
Abstract
Objective:
Anopheles mosquitoes transmit Plasmodium parasites through blood feeding. The oocyst stage in mosquitoes is crucial for Plasmodium transmission. Oocysts can form without a blood meal or the midgut passage and utilize nutrients from additional blood feeding. However, it remains unclear the impact of a blood meal during infection on oocysts. The present study evaluated how a blood meal during infection impacts oocyst growth to better understand oocyst development.
Methods:
We used a novel method for Plasmodium infection known as enema injection, which involves injecting Plasmodium berghei (ANKA strain) into the midgut lumen of Anopheles stephensi mosquitoes (STE2 strain) via the anus, traversing the midgut wall without a blood meal. We compared the size of oocysts in mosquitoes infected by enema injection alone, hemocoel injection alone, and each infection method combined with uninfected blood feeding, as well as those only with infected blood feeding.
Results:
By enema injection with Plasmodium ookinetes, oocysts formed solely in the mosquito’s midgut. Oocysts from enema-injected mosquitoes were similar in size to those from hemocoel-injected mosquitoes. Oocysts from mosquitoes infected by enema injection combined with uninfected blood feeding were larger than oocysts from mosquitoes infected by enema injection alone. However, the size of oocysts from mosquitoes infected by hemocoel injection was not affected by the presence or absence of blood feeding.
Conclusion:
Enema injection with Plasmodium ookinetes is applicable to Anopheles mosquitoes. Using the enema injection method, we suggest that a blood meal during infection might facilitate oocyst growth within the midgut.
Introduction
Anopheles mosquitoes transmit Plasmodium parasites, causing malaria. The Plasmodium life cycle involves mosquitoes and vertebrate hosts. It begins with the mosquito ingesting infected blood, leading to the transformation of gametocytes into motile ookinetes in the midgut. These ookinetes pass through midgut epithelial cells and become oocysts under a basal lamina within 2 days after blood feeding. The oocysts enlarge over time, forming numerous sporozoites inside. After more than 12 days, mature oocysts rupture, releasing sporozoites into the hemocoel. These sporozoites enter the salivary glands and are transmitted to a new vertebrate host during the next blood feeding (Singh et al., 2021; Smith et al., 2014). The oocyst stage is critical, as it produces sporozoites capable of infecting the vertebrate host.
Prior research examined the role of bloodmeal and the midgut passage in oocyst development using hemocoel injection, in vitro culture of Plasmodium parasites, and additional blood feeding. The hemocoel injection method, in which ookinetes are injected into the mosquitoes’ hemocoel, bypasses both the blood meal and the midgut passage. Injected ookinetes subsequently transform into oocysts within the mosquito (Alavi et al., 2003; Deligianni et al., 2018; Haraguchi et al., 2023; Nacer et al., 2008; Paskewitz and Shi, 2005; Weathersby, 1952). In addition, oocysts form without basal lamina components in vitro (Carter et al., 2007; Nacer et al., 2008). These findings demonstrate that blood meal and the midgut passage are unnecessary for oocyst formation.
Additional blood feeding, taken after oocyst formation, has been shown to promote oocyst growth. It is believed that a blood meal rich in nutrients, such as amino acids and lipids, supplies oocysts with resources for their growth (Habtewold et al., 2021; Kwon et al., 2021; Shaw et al., 2020). However, it remains unclear whether the blood meal taken during infection, prior to oocyst formation, affects oocyst growth. If confirmed, this could indicate a novel possibility that Plasmodium parasites possess a cross-stage energy storage mechanism. Such a discovery would be crucial for advancing our understanding of the Plasmodium life cycle.
The enema injection method, used in other research fields, mimics the natural infection route by introducing pathogens into the mosquito’s midgut via the anus (Klowden, 1981; Nuckols et al., 2013; Putnam and Scott, 1995). No reports exist of Plasmodium infection in Anopheles mosquitoes through enema injection. However, this method provides a novel condition in which ookinetes pass through the midgut wall without a bloodmeal, an advantage. This method is considered the only way to deliver Plasmodium parasites directly into the midgut without a blood meal, as a blood-immiscible Plasmodium mixture, when orally fed, typically enters the crop before flowing into the midgut (Calkins et al., 2017).
