
Abstract
Backgrounds:
Leishmaniasis is one of the neglected tropical diseases, distributed across 89 countries in both the Old and New Worlds. Among the 54 identified Leishmania species, 21 are known to be pathogenic to humans. Cutaneous leishmaniasis (CL) is primarily caused by L. major and L. tropica, while visceral leishmaniasis (VL) in Iran is caused by L. infantum. Accurate detection and species identification of Leishmania spp. are crucial for more effective treatment, epidemiology, and control strategies for the disease. Among the molecular targets recently used for detecting Leishmania species, the heat-shock protein 70 (Hsp70) gene has proven to be highly suitable.
Methods:
This study aimed to establish and evaluate a SYBR Green real-time PCR targeting the Hsp70 gene to identify and differentiate three Leishmania species: L. major, L. tropica, and L. infantum in clinical specimens. A total of 219 microscopic smears, consisting of both positive and negative leishmaniasis cases diagnosed by microscopy, were subjected to DNA extraction and the Hsp70 real-time PCR assay designed in this study.
Results:
Based on the analysis of the melting temperature (Tm) of the amplified Hsp70 target, 115 microscopy-positive smears were identified, comprising 70.4% L. major, 23.5% L. tropica, and 6.1% L. infantum. All results were confirmed using a commercial diagnostic kit. Sanger sequencing of selected positive amplicons unequivocally confirmed the accuracy of this method in identifying and distinguishing the three Leishmania species.
Conclusions:
The Hsp70 real-time PCR can be considered an effective method for detecting and identifying Leishmania species from microscopic slides prepared from CL and VL cases in different regions of Iran.
Introduction
Protozoan parasites belonging to the genus Leishmania are the causative agents of leishmaniasis (Steverding, 2017). The genus comprises three subgenera, each containing various complexes, with each complex consisting of multiple species responsible for different forms of leishmaniasis (Ramírez et al., 2016). To date, a total of 54 species of Leishmania have been identified, of which 21 are known to be pathogenic to humans (Bañuls et al., 2007). Leishmaniasis has a worldwide distribution and is endemic in various regions, including Asia, Africa, the Mediterranean, and the Americas. Currently, more than 89 countries are affected by this disease, with an estimated 12 to 15 million individuals infected. Additionally, 350 million people remain at risk of contracting the disease, and approximately 1.5 to 2 million new cases are reported annually (Torres-Guerrero et al., 2017).
Leishmaniasis ranks second among tropical infectious diseases in terms of prevalence and fourth in terms of mortality rates (Bern et al., 2008). The disease manifests in several clinical forms, depending on the site of parasite infection, including cutaneous, mucocutaneous, visceral, diffuse cutaneous, and postkala-azar dermal leishmaniasis. Cutaneous leishmaniasis is the most common form, with over 90% of cases reported in countries such as Iran, Afghanistan, Saudi Arabia, Syria, Algeria, Tanzania, Brazil, and Peru. Visceral leishmaniasis, meanwhile, predominantly affects regions in Asia (Bangladesh, India, Nepal), Africa (Ethiopia, Kenya, Sudan), and the Americas (northeastern Brazil), accounting for more than 90% of the global reported cases (Akhoundi et al., 2017; Alemayehu and Alemayehu, 2017; Torres-Guerrero et al., 2017).
In West Asia, leishmaniasis is primarily caused by L. infantum and L. donovani, which are responsible for visceral leishmaniasis, and L. major and L. tropica, which cause cutaneous leishmaniasis. In Iran, cutaneous leishmaniasis due to L. major and L. tropica is the most significant form of the disease, with nearly 20,000 cases reported annually (Ghatee et al., 2020). L. infantum is the causative agent of visceral leishmaniasis, also known as kala-azar, which can be fatal if left untreated. Kala-azar ranks second in mortality and seventh in disability-adjusted life years among infectious and tropical diseases. In Iran, visceral leishmaniasis is endemic in the south and northwest, with about 100 symptomatic cases reported each year (Mohebali et al., 2023). In recent years, cases of cutaneous leishmaniasis caused by L. infantum have also been reported in the country (Sabzevari et al., 2021).
