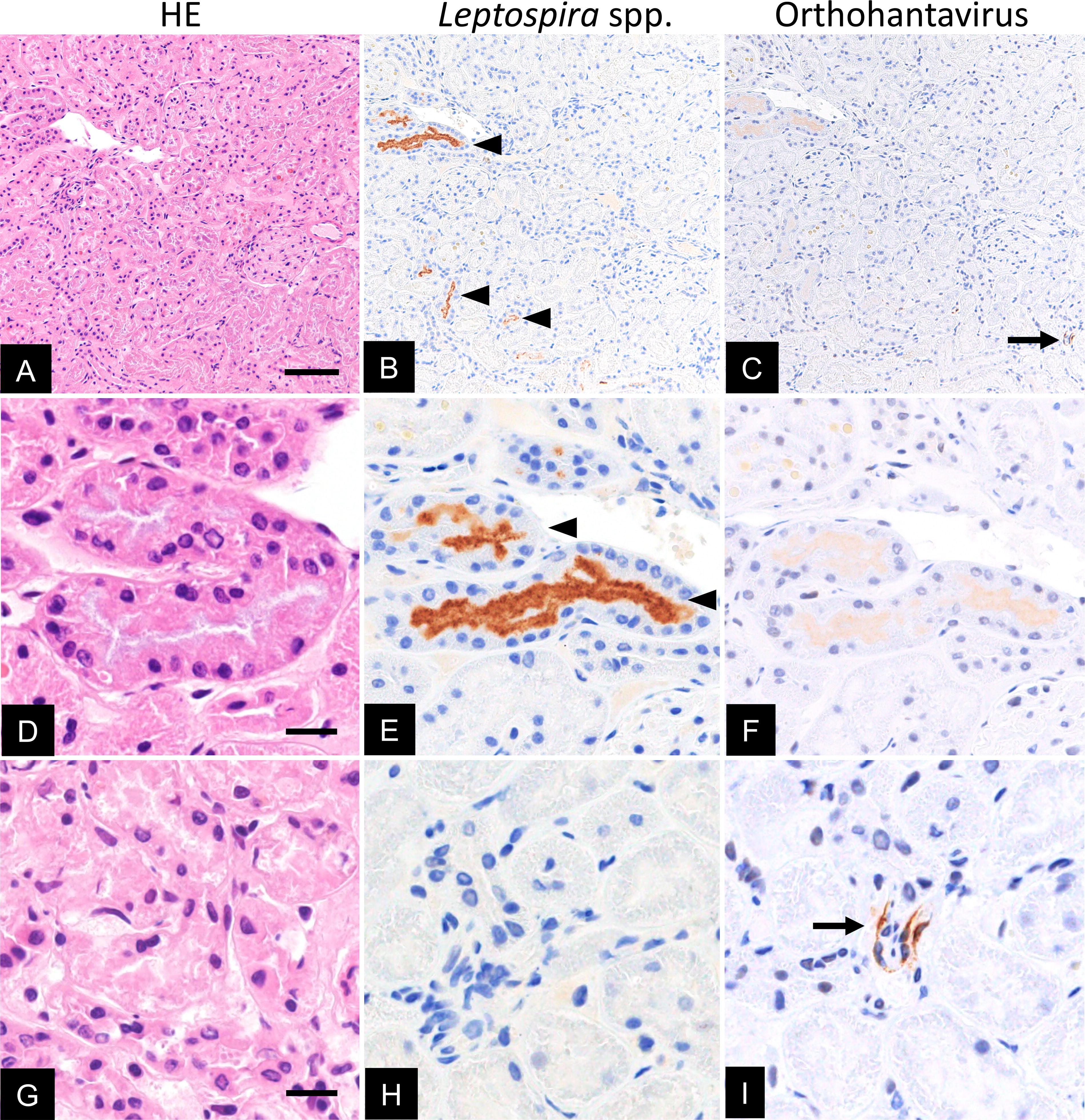
Histopathology of natural coinfections with Leptospira spp. and orthohantavirus in R. norvegicus. The kidney was stained with hematoxylin and eosin (HE; scale bar: 200 μm
Discussion
This study revealed a high SEOV carriage rate (30%) in R. norvegicus captured in Bogor, Indonesia (Table 1). A high carriage rate of hantaviruses in rodents has been reported using IgG ELISA and RT-PCR in Indonesia, with 33.9% in the Thousand Islands, 33% in Jakarta, and 10.2% in Maumere Flores Island (Hadi & Penyakit 1992; Ibrahim et al. 2013; Lukman et al. 2019). There are 14 reported cases of acute hantavirus infections in humans in Indonesia, including a traveler returning to Germany from a recent visit to Sulawesi, Indonesia (Groen et al. 2002; Hofmann et al. 2018; Kosasih et al. 2011; Lie et al. 2018; Suharti et al. 2009). Among them, 10 cases showed strong positive reactivity with the Puumala virus (PUUV) serotype by immunofluorescent assay; however, unexpected cross-reactivity among hantavirus serotypes and the potential existence of PUUV in Indonesia remains unclear (Lukman et al. 2019). One case was strongly positive for anti-Hantaan virus by ELISA, although SEOV RNA was detected in rodents trapped around the patient’s house. Three recent patients, including the German traveler, were diagnosed with SEOV infection using RT-PCR. These studies indicated that hantaviruses, predominantly SEOV, are endemic to Indonesia. It is well known that Leptospira spp. and hantavirus are excreted via urine from maintenance hosts of rodents. Therefore, kidney tissues are primarily used for gene detection and pathogen isolation. However, hantavirus antigens have been equally detected in the lungs, heart, liver, and kidneys (Noack et al. 2020). In this study, we conducted gene detection of hantaviruses from liver tissues because we used one kidney for Leptospira isolation, whereas another kidney was used for IHC studies.
This study revealed coinfection with Leptospira spp. and hantavirus in 12.5% (10/80) of R. norvegicus in Bogor. The coinfection rate was consistent with a previous study from Croatia (15.9%; 7/44); however, higher than those from Hungary (2.6%; 9/338) and Germany (0.9%; 6/660) (Kurucz et al. 2018; Obiegala et al. 2017; Tadin et al. 2012). This rate was also higher than that reported in a previous study conducted in Flores Island, Indonesia (1.8%; 2/114) (Mulyono et al. 2017). Further studies are needed to clarify the influence of coinfection with these pathogens in rats, such as the