We evaluated the effects of a bloodmeal during infection on Plasmodium oocyst growth. Our findings demonstrated that the enema-injection method is applicable to Plasmodium berghei infection in Anopheles stephensi mosquitoes. Using this method, we identified that an infected bloodmeal in the midgut may facilitate oocyst growth.
Materials and Methods
Ethics statement
Experiments involving animal subjects were conducted in compliance with the national and institutional guidelines for the care and use of laboratory animals. Specifically, the experiments adhered to the Guidelines of Animal Experiments of Kitasato University and received approval from the Kitasato University Institutional Animal Care and Use Committee (approval numbers: 21-031 and 24-049).
Mosquito rearing
An. stephensi (STE2 strain) were bred in an insectary at 19°C, following a 14 h:10 h light–dark cycle, and were provided with a 10% (w/v) sucrose solution. Mosquitoes aged between 3 and 10 days were utilized for the experiments.
Parasites and in vitro culture of ookinetes
We employed green fluorescent protein-expressing P. berghei (ANKA strain). The in vitro culture and purification of ookinetes were carried out according to previous reports (Haraguchi et al., 2023; Nakayama et al., 2021). Briefly, infected blood from Institute of Cancer Research mice (Charles River Laboratories Japan Inc., Yokohama, Japan) was cultured with Roswell Park Memorial Institute-1640 medium (Thermo Fisher Scientific Inc., Waltham, MA, USA), supplemented with 25 mM 4-(2-hydroxyethyl)-1-piperazineethanesulfonic acid (Dojindo Laboratories, Kumamoto, Japan), 0.4 mM hypoxanthine (Sigma-Aldrich, St. Louis, MO, USA), 24 mM NaHCO3 (Fujifilm Wako Pure Chemical Corporation, Osaka, Japan), and 12.5 mg L−1 gentamicin reagent solution (Thermo Fisher Scientific Inc.). This was maintained at 19°C for a duration of 20–24 h. A MidiMACS separator system (LS Columns; Miltenyi Biotec, Bergisch Gladbach, Germany) was utilized to purify ookinetes from the culture.
Enema injection
The enema injection procedure was conducted according to the method detailed by Nuckols et al., albeit with certain modifications (Nuckols et al., 2013). A female mosquito was anesthetized using a cold temperature method and subsequently positioned supine on a petri dish, which was lined with a paper towel and kept at room temperature. Using a Nanoject II automatic nanoliter injector (Drummond Scientific Company, Broomall, PA, USA), 69 nL of either trypan blue solution or 5,000 ookinetes was administered into the mosquito’s midgut via the anus. For enema injection combined with uninfected blood feeding, the mosquitoes sucked on an uninfected mouse for 30 min within 2 h after enema injection, leading to blood engorgement.
Hemocoel injection and infected blood feeding
Hemocoel injection was carried out in line with the methods described in earlier reports (Haraguchi et al., 2024, 2023). A total of 69 nL of 5,000 ookinetes was introduced into the mosquito’s hemocoel using a Nanoject II automatic nanoliter injector (Drummond Scientific Company). For mosquitoes subjected to hemocoel injection combined with uninfected blood feeding, the ookinetes were injected into the hemocoel within 2 h following a 30-min uninfected blood-feeding session.
Infected blood feeding was executed for a duration of 30 min at room temperature, utilizing anesthetized mice infected with P. berghei, according to the method outlined in a previous study (Nakayama et al., 2021).
Examination of the impact of a bloodmeal during infection on P. berghei oocyst growth in Anopheles mosquitoes
For the infection experiment, a group size ranging from 3 to 50 mosquitoes was utilized. Experimental procedures are outlined in Fig. 1. The mosquitoes were subjected to enema injection, hemocoel injection, a combination of each infection method with uninfected blood feeding, and infected blood feeding. After 14 days of infection, the mosquitoes were dissected and examined using a fluorescence microscope (Leica M205 FA, Leica Microsystems, Wetzlar, Germany). The diameters of the oocysts were measured using ImageJ software. Statistical analyses were conducted using GraphPad Prism version 8.4.3 software (GraphPad Software Inc., San Diego, CA, USA). The significance of the oocyst diameter was determined using a one-way ANOVA test and Tukey’s multiple comparisons test (p < 0.05).
