
Abstract
Background:
Toxoplasmosis is caused by infection with Toxoplasma gondii. No Symptoms in healthy people. Notably, very dangerous symptoms in immunocompromised, or patients with immune diseases. Previous research has shown that the parasite’s resistance to drugs continues to emerge and has indicated this resistance as a cause for concern. In this context, researchers have a great responsibility to search for alternative treatments, as well as to develop existing ones. Essentially, this improves the therapeutic efficacy of drugs and prevents the emergence of resistance to them. The present study aims to evaluate antitoxoplasma effects of niosomal loaded curcumin and silymarin and their synergistic effects with clindamycin against T. gondii RH strain in vitro.
Materials and Methods
: Experiments were conducted on the tachyzoites of T. gondii RH-strain, based on: the free and nieosomal compounds of curcumin and silymarin, in addition to the drug clindamycin. Data were collected to estimate parasite viability during exposure to the therapeutic compounds under study using a special MTT assay ((3-(4, 5-dimethylthiazolyl-2)-2,5-diphenyltetrazolioum bromide) assay: is a colorimetric assay for measuring cellular growth) kit provided by (Bio Idea Company, Tehran, Iran). Hence, the effect of the therapeutic compounds on the parasite load was studied using the quantitative molecular technique real-time PCR.
Results:
The results indicate that the combination of N-silymarin and N-curcumin with clindamycin has active synergistic effects against T. gondii leading to complete elimination of the parasite. Data revealed that curcumin and silymarin in both their free and nisomal forms had inhibitory effects on the parasite, and minimal toxic effects on normal cells.
Conclusions
: The results highlight the successful synergistic effect of clindamycin and the niosomal compounds curcumin and silymarin in completely eradicating the T. gondii RH-strain. This finding contributes positively to the field of safe and effective treatments.
Introduction
Toxoplasma gondii (T. gondii) is an intracellular parasite infecting warm-blooded animals, which includes 30% of the human population, with higher rates in African, South American and European countries (Mose et al., 2020). T. gondii can escape mechanisms of host responses and blood–brain barrier via various mechanisms (Elsheikha et al., 2020).
In most cases, symptoms are not present and sometimes include flu-like and lymphadenopathy. More severe symptoms occur among immunocompromised individuals and in pregnant women causing fetal infection (Deganich et al., 2022; Furtado et al., 2011). In addition, the dormant cells persist within cysts and cerebellar and systemic disease and neuropsychiatric disorders may occur following recrudescence (Tyebji et al., 2019). This mechanism is the transformation of bradyzoites (latent form) into tachyzoites (causes of acute form), rapidly growing forms (Elsheikha et al., 2020; Lyons et al., 2002). Additionally, sporozoites exist in oocysts as the final stage of sexual reproduction, which occurs in the feline intestine (Cerutti et al., 2020).
Considering these, the treatment of cystic stage is difficult, precluding the effective elimination of parasites. The acute and reactivated infections by tachyzoites should be managed (Attias et al., 2020). The combination of pyrimethamine, clindamycin and sulfadiazine, or folinic acid targeting enzymes dihydrofolate reductase and dihydropteroate synthase has been effective against the infection (Dannemann et al., 1992; Katlama et al., 1996).
However, these drugs cause toxicity and allergic (hematological and dermatologic) reactions, need high costs and are unable to inhibit bradyzoites, thus alternative treatments such as atovaquone and azithromycin have been also used (Hodgson et al., 2018). Therefore, discovery of new formulations such as novel molecules is required. Herbal compounds have been investigated against various infectious diseases (Ghasemian et al., 2019; Kanaan et al., 2021).
The antiparasitic effects of curcumin have been demonstrated mainly through antioxidative and anti-inflammatory effects (Esmaeili Rad et al., 2022; Moradi et al., 2023). On the other hand, efficient drug delivery is pivotal to maintaining medicines and preventing unwanted side effects. Hence, niosomes have been outlined promising nanocarriers (Witika et al., 2022; Zarepour et al., 2023). The aim of this study was in vitro assessment of antitoxoplasma effects of niosomal loaded curcumin and silymarin and their synergistic effects with clindamycin.
