
Abstract
Background:
Dengue virus (DENV) infection, caused by serotypes DENV 1–4, represents a significant global public health challenge, with no antiviral drugs currently available for treatment. The host Protein kinase B (AKT) signaling pathway is crucial for DENV infection, presenting a potential target for antiviral drug development.
Objective:
This study aimed to evaluate the antiviral activity of kinase inhibitors that target the AKT pathway, focusing on the compound AT13148. Methods: A mini-screening was conducted to identify kinase inhibitors with antiviral properties against DENV-2. The effects of AT13148 on viral RNA replication and translation were assessed in a dose- and time-dependent manner following DENV-2 entry. The mechanism of action was further investigated by evaluating the impact of AT13148 on AKT kinase activity and phosphorylation status.
Results:
AT13148 exhibited potent antiviral activity against DENV-2, significantly inhibiting viral RNA replication and translation post-entry. The compound was found to inhibit AKT kinase activity through hyperphosphorylation.
Conclusion:
The findings indicate that AT13148 effectively targets the AKT pathway, demonstrating potential as an antiviral therapeutic against DENV-2 by interfering with the virus's post-entry processes. Further in vivo studies are warranted to assess the efficacy of AT13148 in controlling DENV infection.
Introduction
The genus Flavivirus consists of arthropod-borne viruses, including important human pathogens such as Zika virus (ZIKV), dengue virus (DENV 1–4), West Nile virus (WNV), yellow fever virus (YFV), Japanese encephalitis virus (JEV), and tick-borne encephalitis virus featured as (+) ssRNA viruses with similar life cycles (Zhao et al., 2021). DENV infection (serotypes 1–4) through mosquito bites can cause disease symptoms such as dengue fever and severe dengue fever. Dengue infection affects more than 50 million people annually in tropical and subtropical regions and has become a major global public risk (van Leur et al., 2021). The novel QDENGA® (dengue live, attenuated tetravalent vaccine) has been recently approved in Indonesia for use regardless of previous exposure to dengue, providing a promising method to prevent infection. Nevertheless, no antiviral drugs are approved for treating dengue fever. Identifying small molecules against DENV infection is as important as developing novel DENV vaccines (Patel et al., 2022; Samrat et al., 2022).
The viral and host proteins involved in virus infection are major targets for developing novel antiviral drugs (Obi et al., 2021). Viral proteins like E protein, nonstructural protein 3 (NS3), and NS5 are the most common targets trialed in the search for anti-DENV inhibitors. Several compounds are in clinical trials to evaluate the anti-DENV activity (Samrat et al., 2022). However, these compounds targeting viral proteins are more likely to encounter drug resistance due to the rapid mutation of DENV genomic RNA and to fail in clinical trials. The alternative approach to developing anti-DENV drugs targets the host proteins essential for the virus life cycle (Schneider and Hoffmann, 2022). This strategy may help researchers avoid the drug-resistance trap. Generally, the Flaviviruses exhibit the same life cycle progression and require similar host factors; the host targeting strategy is promising to develop a broad spectrum of anti-Flavivirus compounds (Sole Burali et al., 2022). The drugs usually target host pathways or enzymes that are involved in the replication cycles of different viruses.
AKT is a serine/threonine kinase that is an important downstream effector of PI3K. The AKT/PI3K pathway plays an important role in the pathogenesis of viral infections exploited by viruses to facilitate their replication (Albentosa-Gonzalez et al., 2021; Fattahi et al., 2022; Kirsch et al., 2020; Lahon et al., 2021). During the Flavivirus infection, the AKT pathway is activated to maintain the host cell viability and counteract the virus-induced cellular apoptosis. Flavivirus capsid proteins can activate the AKT pathway and enhance downstream signaling pathways that affect cell survival and metabolism, providing a favorable environment for virus replication. The PI3K/AKT pathway may serve as a promising target for developing anti-Flavivirus therapeutics (Okamoto et al., 2017) (Airo et al., 2018).