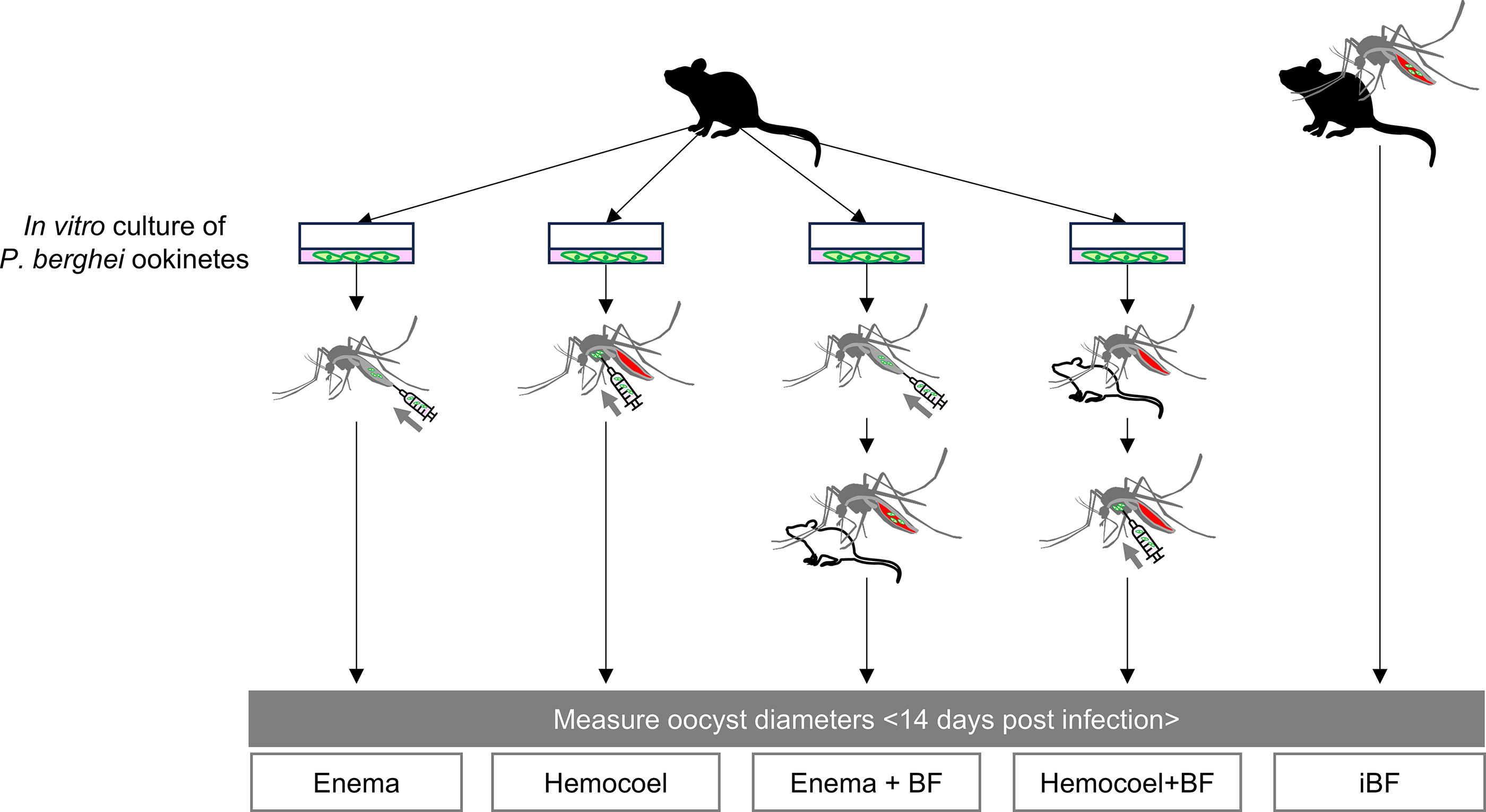
Schematics of the experimental procedure designed to examine the impact of a blood meal during infection on oocyst growth. Anopheles mosquitoes were subjected to five treatments with Plasmodium berghei: enema injection alone (enema), hemocoel injection alone (hemocoel), enema injection combined with uninfected blood feeding (enema + BF), hemocoel injection combined with uninfected blood feeding (hemocoel + BF), and infected blood feeding (iBF). Oocyst diameters in each group were measured 14 days postinfection. A black mouse icon represents a P. berghei-infected source, while a white mouse icon indicates an uninfected source. In the enema, hemocoel, enema + BF, and hemocoel + BF groups, P. berghei ookinetes cultured in vitro were used for infection.