Material and Methods
Niosome preparation and compounds/drug entrapment
To prepare nanoniosomes, surfactants (SpanTM 60 and Tween® 60) (Surya Dermato, Corp) and cholesterol (Sigma) were solubilized into chloroform and methanol (2:1 ratio) and were entirely mixed. Afterward, the solution was added to a specialized rotary balloon at 164 g and under vacuum conditions of 60°C to solubilize the mixture completely.
Next, 10 mL of each compound was added to the lipid film and rotated for 30 min in a distiller device at vacuum conditions for the hydration, and the particle size was decreased by the sonication. The entrapment efficacy (EE%) which is calculated by the ratio of compounds loaded within the niosome to the free forms was also measured. For the separation of free compounds from the entrapped forms, the centrifugation at 14,000 g for 45 min was performed resulting in the precipitation of entrapped form. The absorbance was read at 540 nm and the following formula was used:
Various formulations of niosomes were prepared according to SpanTM 60 and Tween® 60 molar ratio and the encapsulation rate and niosome sizes were determined. The physical and chemical features, sizes, and particle distribution were determined using a zetasizer device. In addition, the niosomes morphology was determined using a scanning electron microscope (SEM). The releasing rate of compounds was assessed using a dialysis bag test.
Cytotoxicity of curcumin and silymarin on vero and HeLa cells
Vero and HeLa cells were purchased from Pasteur Institute of Iran (s previously purchased from the American Type Culture Collection (ATCC) and cultured in Dulbecco’s modified Eagle’s medium (DMEM) (Gibco, USA) and supplemented with 100 mg/mL streptomycin, 100 units/mL penicillin (Invitrogen, Carlsbad, CA, USA), and 10% fetal bovine serum (FBS) (Gibco, USA). Curcumin (MERCK-Schuchardt) and silymarin (Sigma–Aldrich) various concentrations (1–512 µg/mL) were prepared and 100 µL of each was added to wells containing 100 µL of Vero or HeLa cells and incubated at 37°C overnight (with sufficient humidity and 5% CO2). Afterward, the plate was washed using PBS and MTT reagent (100 µL) was added and incubated at 37°C for 2–4 hr. A purple color indicated living cells, and the reaction was ceased using 10% SDS stopper reagent (100 µL).
In vitro cell viability assay
After the preparation of fresh culture of T. gondii RH strain, curcumin and silymarin loaded in niosomes were diluted and tested against the parasite. Clindamycin (Sigma–Aldrich, St. Louis, MI, USA) was used as the antiparasitic drug. Vero and HeLa cell lines were purchased and used as the host cells in suitable conditions (humidity and 5% CO2 incubator at 37°C). The median effective concentration (EC50) of drug/compounds was defined by inhibition of 50% of cells or parasites. In addition, selectivity index (SI) was defined by division of percentage of cells EC50 by parasite EC50. Higher SI index exhibits better activity against parasites with low cytotoxicity on normal cells (Salin et al., 2020), then a special MTT assay kit provided by (Bio Idea Company, Tehran, Iran) was used to determine the vitality of the parasites according to the method used by (Ahmadi et al., 2023).
T. gondii RH strain maintenance
The tachyzoites of T. gondii RH strain were obtained from National institute of genetics of Iran and were passed by the intraperitoneal injection into the healthy BALB/c mice every 2–3 days. For counting of parasites, they were collected from fluid and cultured in tubes. After centrifugation at 200 g for 10 min, the supernatant was collected, which had tachyzoites and was centrifuged again at 1000 g for 10 min to obtain tachyzoites pellet and next cultured in Dulbecco’s Modified Eagle Medium (DMEM) containing 10ug/mL gentamicin, 2 mM glutamine and 10% heat inactivated (56°C for 45 min) FBS 0.5% penicillin-streptomycin (PS), and counted using inverted light microscope (trypan blue dye) (Khan & Grigg, 2017; Salin et al., 2020).
