
Abstract
Background:
In urban Thailand, arboviral infections dominate diagnoses of acute undifferentiated fevers (AUFs) owing to their well-defined epidemiology and characteristic clinical presentations. However, rickettsial diseases, also endemic in this setting, remain under-recognized owing to challenges in early detection.
Objective:
This study aimed to identify potential rickettsial infections among patients with AUF in Bangkok and vicinity utilizing leftover nucleic acid extracted from serum samples from patients initially suspected of but negative for arbovirus infections.
Materials and Methods:
A total of 609 nucleic acid samples were screened for rickettsial bacteria using real-time PCR, targeting the 17-kDa common antigen gene of Rickettsia spp. and the 47-kDa gene of Orientia tsutsugamushi.
Results:
Nine samples were positive for Rickettsia spp. and two were positive for O. tsutsugamushi. DNA sequence and phylogenetic analyses based on partial 17-kDa antigen and citrate synthase (gltA) genes identified the Rickettsia-positive samples as R. typhi in eight cases and R. felis in one case. Analysis of the 56-kDa type-specific antigen gene identified the two O. tsutsugamushi isolates as Gilliam-related genotypes. Although rickettsial diseases typically present with mild symptoms, two patients with R. typhi infection (murine typhus) developed respiratory distress syndrome, highlighting the potential for rare but serious complications.
Conclusion:
This study underscores the critical importance of differential diagnosis and prompt, effective intervention to prevent complications in suspected cases.
Introduction
Acute undifferentiated fevers (AUFs) pose substantial diagnostic challenges in tropical and subtropical regions, including Thailand, because of their diverse etiologies and overlapping clinical manifestations, such as fever, headache, myalgia, nausea, and thrombocytopenia. However, the disease spectrum varies geographically. Although urban centers such as Bangkok experience a higher burden of dengue fever, followed by murine typhus, scrub typhus is a leading cause of AUFs in rural areas, particularly in northern Thailand (Luvira et al., 2019; Wangrangsimakul et al., 2020).
Murine typhus is caused by the bacteria Rickettsia typhi and is typically spread via bites from infected fleas, especially the Oriental rat flea (Xenopsylla cheopis) (Azad et al., 1997). Scrub typhus, by contrast, is caused by Orientia tsutsugamushi and transmitted through the bites of infected trombiculid mites (Chen et al., 2022). Interestingly, as pet ownership becomes more common, cat-flea rickettsiosis, caused by R. felis and potentially transmitted by cat fleas (Ctenocephalides felis), has gained recent attention owing to its potential zoonotic risk (Pérez-Osorio et al., 2008).
Owing to their intracellular nature, it can be difficult to detect disease-causing bacteria in the early phase of infection. Thus, diagnosis of rickettsial diseases is mainly based on clinically compatible symptoms and laboratory findings in consideration of geographic location. Empirical antibiotic treatment is usually started before confirming an infection through convalescent serological testing (Stewart and Stewart, 2021). Although the diseases typically present with mild symptoms, delayed diagnosis, especially in elderly patients, can lead to severe complications such as respiratory distress syndrome or even mortality (Sakamoto et al., 2013; Vaart van der et al., 2014).
To explore the diagnostic landscape of AUFs, this study utilized real-time and nested polymerase chain reaction (PCR) assays to investigate the potential presence of Rickettsia in leftover nucleic acid samples derived from acute-phase serum of patients residing in Bangkok and vicinity who were initially suspected of having an arboviral infection but tested negative. Bacteria were classified by species and genotype based on DNA sequencing and phylogenetic analysis. Although limitations affect the use of patient-derived serum for Rickettsia detection, early identification, even shortly after disease onset, can facilitate effective treatment and potentially prevent serious complications.
Materials and Methods
Nucleic acid samples
The 609 samples used in this study were obtained from leftover nucleic acids extracted from acute-phase serum samples of patients with suspected arbovirus infection and collected between July 2020 and April 2023 at hospitals in Bangkok and vicinity (Bangpakok Hospital Group, Chulalongkorn Hospital, and Banphaeo General Hospital). These samples had previously tested negative for chikungunya, dengue, and zika viruses. Data regarding demographics, travel history, clinical presentation, and laboratory findings were collected along with the samples and recorded in a coded format. There was no direct patient contact and all data remained confidential. The samples were then further analyzed for the presence of Rickettsia DNA. The research proposal was approved by the Institutional Review Board (IRB) of the Ethics Committee of the Faculty of Medicine, Chulalongkorn University, Thailand (IRB No. 0712/66). The IRB of the Ethics Committee for Human Research granted a waiver for written informed consent because all clinical specimens were anonymized.