Herein, a primary screen was conducted on a mini library from Selleck Chemicals LLC containing 17 kinase inhibitors targeting AKT and identified five kinase inhibitors with potential anti-DENV-2 activity. AT13148 was chosen for further study due to its greater efficacy against DENV-2 infection in Huh-7 and A549 cells and its novelty as an antiviral compound. AT13148 is an oral multi-AGC kinase inhibitor primarily designed with potent pharmacodynamic and antitumor activity, which has finished phase I clinical trials. We performed various experiments to understand the possible mechanism of the anti-DENV-2 activity of AT13148.
Methods and Materials
Cell lines, viruses, and chemicals
The Huh-7 cell line was obtained from the JCRB cell bank of Okayama University. The A549, 293T, HeLa, and BHK-21 cell lines were obtained from the American Type Culture Collection. Cells were cultured in DMEM (ThermoFisher) supplied with 10% fetal bovine serum. HUVECs were purchased from ThermoFisher (C0035C; Gibco) and cultured in Medium 200 (Gibco) with a 2% low serum growth supplement (Gibco). DENV-2 (strain 16681) was amplified in C6/36 cells (Pang et al., 2001). For DENV-2 infection, the cells were incubated with the virus (MOI = 1) for 2 h at 37°C with gentle rocking every 15 min. After 2 h, cells were rinsed, overlaid with a complete or designed medium, and incubated at indicated time points. The Kinase Inhibitor Library (Catalog No. L1200) was purchased from SelleckChem LLC. The heparin sodium was purchased from Sigma-Aldrich Inc.
Relative quantitative real-time PCR analysis
To determine the viral RNA level in cells, total RNA was extracted from the treated cells with the E.Z.N.A.® Total RNA Kit (Omega Bio-tek). RNA was converted into cDNA using PrimeScript RT Master Mix (TaKaRa). The reaction was performed with DENV-2 NS5 primers (5′-GGTTTTGGGAGCTGGTTGAC-3′ and 5′-ACTCTAAGAAGCGTGCTCCA-3′) and β-actin primers (5′-GGTTTTGGGAGCTGGTTGAC-3′ and 5′-ACTCTAAGAAGCGTGCTCCA-3′). Each reaction was performed in duplicate, and the comparative cycle threshold (Ct) method was used for the relative quantification of normalized gene expression relative to β-actin mRNA.
Western blotting
Cells were washed and treated with lysis buffer (1% Triton X-100, 20 mM Tris-HCl pH 7.5, 150 mM NaCl, 1 mM EGTA, 1 mM Na2EDTA, 2.5 mM sodium pyrophosphate, 1 mM Na3VO4, 1 mM beta-glycerophosphate, and 1 mg/mL leupeptin) on ice for 20 min, and the supernatants were collected to detect protein concentration by centrifuging at 12,000 × g for 20 min at 4°C. The samples (30 µg) were separated using 10% or 12% sodium dodecyl sulphate-polyacrylamide gel electrophoresis (SDS-PAGE) gel and transferred onto polyvinylidene fluoride (PVDF) membranes. The membranes were blocked for 1 h with 3% bovine serum albumin (BSA) at room temperature and incubated with the desired primary antibodies overnight at 4°C. Consequently, the membranes were washed three times with buffer and incubated with the secondary antibody, IRDye 680RD goat antirabbit secondary antibody or IRDye 800RD goat antimouse secondary antibody (LI-COR Biosciences), for another 2 h at room temperature. Protein detection and analysis were made using the infrared imaging system Odyssey (LI-COR Biosciences). For primary antibodies, Flavivirus group antibody 4G2 (GTX57154), DENV NS3 mAb (GTX124252), and capsid mAb (GTX103343) were purchased from Genetex Inc. GRP78 rabbit pAb (102056-T40) and ACTIN mouse mAb (100166-MM10) were purchased from Sino Biological, Inc. AKT rabbit mAb (#88800) and phospho-AKT (Ser473) rabbit mAb (#31957) were purchased from Cell Signaling Technology, Inc.