Treatment of infected cells
After 80%–90% confluence of Vero cells, a number of 1*106 cells/mL was added to each well of a 96-well plate in triplicate and a number of 1*104/mL tachyzoites (100 µL) were added to them. 100 µL of it was added to another well as negative control and also 100 µL was transferred to another well as positive control with clindamycin. Then, 100 µL of niosomal loaded of each of curcumin and silymarin (concentration ranges of 1–512 µg/mL) and their combinations with clindamycin were added to each well of the test as separate groups and media were incubated for 24 hr. It is worth noting about the concentration of the compounds that the first well had double its concentration (initial volume), equal to 1024 µg/mL, reaching 512 µg/mL in the liquid volume of the well (final volume).
Cell viability was calculated based on the percentage of viability of tested cells per control untreated cells (Bajelan et al., 2020; Salin et al., 2020). T. gondii and sterile water were used as positive and negative control, respectively.
Apoptosis assay
The apoptogenic potential of compounds on HeLa cancer cells was determined by flow cytometry. HeLa cells were seeded in a 6-well plate (2.5 × 105/well), and curcumin and silymarin compounds (100 µg/mL) were added. After 48 hr, the cells were harvested and washed with PBS and subjected to Annexin-V-FLUOS staining kit (Roche Diagnostics, USA) as the protocol. The rate of apoptotic cells was measured utilizing an Attune Acoustic Focusing Cytometer.
Quantification of tachyzoites
The quantitative molecular technique Real-Time PCR was relied upon to investigate the number of tachyzoites of T. gondii RH-strain in each group of samples, during days 0, 3, 6, and 10 of the experiment (Cassaing et al., 2006; Meneceur et al., 2008). The first step; Parasite DNA was extracted from tachyzoites using the Universal Genomic DNA Extraction Kit prepared from (Takara et al., 2006) according to the manufacturer’s protocol attachment.
Real-time PCR fluorescence was detected using the iQTM5 system (Bio-Rad), according to primers based on the specific SAG1 gene (128 bp fragment) according to (Wang et al., 2014) as follows: F: 5’ CTGATGTCGTTCTTGCGATGTGGC3’ and R: 5’GTGAAGTGGTTCTCCGTCGGTGT3’’.
When the amplification process begins, with each amplification including a negative control (distilled water) and a positive control (purified genomic DNA). For the assay and protocol, the contents of each reaction mixture included 9.7 L of sterile distilled water, 2 μL of template DNA, and 0. 4 µL of each primer (20 pM), and 12.5 µL of THUNDERBIRD SYBR® Real-Time PCR Mix (TOYOBO). It should be noted that DNA diluted 10-fold to standard curve variation (in triplicate) was prepared under these conditions: hot started at 95°C (1 min), followed by (denaturation) 40 cycles of 95°C (5 s), then annealing 60°C (15 s), followed by amplification to 72°C (10 s). Cycle threshold (CT) values were calculated, and values <35 were considered positive results. The CT values were obtained with the standard values as a control to determine the expression rate and then the CT values were converted to the actual numbers of parasites present based on the calibration curve and according to the method used before (Yu et al., 2013; Charisse et al., 2021).
Statistical analysis
The results were analysis using chi-squared (χ2) and t-test and analysis of variance (ANOVA) tests at p value <0.05, which was defined as a significant result.
Results
Synthesis and characterization of niosomes
The present results show that the optimal formulation of nanoparticles to obtain effective attributes was considered based on the molar ratio of SpanTM 60, Tween® 60 (50:50), cholesterol:surfactant (1:1) and 1 mg of each compound followed by sonication for 7 min. Furthermore, the highest percentage of encapsulation efficiency (EE%) is contained in the size of 203.2 nm niosomes which is 76.2%, and this is the indicated amount of the active ingredient in curcumin and silymarin that successfully resides inside the niosomes. The release rate for N-curcumin (N-cur) is 33% and N-silymarin (N-sil) 29%, which is less than the release pattern of their free forms, which reached 92% for curcumin and 94% for silymarin, as in (Fig. 1). Statistical analysis of these results showed a significant difference (p < 0.0001).