Molecular detection of Rickettsia spp. and O. tsutsugamushi DNA
Real-time PCR was employed to screen for Rickettsia spp. and O. tsutsugamushi utilizing fluorophore-labeled hybridization probes and specific primers. The primers R17K128F2, R17K238R, and R17K202TAQP targeted the 17-kDa common antigen gene of Rickettsia spp. (111 bp), whereas the primers OT47kDaF, OT47kDaR, and OT47kDaP targeted the 47-kDa gene of O. tsutsugamushi (118 bp) (Jiang et al., 2012; Tantibhedhyangkul et al., 2017). The reactions were performed in a total volume of 15 µL, consisting of 12 µL of SensiFAST reagent (Bioline, London, UK), 0.2 µM of each primer and probe, 3 µL of nucleic acid sample, and nuclease-free water. Amplification was carried out using a Light Cycler® 480 II instrument (Roche, Basel, Switzerland) under the following conditions: 95°C for 5 min, followed by 40 cycles of 95°C for 15 s and 60°C for 30 s.
To identify the species of Rickettsia, a partial DNA fragment of the 17-kDa common antigen gene (369 bp) was amplified by conventional PCR conducted on a Mastercycler® pro with vapo.protect™ (Eppendorf, Hamburg, Germany) with specific primers R17K128F2 and R17k-Rr2608Rnew (Taylor et al., 2016). Amplification of the citrate synthase (gltA) gene (382 bp) was then achieved through nested PCR using the specific primers CS1d-F and CS1273r in the first round, followed by Rp877p and Rp1258n in the second round (Roux et al., 1997).
To identify the genotype of O. tsutsugamushi, a partial DNA fragment of the 56-kDa type-specific antigen (TSA) gene (789 bp) was amplified using semi-nested PCR. Specific primers were used: F584-forward and RTS9-reverse in the first round, followed by RTS8-forward and RTS9-reverse in the second round (Rodkvamtook et al., 2013).
The reactions were prepared in a total volume of 25 µL, consisting of 12.5 µL of AccuStart™ II PCR SuperMix (2×) (Quantabio, Beverly, MA), 0.2 µM of each primer, 3 µL of DNA template, and nuclease-free water. Amplification was then performed under the following conditions: 95°C for 5 min, followed by 40 cycles of 95°C for 30 s, 55°C for 30 s, and 68°C for 30–60 s, with a final extension at 68°C for 2 min.
PCR amplicons of the genes were visualized using gel electrophoresis. The nucleotide sequences of the DNA fragments were verified using Sanger sequencing performed by 1st BASE, Selangor, Malaysia, followed by analysis with the DNAMAN program (Lynnon Biosoft, San Ramon, CA). The sequences of the probes and primers used are listed in Table 1.
Primers and Probes Used for Molecular Detection of Rickettsia spp. and Orientia tsutsugamushi
TSA, type-specific antigen.
Phylogenetic analysis
A maximum-likelihood tree of the nucleotide sequences was constructed using MEGA version 11 (https://www.megasoftware.net/).
Results
Detection of rickettsial DNA
Employing a real-time PCR assay targeting the common rickettsial 17-kDa antigen gene and the 47-kDa outer membrane protein antigen of O. tsutsugamushi, 9 of the 609 samples (1.48%) were identified as positive for Rickettsia spp., including isolates AB3169, AB3572, AB3658, AB3672, AB3767, AB3775, AB3799, AB4198, and AB4580. Additionally, 2 of the 609 samples (0.32%) were identified as positive for O. tsutsugamushi, including isolates AB3696 and AB4595.
Further identification of the rickettsial species was achieved by amplifying partial DNA fragments of the 17-kDa common antigen (369 bp) and citrate synthase (gltA) (382 bp) genes from the Rickettsia-positive samples. For O. tsutsugamushi—positive samples, the 56-kDa TSA gene (789 bp) was amplified (Supplementary Fig. S1). The nucleotide sequences were deposited in the GenBank database under accession numbers OR567470–OR567478 for the 17-kDa antigen gene, OR567479–OR567487 for the gltA gene, and OR567488–OR567489 for the 56-kDa TSA gene.