Accurate diagnosis of specific types of leishmaniasis is crucial for implementing appropriate therapeutic interventions and predicting disease outcomes. Given the wide range of nonhuman reservoir hosts for Leishmania and the expansion of disease vectors, precise identification of Leishmania species is critical for controlling the disease and understanding the complex epidemiological relationships between the parasite, host, reservoir, and vector. Multilocus enzyme electrophoresis has traditionally been the primary method for diagnosing Leishmania species (Rioux et al., 1990). However, in recent years, molecular markers based on nucleotide sequences of specific genes have emerged as more reliable and widely accepted diagnostic tools (Reithinger and Dujardin, 2007). These molecular markers have proven superior in accuracy and are gaining prominence in the diagnosis of Leishmania. More recently, real-time PCR has significantly advanced the detection and identification of Leishmania species. This method offers a rapid response time, greater analytical sensitivity compared to conventional PCR, and eliminates postPCR steps, thereby reducing contamination risks (de Morais et al., 2016; Galluzzi et al., 2018). Among the commonly targeted genes for species determination and taxonomic/phylogenetic studies is the heat-shock protein 70 (Folgueira et al., 2007) gene (Derghal et al., 2022). It has been demonstrated that Hsp70 gene polymorphism can effectively distinguish between different Leishmania species (Folgueira et al., 2007; Zampieri et al., 2016). Notably, the gene exists in multiple copies within the Leishmania genome and has strong phylogenetic capabilities (Akhoundi et al., 2017).
The primary objective of this study was to utilize the distinctive capabilities of the Hsp70 gene to identify three specific species: L. major, L. tropica, and L. infantum. We aimed to optimize a specific and rapid assay with high sensitivity for the diagnosis of Leishmania infection and for species identification directly from clinical samples.
Materials and Methods
In silico analysis
Reliable reference sequences of the Hsp70 gene from L. major, L. tropica, L. infantum, and some other Leishmania species were obtained from GenBank (http://www.ncbi.nlm.nih.gov/genbank/). In addition, homologous Hsp70 sequences from microorganisms that could potentially be present in clinical samples of patients suspected of leishmaniasis, as well as Hsp70 sequences from humans and Phlebotomus spp. (leishmaniasis vectors), bacteria, fungi, and parasites, were included in the analysis. All sequences were aligned using Geneious Prime® software (https://www.geneious.com). In this study, specific forward and reverse primers targeting the Leishmania-specific Hsp70 gene were designed in-house to allow for the specific amplification of the Hsp70 gene using SYBR Green real-time PCR. This enabled the differentiation of L. infantum, L. major, and L. tropica based on melting temperature analysis. The predicted melting temperatures of the amplicons from the designed primers were evaluated in silico using Geneious Prime® software and the OligoCalc website (http://biotools.nubic.northwestern.edu/OligoCalc.html).
Clinical samples
A total of 219 samples were collected, including 212 skin lesion samples from patients suspected of cutaneous leishmaniasis, referred to health care centers in various regions of Iran: Isfahan (77 positive and 71 negative), Kerman (25 positive), and Tehran (6 positive and 33 negative). Additionally, seven positive bone marrow or lymph node samples from patients with visceral leishmaniasis in Ardabil were included. These samples consisted of Giemsa-stained microscopic smears, following routine diagnostic protocols. The samples were categorized into two groups: leishmaniasis-positive (115 samples) and leishmaniasis-negative (104 samples) based on direct microscopy observed by experienced microscopists. The study received ethical approval from the Ethical Committee of Isfahan University of Medical Sciences (IR.MUI.RESEARCH.REC.1399809).
DNA extraction and PCR amplification
DNA was extracted and purified from the slide smears using the Cellavand® DNA extraction kit. Briefly, an area of approximately 0.5 cm2 from each sample was scraped using a cover glass. The sample was collected into 1.5 mL tubes with 200 µL of the kit’s lysis buffer, incubated at 70°C for 30 min, followed by 95°C for 10 min. After incubation, 200 µL of precipitation buffer and 400 µL of absolute ethanol were added, and centrifuged at 10,000 rpm. The supernatant was discarded, and 500 µL of washing buffer was added to the sediment, followed by centrifugation at 10,000 rpm for 2 min. After discarding the supernatant and drying the sediment, 150 µL of elution buffer was added, and the samples were stored at −20°C until further use for PCR.