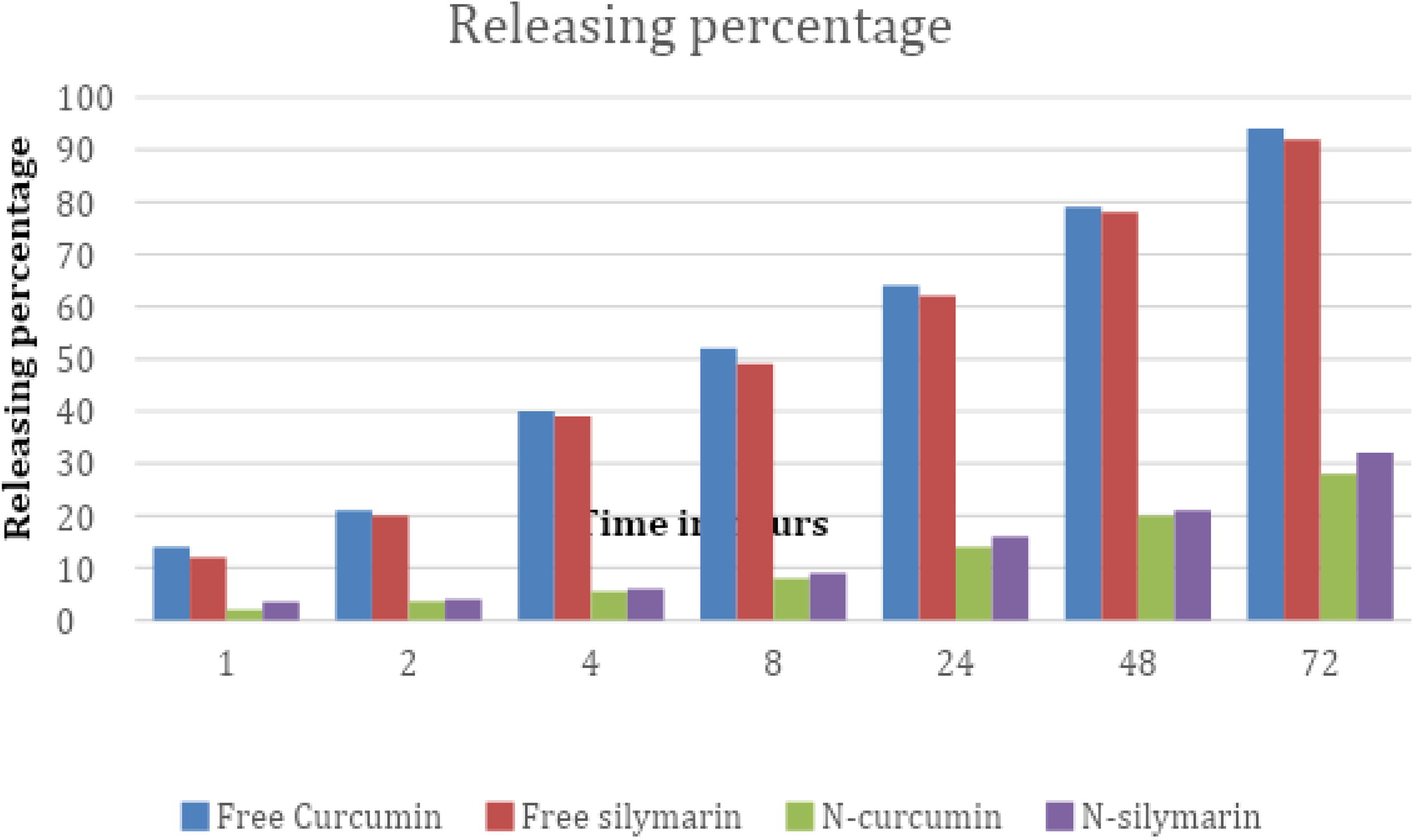
The releasing rate of compounds N-cur and N-sil compared with their free forms.
The results showed that the cell EC50 of clindamycin, curcumin, and silymarin on Vero cells was 46.56 μg/mL, 40.11 μg/mL, and 43.7 μg/mL, respectively, while in HeLa cells included 48.29 μg/mL. ml, 44.2 μg/mL, and 41.9 μg/mL. Data indicate relatively low cytotoxicity to normal cells. Very slight differences in cytotoxicity were observed between the free and niosomal forms of curcumin and silymarin, but they were negligible. Statistical analysis revealed it, and expressed no significant differences (P < 0.064 and P < 0.077, respectively). As Fig.: 2 and 3

Cytotoxicity of both (free and somatic forms of curcumin and silymarin) and clindamycin, against Vero cells after 24 h of exposure.