Phylogenetic analysis
To identify the species of the Rickettsia isolates obtained in this study, nucleotide sequences of the 17-kDa antigen and gltA genes were aligned with those of recognized Rickettsia species deposited in the GenBank database using MUSCLE implemented in MEGA11. Phylogenetic trees were constructed for both the 17-kDa antigen and gltA genes using the maximum-likelihood method and Tamura 3-parameter model, with 1,000 bootstrap replicates.
Of the nine Rickettsia-positive samples, eight were identified as R. typhi based on the phylogenetic analysis. These isolates included AB3169, AB3658, AB3672, AB3767, AB3775, AB3799, AB4198, and AB4580. The partial 17-kDa antigen gene (363 bp) and gltA (306 bp) gene sequences of these Thai isolates exhibited 100% and 99.7–100% identity, respectively, among themselves and 100% and 99.0–99.7% identity, respectively, with R. typhi reference strains. Isolate AB3572 was identified as R. felis based on the sequences of the 17-kDa antigen and gltA genes, which exhibited 100% identity with recognized R. felis sequences in the database (Fig. 1 and Supplementary Tables S1-S2).

Phylogenetic analysis of Rickettsia spp. isolates based on the partial nucleotide sequences of the 17-kDa antigen gene
To characterize the genotype of O. tsutsugamushi isolated in this study, the nucleotide sequences of the 56-kDa TSA gene were aligned with those of other strains isolated in Thailand and other countries. The phylogenetic analysis revealed that both AB3696 and AB4595 were related to the Gilliam prototype and shared 100% identity with isolate MH437, which was isolated in Maehongson Province, Thailand (accession number HM777461) (Fig. 2 and Supplementary Table S3).
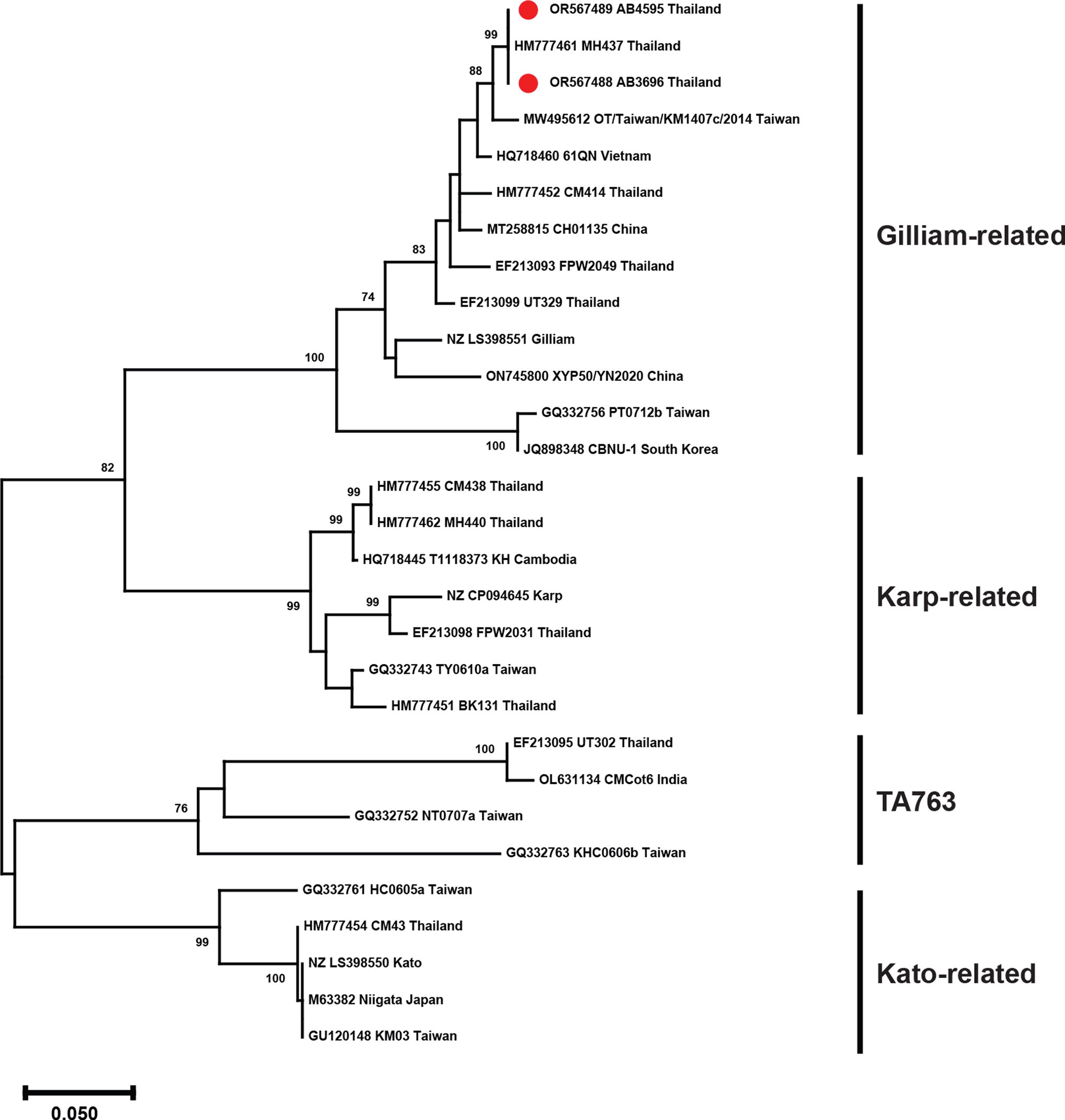
Phylogenetic analysis of Orientia tsutsugamushi isolates based on partial nucleotide sequences of 56-kDa type-specific antigen (TSA) gene. Trees were generated using the maximum-likelihood method based on the Tamura 3-parameter model with a discrete gamma distribution (T92+G). GenBank accession number, isolates, and country are presented for each sequence. Bootstrap values >70% are shown on the branch nodes. The scale bar denotes the number of substitutions per site. Thai strains are indicated by red dots.
Clinical characteristics