We first compared the oocyst diameters across all mosquito groups. The total number of dissected mosquitoes was 9 for enema injection, 31 for hemocoel injection, 4 for enema injection with uninfected blood feeding, 11 for hemocoel injection with uninfected blood feeding, and 11 for infected blood feeding. One experiment involved two or three groups selected from the five groups. Each group was tested more than twice, except for the hemocoel injection with blood-feeding group, which was tested only once. Next, we compared oocyst diameters within mosquitoes based on oocyst number ranges. In the infected blood-feeding group, seven mosquitoes had 1–100 oocysts, two had 101–200 oocysts, and two had >200 oocysts. In the hemocoel-injection group, six mosquitoes had 1–100 oocysts, one had 101–200 oocysts, and six had >200 oocysts. Finally, mosquitoes with <200 oocysts in each group were used to compare oocyst diameters. The number of mosquitoes in each group was as follows: 9 in the enema injection group, 26 in the hemocoel injection group, 3 in the enema injection with uninfected blood-feeding group, 10 in the hemocoel injection with uninfected blood-feeding group, and 8 in the infected blood-feeding groups.
Results
Enema injection with P. berghei was able to apply to Anopheles mosquitoes
We conducted an enema injection using a trypan blue solution to ascertain the distribution of the injected solution within mosquitoes. The abdomens of the mosquitoes injected with the trypan blue solution exhibited a blue stain (Fig. 2A, B). The trypan blue solution was observed within the midgut and showed no leakage into the hemocoel (Fig. 2C).