Cytotoxicity of both (free and somatic forms of curcumin and silymarin) and clindamycin, against HeLa cells after 24 h of exposure.
Moreover, the results showed that the EC50 concentration of clindamycin, curcumin, and silymarin against T. gondii was 4.27 μg/mL, 4.1 μg/mL, and 4.9 μg/mL, respectively. This data indicate that the required concentration of clindamycin and its compounds shows a half-maximal effect on T. gondii, which is a much lower concentration compared to its effect on HeLa and Vero cells, and as a result, the SI calculated, was 11 (46.56/4.27 and 48.29 μg/mL), which means that clindamycin and other compounds under study are more effective on the parasite than on normal cells.
At the beginning of the experiment, cell viability was 100% in all groups, at a concentration of 1 μg/mL of the experimental therapeutic compounds, and with increasing compound concentration multiples, parasite cell viability decreased significantly. The present results showed a rapid decrease in cell viability to 0% at a concentration of 16 μg/mL for the N-curcumin+Clindamycin (N-cur+clin), N-silymarin+Clindamycin (N-sil+cur), and Clindamycin groups. This indicates their superiority in completely eliminating the parasite over the rest of the compounds. Moreover, there was a more effective effect when exposed to the NCur+Clin, compound, which is almost negligible, as there were no statistically significant differences when analyzing this result.
The results also showed that (N-cur) and (N-sil) were superior to the free forms of curcumin and silymarin in their toxic effect, as the cell viability reached 18% with free curcumin and 24% with free silymarin, while the cell viability reached 14% with (N-cur) and 16% with (N-sil) at a concentration of 512 μg/mL, which confirms that both the free and combined forms had an inhibitory effect on the parasite, but did not reach the stage of complete eradication as is the case in its synergistic association with clindamycin. as (Fig.: 4). Statistical analysis of the data indicate the presence of significant statistical differences between the composition and concentration of the therapeutic compounds on the one hand, and the vitality of the parasite on the other hand (p < 0.05).
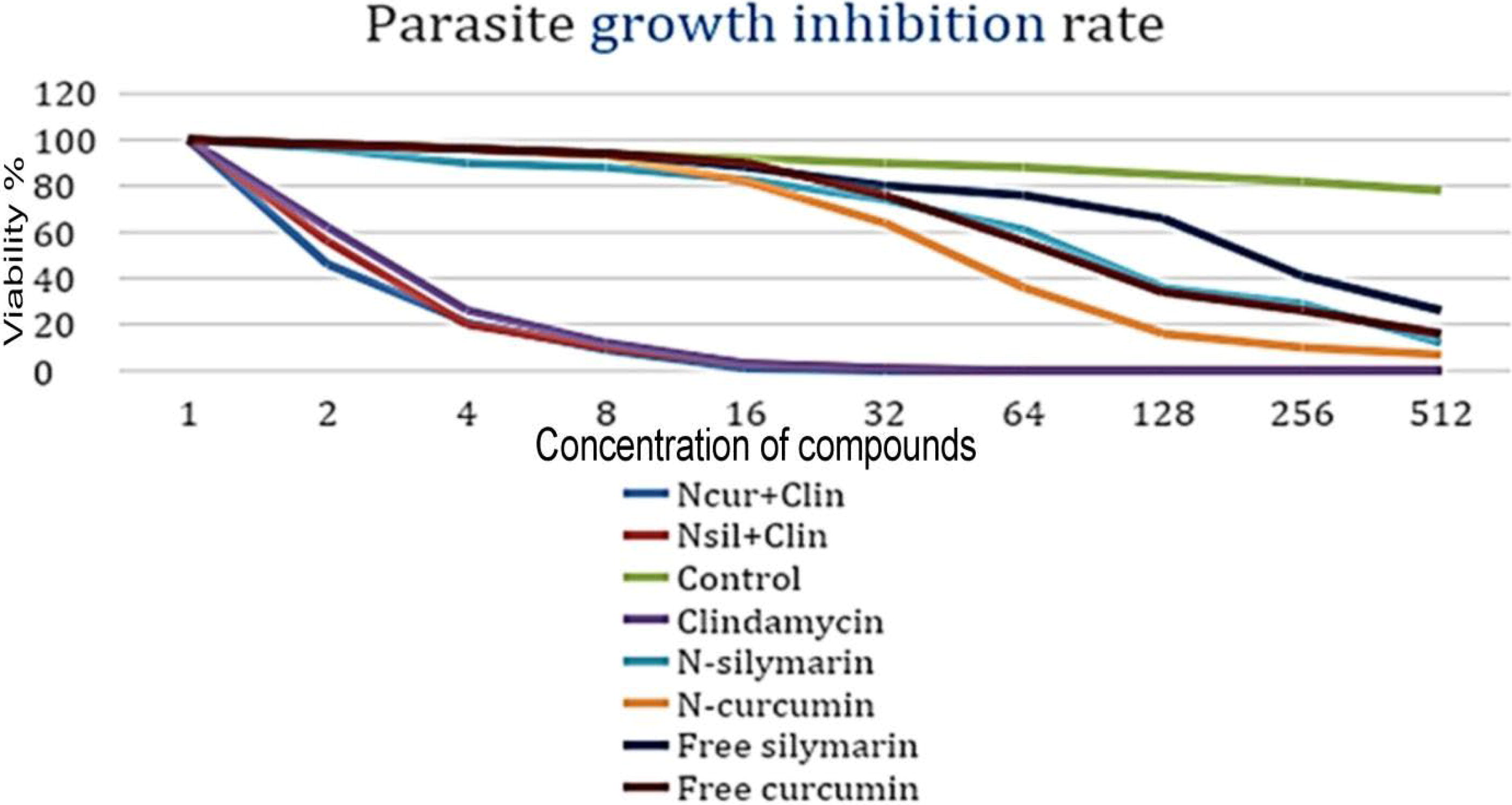
Cytotoxicity effects of clindamycin, free and niosomal forms compounds against T. gondii cells at various time intervals.
Apoptosis of HeLa cells treated with curcumin and silymarin