Flow cytometry
To detect the effect of AT13148 on virus entry, Huh7 cells in 6-well plates were incubated with DENV-2 on ice for 2 h and then washed with cold phosphate buffer saline (PBS) three times. Afterward, cells were incubated with various agents at 37°C with 5% CO2 for 2 h to allow virus entry and then treated with 0.25% trypsin to remove viruses attached to the cell surface. Cells were then treated with BD Cytofix/Cytoperm™ solution and incubated with DENV envelope mouse mAb (GTX629117, Genetex Inc.) and the 488-conjugated mouse secondary antibody at 37°C for 1 h. After washing, the bound DENV-2 and internalized DENV-2 were detected and analyzed using a flow cytometer (BD FACSCalibur™).
Cell viability assay
Cell viability was measured using the cell counting kit-8 (CCK-8) kit (Bimake, CN). Briefly, cells were seeded in 96-well plates. After being treated with AT13148, 10 μL CCK-8 solution was added to each well and incubated at 37°C for 2 h. The optical density value of each well was obtained at a wavelength of 450 nm. The cell viability of each well was calculated.
Determining inhibitory concentration (IC50) of AT13148
To determine the IC50 of AT13148, Huh-7 cells seeded in a 24-well plate at a density of 1 × 105 cells/well were infected with DENV-2 at an multiplicity of infection (MOI) of 1 for 1 h before being treated with AT13148 at six different concentrations: 0, 0.04, 0.2, 1.0, 5.0, and 10.0 μM. After 24 hpi, the viral RNA level was determined using RT-qPCR to assess inhibition efficiency. The inhibition efficiency was fit into a nonlinear regression curve, and the IC50, the concentration that results in a 50% inhibitory effect, was calculated using GraphPad Prism by interpolation.
Viral plaque assays
The plaque assay was conducted to quantify the DENV-2 titers. Briefly, BHK-21 cells were seeded in 24-well plates and incubated overnight. The supernatants of virus-infected samples were added to BHK-21 cells and incubated for 2 h with occasional shaking to ensure maximum infection. The supernatants were removed 2 h later, and the cells were washed twice with PBS before being overlaid with DMEM medium containing 3% carboxymethylcellulose (CMC) and 2% FBS. The overlaid cells were incubated for 7 days. Afterward, the supernatants were removed and washed with PBS until the CMC was washed out. The cells were then fixed and stained with 4% paraformaldehyde (PFA) and 1% crystal violet for 15 min. The virus titers were calculated in plaque-forming units per mL.
Statistical analysis
Statistical analysis was performed using a two-tailed unpaired t-test in GraphPad Prism software (La Jolla, CA). The data are presented as the means ± standard deviations (SDs; n = 3 or as otherwise indicated). All experiments were repeated at least twice.
Results
Kinase inhibitor AT13148 targeting AKT exhibits high anti-DENV efficacy
Considering the important role of the AKT pathway in DENV-2 infection, 17 inhibitors in the Kinase Inhibitor Library (Catalog No. L1200, SelleckChem) targeting AKT were tested on A549 and Huh7 cells for the anti-DENV-2 activity. As expected, AT7867, A-674562, and AT13148 exhibited high antiviral efficacy tested on Huh7 cells (Fig. 1A), and four compounds named Triciribine, AT7867, Uprosertib, and AT13148 exhibited high antiviral efficacy tested on A549 cells (Fig. 1B). Compounds AT13148 and AT7867 could strongly inhibit the expression of DENV-2 RNA in both cell lines at 10 μM without affecting cell viability (Fig. 1C, D). Interestingly, some compounds, such as AZD5363 and LY3023414, could promote DENV-2 RNA amplification (Fig. 1A, B). AT13148 was selected to further study the anti-DENV-2 activity (Fig. 1E). To further confirm the result of the viral load reduction assay, we evaluated the antiviral activity of AT13148 against DENV-2 in the BHK-21, 293T, HeLa, and HUVEC cell lines. AT13148 significantly reduced the DENV-2 RNA load in these cell lines (Fig. 1F).

Kinase inhibitor AT13148 exhibits potent antiviral activity against DENV-2 without obvious cytotoxicity.