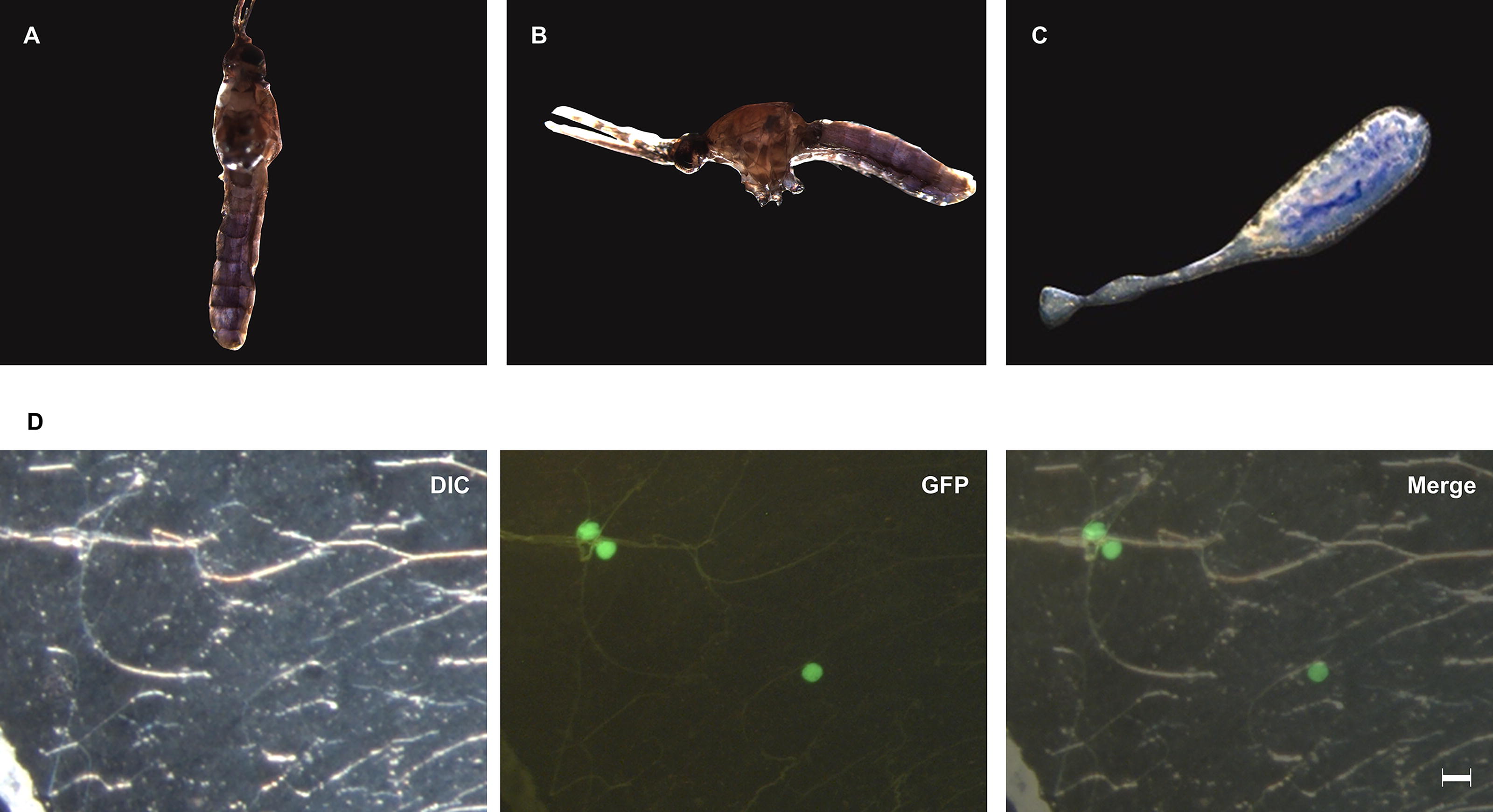
Enema injection into mosquitoes.
Next, we examined whether P. berghei could form oocysts in An. stephensi through enema injection with 5,000 in vitro cultured ookinetes. In the mosquitoes subjected to enema injection with ookinetes, oocysts were formed only at the midgut (Fig. 2D). The mean survival rate was lower in mosquitoes subjected to enema injection (30.8%) compared with those subjected to hemocoel injection (60.3%, Table 1). The prevalence was also lower in mosquitoes subjected to enema injection (20.0%) compared with those subjected to hemocoel injection (48.6%, Table 1).
Comparison of Infections Between Enema and Hemocoel Injection
Blood meal during infection influences oocyst growth of P. berghei in the Anopheles midgut
We examined the effect of a blood meal on oocyst growth in mosquitoes, using five treatments. The mean oocyst diameters were 18.80 ± 9.38 µm in the mosquitoes with enema injection alone, 18.70 ± 10.37 µm in those with hemocoel injection alone, 32.02 ± 7.61 µm in those with enema injection combined with uninfected blood feeding, 17.86 ± 8.84 µm in those with hemocoel injection combined with uninfected blood feeding, and 18.32 ± 12.46 µm in those with infected blood feeding (Fig. 3). Mosquitoes with enema injection combined with uninfected blood feeding had significantly larger oocysts than the other four treatments (p < 0.01, Fig. 3). No significant difference was observed in oocyst diameter among the other four treatments (Fig. 3).