Real-time PCR was performed in a final volume of 30 µL, consisting of 0.3 µM of each primer (MITHSP-F: 5′-ATGACCAAGATGTACCAGAG-3′ and MITHSP-R: 5′-ACCGCTCATGCCGCTCAT-3′), designed in this study to target the Leishmania-specific Hsp70 gene. The reaction included 10 µL of extracted DNA and 15 µL of 2x SYBR Green master mix (Cellavand®, Isfahan, Iran). The PCR protocol began with a 3-minute predenaturation step at 95°C, followed by 40 cycles of amplification: 10 s at 95°C, 30 s at 50°C, and 30 s at 72°C. Melting temperature analysis was conducted from 80°C to 97°C, with 1°C increments per second. All samples, as well as water as the negative control and L. major MRHO/IR/75/ER, L. tropica MHOM/IR/02/Mash10, and L. infantum MCAN/IR/14/M14 (obtained from culture) as positive controls, were tested in duplicate. DNA extraction for the positive controls followed the same procedure.
To validate the SYBR Green real-time PCR results, all positive and negative samples were tested using the commercial Cellavand Leishmania diagnostic kit® (https://cellavand.com). This kit uses TaqMan probe-based real-time PCR for detecting and identifying Leishmania, employing four distinct fluorescent oligonucleotides: pan-Leishmania species (FAM), L. major (FAM), L. tropica (HEX), and L. infantum (HEX), designed to amplify specific regions of rDNA. For each sample, two separate duplex reactions were conducted: one for L. major and L. tropica and another for L. infantum and pan-Leishmania. Each reaction consisted of a final volume of 30 µL, using 10 µL of DNA template.
To confirm the accuracy of the SYBR Green real-time PCR results, one positive amplicon for each species (L. major, L. tropica, and L. infantum) was subjected to Sanger sequencing (Core Facilities, Isfahan University of Medical Sciences, Isfahan, Iran). Positive samples were selected based on electrophoresis of several cases on 2% agarose gel, with those showing sharp bands and no smearing or nonspecific amplicons chosen for sequencing. The resulting sequences were aligned with reference sequences in GenBank using the BLAST program (https://blast.ncbi.nlm.nih.gov/Blast.cgi).
In this study, direct microscopic examination of Giemsa-stained smears by professional microscopists was considered the gold standard for determining the sensitivity and specificity of the developed real-time PCR. The sensitivity and specificity of the melting temperature analysis for identifying L. major, L. infantum, and L. tropica were also compared with those obtained using the Cellavand Leishmania and Leishmaniasis Diagnostic kit®.
Results
Following in silico analysis and multiple sequence alignment of the Hsp70 gene across various Leishmania species, a pair of primers was selected for the detection and identification of L. major, L. tropica, and L. infantum using real-time PCR based on melting temperature analysis (Fig. 1). When applied in practical PCR experiments, the average melting temperatures for L. major, L. tropica, and L. infantum were observed as 90.77°C (SD: 0.05), 89.79°C (SD: 0.05), and 87.79°C (SD: 0.04), respectively. Positive control results confirmed that the specific melting temperatures of each species allowed accurate identification, verifying the primer set’s ability to distinguish the target species. Further evaluation of the designed primers revealed a primer dimer with a melting temperature of 78.5°C, which was significantly lower (9–12°C difference) than those of the true positive samples. This distinct melting curve pattern exhibited no overlap with the true positive results, allowing clear differentiation between species by a 1–2°C margin.