AT13148 suppress DENV-2 with dose- and time-dependent effect
To evaluate the antiviral effect of AT13148 under different conditions, the viral protein and vRNA levels were assayed at different time points after infection with series doses in Huh7. Both structural protein capsid and NS3 were reduced when treated with AT13148 (10 μM) at 24, 36, and 48 hpi (Fig. 2A). Viral RNA load was significantly reduced at 24, 36, and 48 hpi with 1, 5, and 10 μM AT13148, respectively (Fig. 2C). The IC50 value of AT13148 against DENV-2 was 573.4 nM (Fig. 2B), while cell viability was unaffected under 10 μM (Fig. 1C), indicating that the selective index of AT13148 on DENV-2 was higher than 17.4. Furthermore, infectious particles in the supernatant of DENV-2-infected Huh7 cells were reduced when treated with AT13148 (10 μM) at 48 hpi (Fig. 2B). These results suggest that AT13148 inhibits DENV-2 replication at different time points in a dose-dependent manner.
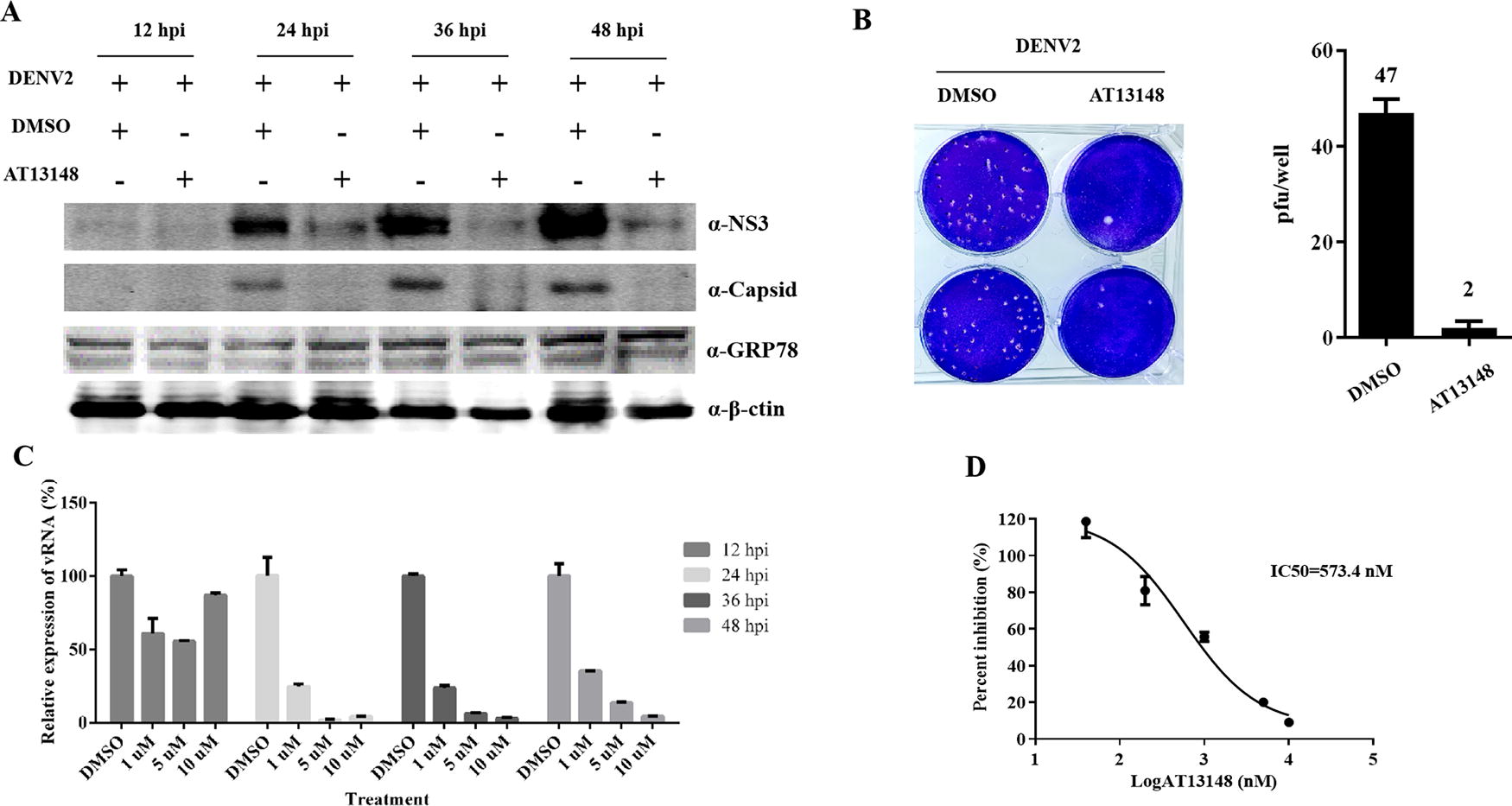
AT13148 significantly suppresses DENV-2 infection in vitro.