The results found that the apoptosis induction or survival rate of HeLa cells in exposure to 100 µg/mL of curcumin and silymarin: 4% ± 12 for curcumin and 6% ± 31 for silymarin, which were nonsignificantly different, compared with control (water being 3.5%±29). Therefore, bioactive compounds curcumin and silymarin had no potent inductive cell death effects on HeLa cells.
Parasitoid quantification using molecular methods
The results of performing tachyzoite quantification according to CT values using real-time PCR indicated: that the number of tachyzoites upon exposure to clindamycin, NCur+NSil and NSil+Clin were almost similar, with no significant differences. All of them indicate the effectiveness of the compounds under trial in reducing the parasite load. The combinations N-cur+Cli, N-sil+Clin and Clindamycin showed superiority in completely eliminating the parasite. In particular, this confirms the existence of a synergistic relationship, significantly enhancing the lethal effects compared with free forms and free nesosomes. The data confirm the statistically significant differences between the combination of silymarin and curcumin with clindamycin for synergistic antiparasitic effects (p < 0.0001), resulted in a significant reduction in T. gondii load compared to the individual forms (p < 0.001). as (Fig. .5):

The parasite load in various treatment groups; CT (relative expression), in experiment’s days.
Discussion
Challenges in the toxoplasmosis treatment include resistance, side effects, limited choices, timing of treatment, complications, high treatment costs and coinfections (Alday & Doggett, 2017; Konstantinovic et al., 2019). In this study, after niosome preparation, the entrapment of silymarin and curcumin singly and in combined forms with clindamycin were implemented and their antitoxoplasma effects were evaluated.
Nanovesicular carriers such as niosomes have the ability to carry lipophilic and hydrophilic compounds or drugs, supporting those drugs that are rapidly degradable. Furthermore, increased solubility, systemic circulation, and reduced toxicity are other aspects of these types of nanocarriers. In this context, niosomes contribute to targeted and controlled drug delivery, improving their bioavailability (Esmaeili Rad et al., 2022; Zarepour et al., 2023). Niosomal drug delivery has been applied to many drugs in the treatment of different diseases such as: cancer, inflammatory diseases, infectious diseases, and adjuvant drug delivery (Moammeri et al., 2023; Witika et al., 2022).