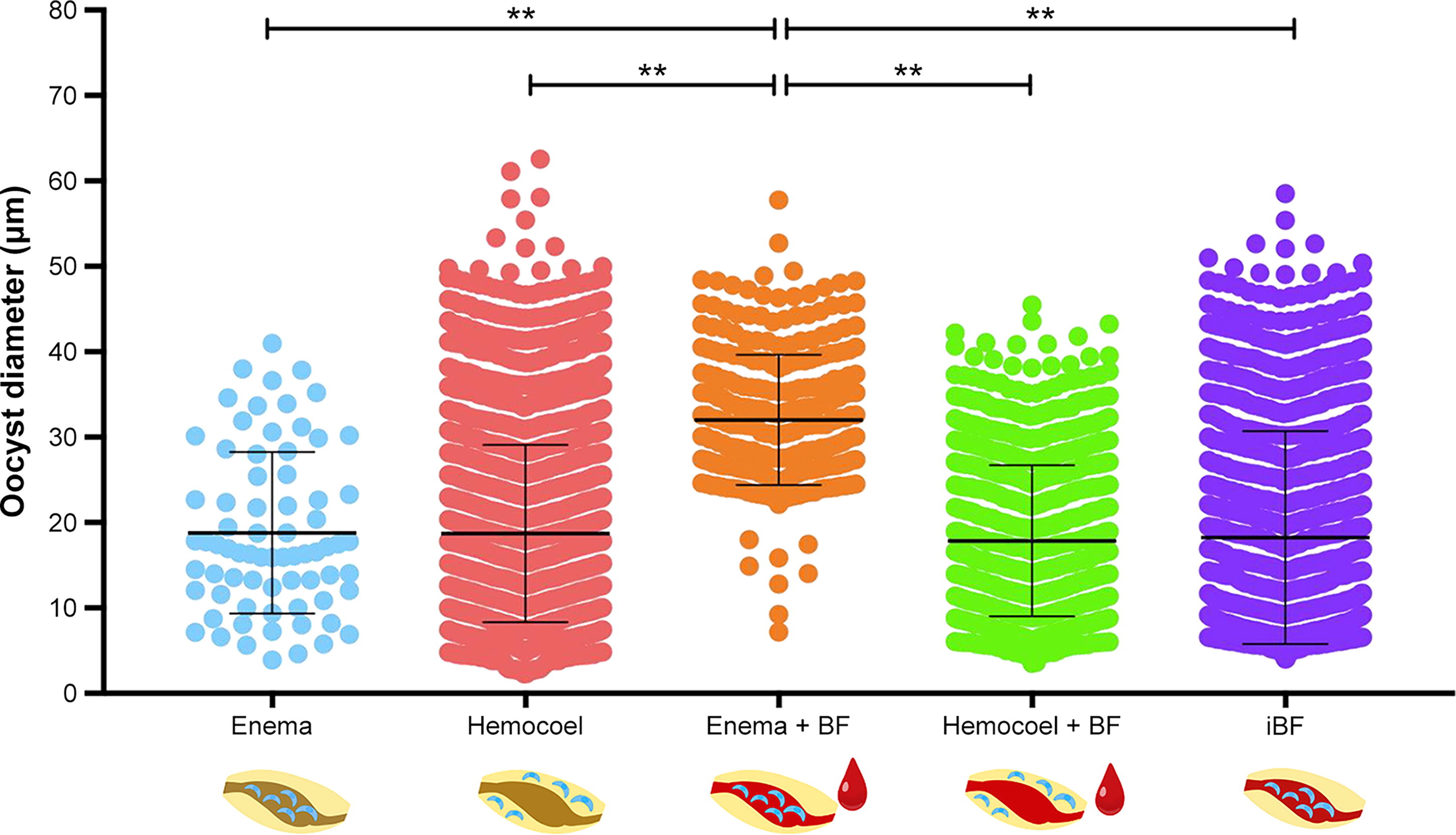
Oocyst diameters in mosquitoes subjected to five treatments. The oocyst diameters of Plasmodium berghei in Anopheles stephensi mosquitoes subjected to enema injection alone (enema), hemocoel injection alone (hemocoel), enema injection combined with uninfected blood feeding (enema + BF), hemocoel injection combined with uninfected blood feeding (hemocoel + BF), and infected blood feeding (iBF) are presented. The oocyst diameters were compared using a one-way analysis of variance (ANOVA) test and Tukey’s multiple comparisons test (**p < 0.01, ***p < 0.001, ****p < 0.0001). A bold horizontal line represents the mean value, and error bars represent the standard deviation.
The number of oocysts exhibited considerable variation when oocysts were formed in mosquitoes via hemocoel injection alone and infected blood feeding (Fig. 4A). A previous study indicated a decrease in oocyst diameter as the number of oocysts increases (Habtewold et al., 2021). Consequently, to address the concern that the quantity of oocysts might impact their size, we investigated the relationship between the number of oocysts per mosquito and the oocyst diameter by examining mosquitoes at 14 days postinfection, which were infected either by blood feeding or hemocoel injection. Mosquitoes infected through enema injection were not included in this analysis because the number of oocysts formed was less than 100 in all instances.