AT13148 suppress DENV-2 after entry
To elucidate the antiviral mode for AT13148, the effects of AT13148 on the DENV-2 life cycle were studied in Huh7 cells. 4G2 is a pan-Flavivirus envelope antibody that could neutralize DENV-2 infection. AT13148 had no suppressive effect on viral binding and endocytosis, as pretreatment with AT13148 did not inhibit DENV-2 infection, while 4G2 could significantly neutralize DENV-2 infection (Fig. 3A, B). AT13148 exhibited an inhibitory effect even at 12 hpi (Fig. 3C). Combined, these data illustrate that AT13148 potently inhibits DENV-2 replication postvirus entry.
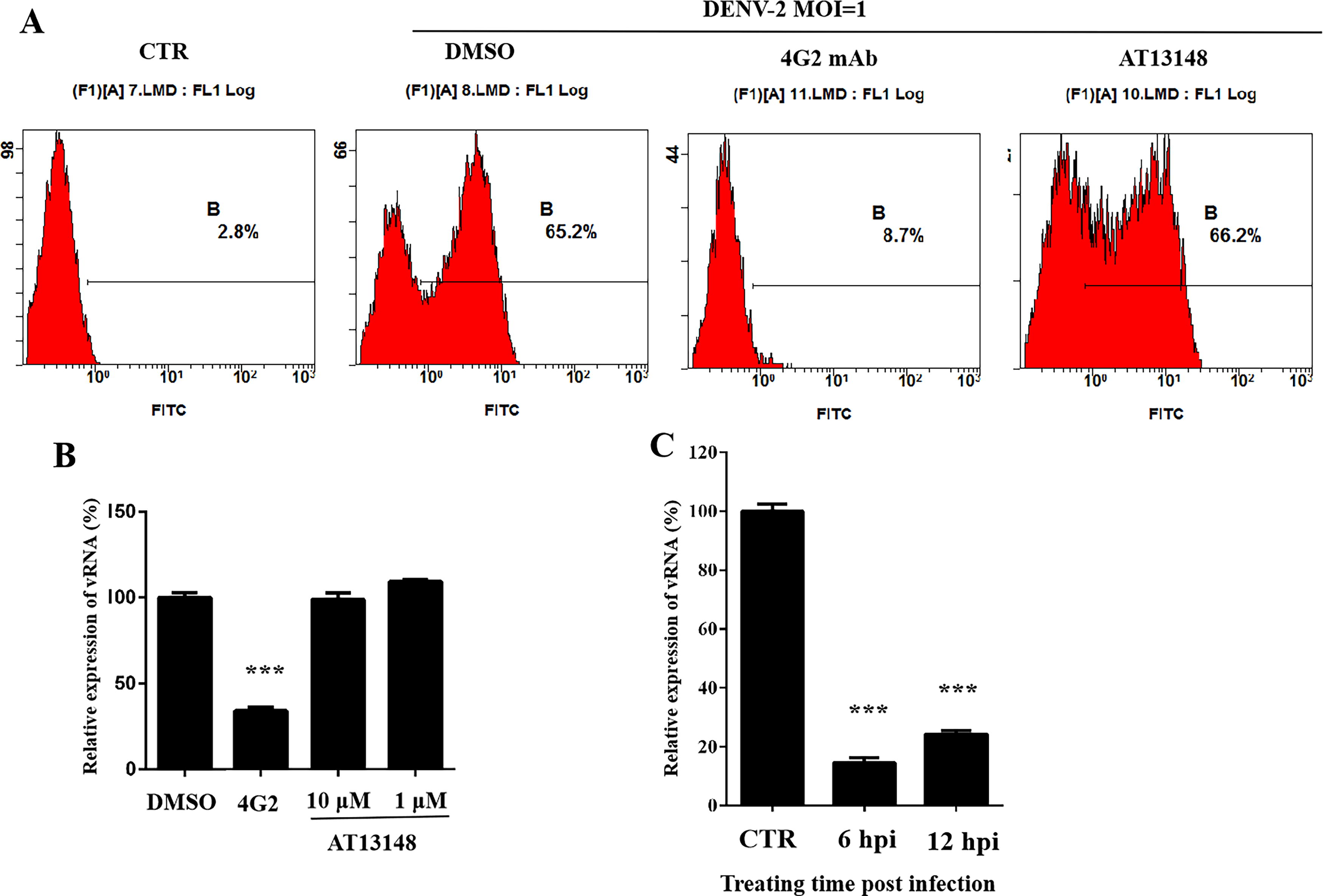
AT13148 inhibits DENV-2 replication postvirus entry. Huh7 cells were treated with 10 μM AT13148, 10 μM DMSO, and 20 μg/mL 4G2 and then infected with DENV-2;
Antiviral effect of AT13148 was mediated by downregulation of AKT activity
Previous studies show that the anti-DENV mechanism of AR-12 involves downregulating AKT and 78 kDa glucose-regulated protein (GRP-78) (Chen et al., 2017). However, we did not find that GRP78 was downregulated when treated with the AKT inhibitor AT13148 (Fig. 2A). Instead, the results show that AT13148 substantially hyperphosphorylated the AKT protein (Fig. 4), which is consistent with a previous study (Peeters et al., 2022). Interestingly, previous studies have indicated that vimentin is a novel AKT target mediating motility and invasion of cells (Zhu et al., 2011). To further verify the activity of AKT, we examined the protein level and posttranslational modification of vimentin. AT13148 reduced the phosphorylation level of vimentin in a dose-dependent manner while the total vimentin remained unchanged (Fig. 4). In this regard, these data suggested that AT13148 inhibits DENV-2 replication through inactivation of AKT kinase.

Western blot analysis of the AKT pathway involved in antiviral activity. Huh7 cells were treated for 24 h with DMSO (lane 1), AT13148 (lane 2), DENV-2 (lane 3), and DENV-2 with different concentrations of AT13148 (lanes 4–6); cell lysates were subjected to SDS-PAGE and immunoblot analysis with antibodies to DENV NS3, total AKT, phospho-AKT, total vimentin, phospho-vimentin, and β-actin.