The current results show that the release rate of free therapeutic compounds was much higher than that of their compound forms. This is due to the fact that drugs bound to nanoparticles are automatically designed to release the drug in a slow and continuous mechanism, which reduces the side effects of the drug and increases its efficiency (Xu et al., 2020; Zeb et al., 2022). This slow release is a controlled release to improve the targeting of the drug to the affected tissues, and it may play a role in improving drug absorption and maintaining the treatment effect for a long time (Weng et al., 2020; Zeb et al., 2022).
The present results reveal that the combination of N-silymarin and N-curcumin with clindamycin had synergistic antiparasitic effects. This may be due to the fact that some plant extracts enhance the efficacy of drugs on the one hand, and reduce the drug resistance that the parasite may carry on the other hand (Elazab and Arafa, 2022). This is consistent with (Cheraghipour et al., 2021) who confirmed that the efficacy of atovaquin against T. gondii is significantly increased when combined with Myristica fragrans extract, proving the existence of a synergistic relationship that exceeds the ability of the drug alone.
The niosomal forms of curcumin and silymarin were observed to be superior to their free forms in inhibiting the parasite’s vitality. This may be due to the specific targeting property of the nanoparticles, which ensures that the compound reaches the target cell (Gavas et al., 2021; Tripathi et al., 2023).
It was clear that curcumin and silymarin in their free and niosomal forms have the ability to inhibit the parasite over a certain period of time. Complete elimination of the parasite was only achieved when combined with clindamycin and its agreement with (Rageh et al., 2022) in their laboratory experiments on mice on the possibility of using N-cur as an adjuvant to increase the efficiency of spiramycin treatment in the treatment of acute toxoplasmosis, as an enhanced compound for biological therapy. However, this result does not agree with (Azami et al., 2018), the single N-Cur is superior to atovaquone in treatment of toxoplasmosis. This may be attributed to the design of the experiment, the different parasite strains studied, the nature of the host, and the surrounding conditions of the experiment.
The therapeutic compounds under study achieved high efficiency in rapidly reducing the numbers of T.gondii, from the first days of the experiment, and this continued gradually until the parasite disappeared completely on the 10th day. This refers to their efficiency in reducing the parasite load. This is consistent with the findings of (Madi et al., 2012) on the effectiveness of clindamycin alone at low concentrations, for 21 days in gradually reducing the parasitic load of T. gondii in AIDZ patient. This result is also supported by (Yapar et al., 2005) in their reference to the effectiveness of clindamycin alone in treating cerebral toxoplasmosis, and its effective effect in reducing the parasitic load, until it is completely eliminated.
The results concluded that the synergistic effects of N-cur+clin, N-sli+clin were effective in eliminating T. gondii. This indicates a positive interaction between them. It led to the emergence of properties that increase the antiparasitic efficacy of the drug, prevent the parasite from developing resistance to the drug, and reduce the potential side effects of the drug. The limitations of this study: lack of evaluation of clinical isolates of parasites from patients. Lack of in vivo evaluation and molecular testing of gene expression, to reveal the precise mechanisms of action, and knowledge of the pathways affected by these compounds. Furthermore. Specifically, screening of bioactive herbal compounds contributed to the discovery of new antiparasitic agents, nontoxic to the human body. Further studies are needed in this area.
Conclusions
In this research, the focus was on the synergistic effect between clindamycin and the free and N-curcumin forms of curcumin and silymarin in the treatment of toxoplasmosis. The results confirmed that the use of a combination of compounds N-cur+clin, N-sli+clin led to improved treatment efficacy, compared with using each compound alone for treatment. These results contribute to clarifying the efficacy of the compounds under study in the field of drugs, some limitations must be taken into consideration, such as the urgent need for more similar studies. Researchers should continue to investigate the synergism of compounds, drugs and plant extracts, in different frameworks, in order to develop practical applications for these important treatment strategies.
Footnotes
Acknowledgments
This study was carried out with the assistance of the laboratories of the biology Department, College of Science, Al-Qadisiyah University.
Author Disclosure Statement
No competing financial interests exist.
Funding Information
No funding was received for this article.