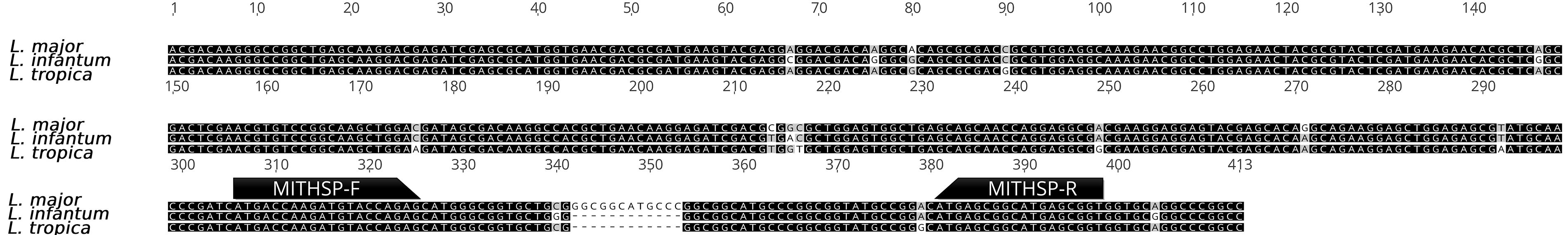
Comparison of HSP70 gene sequences of L. major, L. infantum, and L. tropica and the position of the selected primers used in real-time PCR based on melting temperature analysis.
The melting temperature analysis of the amplified Hsp70 target using these primers confirmed that all external negative controls were negative. These controls included DNA from fungi (Candida albicans, C. parapsilosis, C. tropicalis, Trichophyton rubrum, T. interdigitale, T. mentagrophytes, Aspergillus fumigatus, A. terreus, Rhizopus oryzae, R. microspores, Lichtheimia sp., and Mucor sp.), parasites (Acanthamoeba sp., Plasmodium berghei, Echinococcus granulosus, and Toxoplasma gondii), bacteria (Serratia marcescens, Staphylococcus saprophyticus, S. aureus, and Klebsiella sp.), and normal human blood samples.
In the clinical assay of 219 specimens from patients, no specific amplification was observed in all 104 samples that were negative by direct microscopy. However, 115 samples that were positive by direct microscopy were confirmed as Leishmania based on their melting curve patterns. Species identification using Hsp70 melting temperature analysis identified 81 samples (70.4%) as L. major, 27 samples (23.5%) as L. tropica, and 7 samples (6.1%) as L. infantum. Figure 2 provides examples of melting curve patterns generated from the amplification of DNA extracted from smear samples of infected patients using the developed real-time PCR assay. The results of the melting temperature analysis were further validated through sequencing of three selected PCR amplicons, which matched the reference sequences