number of pathogens excreted in the urine of coinfected rats. In human cases, 7 of 31 acute severe leptospirosis cases were positive for both Leptospira spp. and SEOV in Sri Lanka (Sunil-Chandra et al. 2015). Although there is no information on whether coinfection enhances disease severity, coinfection in humans may affect the outcome of the disease, and coinfection in rats poses a potential risk.
In IHC, both Leptospira and hantavirus antigens were observed in several fields under low magnification, whereas both antigens did not merge and were not in close proximity to each other (Fig. 1B and C). In rats naturally infected with SEOV, mild inflammation was observed in the lungs; however, not in the spleen and kidney tissues (Maas et al. 2019). Renal colonization by Leptospira spp. in rats results in a lack of histopathology (Athanazio et al. 2008). The kidney tissues coinfected with Leptospira and hantavirus in this study did not show inflammatory lesions upon hematoxylin and eosin staining (Fig. 1A, D, and G). This could be due to the reservoir hosts of Leptospira spp. and hantavirus, differences in age, and prolonged exposure to Leptospira and hantavirus. During leptospiral infection in renal proximal tubule epithelial cells, several Leptospira strains induce cadherin extracellular domain shedding to reduce cell–cell adhesion (Sebastián et al. 2021). Additionally, orthohantavirus infection induces the disengagement of adherens junctions through the internalization of cadherin (Krautkrämer et al. 2011). Although Leptospira spp. and orthohantavirus colonize different cells in the kidney tissues, coinfection may be a mutually beneficial relationship for both pathogens.
This study revealed a high prevalence of SEOV and coinfection with Leptospira spp. and SEOV in R. norvegicus captured in an urban area of Bogor, Indonesia. Even though coinfection was caused by accidents in areas of high prevalence of Leptospira spp. and hantavirus in rats, further studies focusing on histopathological analysis are required to gain insights into assessing the correlation of rodent-borne zoonotic pathogens in maintenance hosts.
Footnotes
Acknowledgments
The authors thank the staff of the Indonesian Research Center for Veterinary Science and Bogor Municipality Food Security and Agriculture Service for their assistance.
Author Disclosure Statement
The authors declare no conflicts of interest.
Funding Information
This study was partially supported by the Science and Technology Research Partnership for Sustainable Development (SATREPS) (