Discussion
Host cell kinases are important host factors involved in virus life cycle and pathogenesis. The AKT pathway is reported to be activated by Flavivirus infection and to be involved in crucial processes such as cell survival to maintain host cell viability until the end of the viral cycle (Albentosa-Gonzalez et al., 2021; Okamoto et al., 2017). The AKT pathway is a promising target for developing antidengue therapeutics. Herein, a mini library containing 17 AKT-targeted kinase inhibitors was tested in A549 and Huh-7 cells for their anti-DENV-2 activity. As expected, several compounds exhibited potent anti-DENV-2 activity (Fig. 1). Although several compounds had little effect on DENV-2 replication, the inhibition activity of these compounds was associated with different downstream pathways of AKT.
Herein, AT13148 exhibited antiviral activity for the first time. AT13148 is a potent inhibitor of selected AGC kinases, including AKT, PKA, SGK, p70S6K, and ROCK, and was originally designed as a novel oral inhibitor with potent antitumor activity (Yap et al., 2012). AT13148 shows good pharmacodynamics and has finished phase I clinical trial (McLeod et al., 2020). The anti-DENV-2 effect of AT13148 was further investigated in the other five cell lines and strongly inhibited DENV-2 replication at noncytotoxic concentrations. Similarly, AR-12, a potent inhibitor of PDK-1 with anticancer activity, has shown a greater inhibition of PI3K/AKT signaling and suppressed DENV and ZIKV infection in vitro and in vivo (Chen et al., 2017). The AR-12 derivatives could also suppress DENV, ZIKV, and JEV replication at nanomolar concentrations, exhibiting 10-fold improvements in the efficacy and selectivity compared to AR-12 (Yang et al., 2018). The structural modifications of the compound scaffold enhance antiviral potency, facilitating progression to clinical trials.