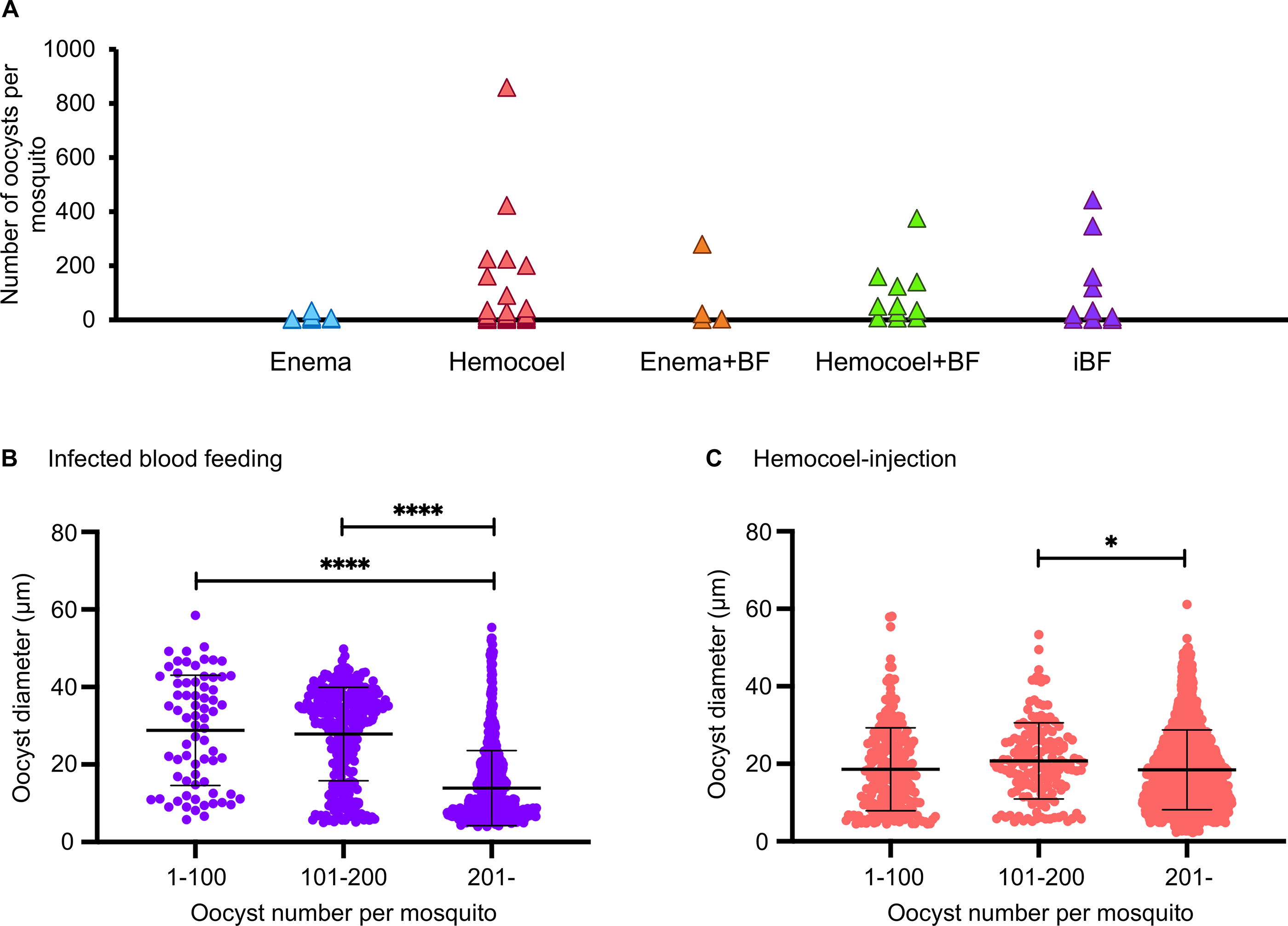
Relationship between the number of oocysts per mosquito and oocyst diameters in mosquitoes infected through blood feeding or hemocoel injection.
In the case of infected blood feeding, the mean oocyst diameters were 28.81 ± 14.13 µm in mosquitoes with 1–100 oocysts, 27.89 ± 12.03 µm in those with 101–200 oocysts, and 13.91 ± 9.68 µm in those with more than 200 oocysts (Fig. 4B). For hemocoel injection, the mean oocyst diameters were 18.61 ± 10.68 µm in mosquitoes with 1–100 oocysts, 20.77 ± 9.82 µm in those with 101–200 oocysts, and 18.46 ± 10.28 µm in those with more than 200 oocysts (Fig. 4C). The oocyst diameters in mosquitoes with more than 200 oocysts were significantly smaller than those in mosquitoes with fewer than 200 oocysts in the infected blood-feeding and hemocoel injection groups (p < 0.05, Fig. 4B, C). Consequently, we opted to use mosquitoes with fewer than 200 oocysts for subsequent oocyst diameter comparisons.