Murine typhus, caused by infection with R. typhi, was identified in eight patients spanning a wide age range, from 5 to 69 years old, between September 2020 and April 2023. This suggested that people of all age groups are susceptible to R. typhi throughout the year. Of the eight murine typhus cases, four were identified in Bangkok residents and four in residents of Samut Prakran. Typical symptoms were observed in the murine typhus patients, including fever (body temperature 37–39.6°C), headache, myalgia, fatigue, nausea, vomiting, and elevated liver enzyme (aspartate aminotransferase and alanine aminotransferase) levels. Three patients with murine typhus experienced thrombocytopenia. Interestingly, we found that two murine typhus patients, aged 68 and 69 years, developed respiratory distress syndrome while hospitalized. Both patients had pleural effusion and lung infiltration (Table 2 and Fig. 3).

Chest radiographs of murine typhus patients with respiratory distress syndrome identified in this study, infected with isolates AB3775
Demographic and Clinical Characteristics of Rickettsial Infections Identified in This Study
Abnormal laboratory results.
ALT, alanine aminotransferase; AST, aspartate aminotransferase; F, female; M, male; NI, no information; NT, not tested; PLT, platelet; T, temperature; WBC, white blood cell.
A single case of cat-flea rickettsiosis, caused by infection with R. felis, was diagnosed in February 2021. The 33-year-old male patient (AB3572) presented with mild symptoms of headache (body temperature 36.9°C), myalgia, and arthralgia. He was not admitted to the hospital, and only a finding of chikungunya in the context of fever with arthralgia was documented in laboratory tests.
Two scrub typhus cases, caused by infection with O. tsutsugamushi (AB3696 and AB4595), were identified in this study. The first patient was a 31-year-old male with a history of travel to Northern Thailand 2 weeks before presenting with fever (body temperature 39.1°C), sore throat, headache, and myalgia. The second patient was a 29-year-old male living in Samut Sakhon Province, who presented with fever (body temperature 38.2°C), headache, myalgia, nausea, and vomiting. Notably, neither patient exhibited an eschar.
Discussion
By leveraging leftover nucleic acid samples extracted from acute-phase serum samples of patients primarily suspected of having arbovirus infection, this study successfully detected Rickettsia. Of 609 samples analyzed, real-time PCR identified 9 cases positive for Rickettsia spp. and 2 cases positive for O. tsutsugamushi. Subsequent phylogenetic analyses based on partial 17-kDa common antigen and gltA gene sequences revealed that eight of the Rickettsia isolates belonged to the R. typhi species, whereas the remaining isolate was identified as R. felis. Notably, the validated sequences of the 56-kDa TSA gene of the O. tsutsugamushi isolates exhibited a close relationship to the Gilliam genotype.