AKT is a central pathway in cell metabolism and participates in various cell actions related to virus infection (Albentosa-Gonzalez et al., 2021; Cuartas-Lopez et al., 2018; Okamoto et al., 2017). The antiviral effect of AT13148 on the DENV-2 replication cycle is investigated on Huh7 cells, which represent a physiologically relevant cell line that can be used to represent liver damage associated with DENV infection in vitro (Tafere et al., 2020). The RT-qPCR and flow cytometry results showed that AT13148 did not influence the attachment and endocytosis steps of DENV-2. Although the AKT pathway was shown to modulate the actin and cadherin actions that are associated with virus entry, AT13148 acts on postentry steps of the DENV-2 replication cycle. After DENV-2 infection, the data showed that AT13148 attenuated viral RNA replication as early as 12 hpi and also inhibited the translation of the viral nonstructural protein NS3 and structural protein capsid in DENV-2-infected Huh7 cells. Moreover, the infectious virions in the cultural supernatant were also reduced when treated with AT13148. Since the Flavivirus harbors some replication strategy, AT13148 is likely to inhibit other Flavivirus infections with broad-spectrum antiviral effects, which need to be investigated on JEV, ZIKV, YFV, and WNV infections in the future study.
Posttranslational modification plays an important role in regulating kinase activity. Previous studies show that pAKT Th308 and Ser473 are detected as early as 1–5 min after DENV-2 infection and maintain cell survival by antagonizing the apoptosis signal (Cuartas-Lopez et al., 2018). The level of pAKT Ser473 in AT13148-treated cells is upregulated in DENV-2-infected cells. This is consistent with previous studies that AKT inhibitors can lead to hyperphosphorylated AKT and downregulated kinase activity (Peeters et al., 2022). The hyperphosphorylation has been seen with other ATP-competitive inhibitors of AKT and shown to be because of direct inhibitor binding, rather than a feedback pathway. Vimentin is the downstream effector of AKT (Zhu et al., 2011). In this study, hyperphosphorylated AKT inhibits the pVimentin Ser39 and Ser83 in AT13148-treated cells. AR-12 was shown to significantly downregulate the GRP78 expression and PI3K/AKT activity in DENV-infected cells (Chen et al., 2017). However, our studies show that despite the hyperphosphorylated AKT, the GRP78 protein level remains unchanged when infected Huh7 cells were treated with AT13148. This could be due to the different action of AT13148 on AKT.
In conclusion, our study identified the antiviral activity of AT13148 in DENV-2 replication. We demonstrated that AT13148 suppresses DENV-2 infection in the postentry step by inhibiting vimentin phosphorylation by targeting the AKT pathway. However, the PI3k/AKT/mTOR pathway is complex in regulating cell metabolism and has multiple substrate signals, and other downstream signals may also contribute to the antiviral activity of AT13148. Further studies are required to define the broad-spectrum anti-Flavivirus activity and test the anti-Flavivirus activity of AKT inhibitors in vivo to evaluate the safety and efficacy.
Footnotes
Authors’ Contributions
Conceptualization: Y.C.D. and Y.F.L.; methodology: C.T.Y., P.J.H., W.Y., and Y.F.L.; data analysis: J.Y.; writing—original draft preparation: Y.C.D.; writing—review and editing: Z.K.X., F.L.Z., and Y.F.L.; and funding acquisition: Y.C.D. and P.J.H. All authors have read and agreed to the published version of the article.
Author Disclosure Statement
The authors declare no conflicts of interest.
Funding Information
The present study was supported by grants from the National Natural Science Foundation of China (Nos. 81702003 and 81801995) and the Key Research and Development Program of Shaanxi Province (No. 2023-ZDLSF-31).