already deposited in GenBank under accession numbers MF137828.1, LC791486.1, and CP086051.1, corresponding to L. infantum, L. tropica, and L. major, respectively. Additionally, a commercial leishmaniasis diagnostic kit successfully identified all 115 positive microscopic samples and also identified 3 samples out of the 104 negative microscopic samples as positive (Table 1).
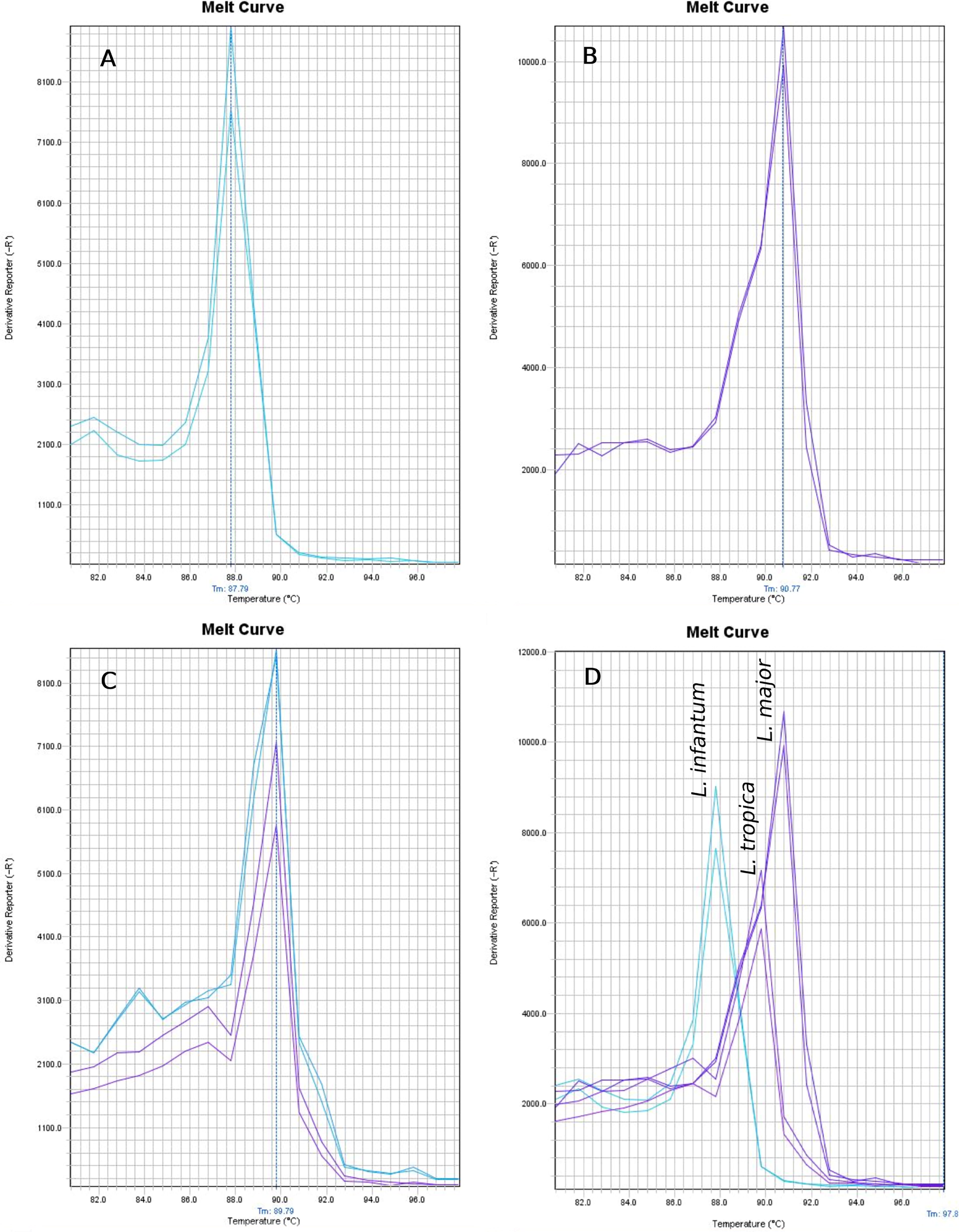
Melting temperature curve after partial amplification of Hsp70 gene.
The Results of Identification of L. major, L. tropica and L. infantum by SYBR Green Real Time PCR Followed by Melting Temperature Analysis of Hsp70 Gene Compared to the Results Obtain from Parasitology Direct Microscopy
Species identification of Leishmania spp. was performed using both Hsp70 melting temperature analysis and the commercial diagnostic kit, as shown in Table 2. All positive samples from Isfahan were identified as L. major, all positive samples from Ardabil were diagnosed as L. infantum, 23 out of 25 samples from Kerman were L. tropica and 2/25 were L. major, and 4 out of 6 samples from Tehran were L. tropica and 2/6 were L. major. Using parasite identification in stained positive and negative samples as the gold standard for leishmaniasis diagnosis, the sensitivity and specificity of the single real-time PCR targeting Hsp70 were calculated to be 100%. The sensitivity and specificity of species identification by melting temperature analysis were also compared with the diagnostic kit results, yielding a sensitivity of 96.43% for L. major and 100% for L. infantum and L. tropica, with a specificity of 100% for all three species.