The presence of animal reservoirs, their associated ectoparasites, and the microbial pathogens they harbor play a crucial role in facilitating zoonotic transmission to humans. This is particularly evident in the increased incidence of flea-borne rickettsioses within tropical urban settings, in which rodent infestations only exacerbate the problem (Blasdell et al., 2022). Notably, the oriental rat (Rattus rattus), trapped in the Salaya suburb of Bangkok, Thailand, has been identified as a reservoir host for both R. typhi, the causative agent of murine typhus, and R. felis, the causative agent of cat-flea rickettsiosis (Prompiram et al., 2020). Consistent with these observations, a prevalence study conducted in Bangkok revealed that a number of trapped small mammals, including members of the Rattus complex, were infected with R. typhi or R. felis (Rungrojn et al., 2021). Moreover, domesticated cats and dogs have also been identified as susceptible to R. typhi and R. felis infection, and these animals can, therefore, potentially serve as additional sources of bacterial transmission to humans (Mongkol et al., 2018; Phoosangwalthong et al., 2018).
Unlike flea-borne rickettsioses, scrub typhus, caused by O. tsutsugamushi, exhibits a higher prevalence in rural areas, particularly in the northern region of Thailand (Wangrangsimakul et al., 2020). This geographic disparity suggests that scrub typhus cases in urban settings are primarily associated with travel to endemic areas in which disease vectors and the pathogen circulate. Notably, one of the scrub typhus patients in the present study had a history of travel to Chiang Mai and Nan Provinces in the northern region of Thailand. Interestingly, the 56-kDa TSA gene nucleotide sequence from this patient’s isolate exhibited similarity to an O. tsutsugamushi isolate found in Mae Hong Son Province, further supporting the link between travel and infection (Fig. 3).
Remarkably, we observed two cases of severe murine typhus with pulmonary system involvement in elderly patients. Previous studies indicated that delayed diagnosis, particularly in elderly individuals, and inadequate response to antibiotic treatment are significant risk factors for severe disease (Civen and Ngo, 2008; Sakamoto et al., 2013). Although murine typhus typically presents with mild clinical symptoms, it is essential to recognize that severe disease with pulmonary manifestations can occur, as reported in this study. Therefore, we recommend prioritizing the investigation for common vector-borne diseases like murine typhus, particularly in patients presenting with pulmonary involvement. This recommendation, particularly relevant in regions with high murine typhus prevalence, such as Bangkok (Luvira et al., 2019), emphasizes the importance of awareness and timely consideration of murine typhus in relevant clinical contexts to ensure accurate diagnosis and appropriate management.
Owing to their intracellular nature, Rickettsia are carried by immune cells like monocytes and macrophages through the bloodstream and preferentially infect vascular endothelial cells (Sahni et al., 2019). This limited presence in the blood makes diagnosing Rickettsia infections a challenge. Buffy coat samples are often a better option for detecting Rickettsia. However, investigating serum by PCR-based assay offers a promising alternative (Nascimento do et al., 2009). In this study, utilizing remaining nucleic acid samples extracted from acute-phase serum also showed successful detection of Rickettsia. A limitation of our study is the lack of serological testing. This reliance solely on PCR-based detection could lead to false-negative results owing to the typically low number of bacteria and limited analytical sensitivity, potentially underestimating the true incidence. Nevertheless, this study, despite its limitations, demonstrated the feasibility of detecting Rickettsia infections and emphasized the importance of prompt diagnosis to prevent disease progression and complications such as pulmonary involvement.
Footnotes
Acknowledgments
This study would not have been possible without the active support of all the staff at the Center of Excellence in Clinical Virology, Faculty of Medicine, Chulalongkorn University.
Data Availability Statement
The authors confirm that the data supporting the findings of this study are available within the article and its supplementary materials.
Author Disclosure Statement
The authors declare that they have no conflict of interest.
Funding Information
This work was supported by
Supplementary Material
Supplementary Figure S1
Supplementary Tables S1
Supplementary Tables S2
Supplementary Tables S3
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.