Comparison of Leishmania Species Identification Results Using Hsp70 Gene Melting Curve Analysis and the Commercial Leishmania and Leishmaniasis Diagnostic Kit
Discussion
In the present study, we designed, developed, and evaluated a straightforward and reliable real-time PCR approach based on the melting curve of an appropriate genetic target (Hsp70) for both the detection and identification of the main causative agents of cutaneous and visceral leishmaniasis, namely L. major, L. tropica, and L. infantum. To improve the simplicity of the PCR amplification, a single pair of primers was designed for use in a single reaction tube to detect/identify all three species. The presence of multiple copies of the Hsp70 gene and sufficient variation in the sequences of the three species, along with the establishment of an appropriate and easy-to-perform qPCR assay, made this diagnostic approach desirable. In agreement with this study, several reports have introduced Hsp70 as a suitable target for the differentiation of Leishmania species. Some studies have reported a high-resolution melting (HRM) analysis of the Hsp70 target to identify different species of Leishmania (Zampieri et al., 2016; Filgueira et al., 2020; Ferreira et al., 2021; Derghal et al., 2022; Filgueira et al., 2023). In the study by Derghal et al., L. major, L. tropica, and L. infantum were differentiated by different melting temperatures of 70.60 ± 0.07, 69.92 ± 0.03, and 70.82 ± 0.03, respectively (Derghal et al., 2022). Likewise, Zampieri et al. were able to separate L. major with a Tm of 85.23 ± 0.03 and L. tropica with a Tm of 84.40 ± 0.02 (Zampieri et al., 2016). Some other studies that have targeted Hsp70 to identify Leishmania species either did not include any of the three species used in our study or identified only one of them (Hernández et al., 2014; León et al., 2017; Salvioni Recalde et al., 2019). In another study, the internal transcribed spacer 1 (ITS1) in rDNA was used to identify Leishmania spp., which showed the melting temperatures of 81.8–82.9, 80.2–80.8, and 79.5–80.2 for L. major, L. tropica, and L. infantum, respectively (de Almeida et al., 2017). Although HRM analysis has been successfully used for the identification of Leishmania species, there are limitations to the widespread use of this method in diagnostic laboratories. For example, the insufficient difference in melting temperature for different species may lead to overlap in the curves due to changes in reagents, PCR machines, etc., making the interpretation of results difficult. Other limitations include difficulties in the availability of a suitable HRM kit, as well as a specific real-time PCR machine and high-resolution-specific software for differentiating species with a melting temperature difference of less than 0.5 degrees. Notably, in the method established in the present study, the difference in melting temperatures between the three common species in Iran, namely L. major, L. tropica, and L. infantum, was sufficient for easy distinction of the species and does not have the limitations of the HRM analysis method.
Among the considerations of the real-time PCR melting temperature analysis method used in this study is the necessity of using a DNA extraction method with high-quality purification to minimize the factors affecting the melting temperature changes in different samples. Another consideration is the lower sensitivity of the Hsp gene in terms of the number of copies compared to genetic regions such as ribosomal DNA and kinetoplast DNA, which may cause false-negative results in cases with low parasite loads. The commercial kit reported 2.9% of the samples that were negative by direct observation as positive. This discrepancy may be due to the greater sensitivity of the kit compared to microscopy. Although the microscopic observation of parasites is known as the gold standard for diagnosis, it increases problems such as fatigue of the microscopist after observing several samples and the possibility of reporting false-negative cases. The comparison of the commercial kit and the real-time PCR assay introduced in this study shows that the commercial kit had a higher sensitivity in the diagnosis of leishmaniasis. This likely depends on the target used in the kit, which has more copies in the Leishmania genome compared to the Hsp70 target, as well as the reproducibility and higher efficiency of the reagents used in the diagnostic kit.
L. major is the cause of cutaneous leishmaniasis in Isfahan, and the obtained results were expected. This finding is also true for Ardabil samples, where all examined samples were identified as L. infantum, which is the cause of visceral leishmaniasis in the region. While two cases of L. major were detected in the samples from Kerman province, considering the presence of both species, L. tropica and L. major, in this province, the obtained results are consistent with the epidemiology of Leishmania.
Conclusion
Since rapid and accurate detection and identification of Leishmania species are crucial for clinical and epidemiological purposes, in this study, the heat-shock protein 70 (Hsp70) gene was successfully targeted for a SYBR Green real-time PCR assay to detect L. major, L. tropica, and L. infantum in clinical specimens. The difference in the established melting temperature was sufficient for the easy distinction of the three species. The assay can be considered an effective method for identifying Leishmania species on microscopic slides prepared from cutaneous or visceral leishmaniasis cases in different parts of Iran.
Footnotes
Author Disclosure Statement
No competing financial interests exist.
Funding Information
This study was conducted with the financial support of Isfahan University of Medical Sciences (Grant number 199655). The Core Research Facilities at Isfahan University of Medical Sciences provided effective cooperation in advancing this study. The authors thank Professor Iraj Sharifi from Kerman University of Medical Sciences and Dr. Zarei from Tehran University of Medical Sciences for supplying some clinical samples.