Drawing on the results from Fig. 4, we reanalyzed the data presented in Fig. 3. For mosquitoes with fewer than 200 oocysts, the mean oocyst diameters were 18.80 ± 9.38 µm for those with enema injection alone, 19.57 ± 10.62 µm for those with hemocoel injection alone, 27.98 ± 11.48 µm for those with enema injection combined with uninfected blood feeding, 18.53 ± 8.85 µm for those with hemocoel injection combined with uninfected blood feeding, and 28.08 ± 12.49 µm for those with infected blood feeding (Fig. 5). There was no significant difference in oocyst diameter between mosquitoes subjected to enema injection alone and those subjected to hemocoel injection alone (p = 0.9768, Fig. 5). Notably, the diameter of oocysts in mosquitoes infected through enema injection combined with uninfected blood feeding was larger than those in mosquitoes infected through enema injection alone (p < 0.01) and comparable with those in mosquitoes subjected to infected blood feeding (p > 0.9999, Fig. 5). There was no significant difference in oocyst diameter between mosquitoes infected through hemocoel injection, regardless of whether they had uninfected blood feeding or not (p = 1.041, Fig. 5). Moreover, oocysts formed in mosquitoes subjected to either enema injection or hemocoel injection were smaller than those formed in mosquitoes fed infected blood (p < 0.0001, Fig. 5).

Impact of blood meal during infection on oocyst growth. The diameters of Plasmodium berghei oocysts were compared in Anopheles stephensi mosquitoes subjected to enema injection alone (enema), hemocoel injection alone (hemocoel), enema injection combined with uninfected blood feeding (enema + BF), hemocoel injection combined with uninfected blood feeding (hemocoel + BF), and iBF using a one-way ANOVA test and Tukey’s multiple comparisons test (**p < 0.01, ***p < 0.001, ****p < 0.0001). Only mosquitoes with fewer than 200 oocysts were selected for this analysis. A bold horizontal line represents the mean value, and error bars represent the standard deviation.
Discussion
In the present study, we aimed to investigate the impact of a bloodmeal during infection on the oocyst growth of Plasmodium parasites. Initially, we demonstrated that the enema injection method can be applied for P. berghei infection in An. stephensi mosquitoes. Subsequently, we compared oocyst diameters using the enema injection method to examine the impact of a bloodmeal during infection on oocyst growth.
We first applied enema injection, a method where the solution is injected into the midgut through the mosquito’s anus, to An. stephensi mosquitoes with trypan blue. The confinement of trypan blue within the midgut demonstrated that the enema-injected solution could effectively deliver its contents to the lumen of the mosquitoes’ midgut. Oocysts were exclusively formed in the midgut of mosquitoes subjected to enema injection with P. berghei ookinetes, similar to those in mosquitoes infected through blood feeding. This contrasted with mosquitoes injected into the hemocoel, where oocysts formed throughout the body (Carter et al., 2007). These results suggest that the ookinete mixture can be delivered into the midgut lumen, enabling ookinetes to traverse the midgut cells and transform into oocysts located between the midgut cells and basal lamina in enema-injected mosquitoes.
A previous study suggested that enema injection is less burdensome than hemocoel injection, wherein a glass needle is inserted into the thorax (Nuckols et al., 2013). However, the survival rate was lower in enema-injected mosquitoes compared with hemocoel-injected ones in the present study. Thus, enema injection can be used to study Plasmodium development in Anopheles mosquitoes, although further technical refinement is necessary. For instance, reducing the volume of the injected solution or decreasing the injection speed may improve the survival rate. Lower prevalence in enema injection could be due to ookinetes being exposed to the mosquitoes’ anti-Plasmodium responses during midgut wall invasion. For example, some midgut bacteria produce reactive oxygen species intermediates that damage ookinetes (Cirimotich et al., 2011a, 2011b; Stathopoulos et al., 2014). Damaged midgut cells produce reactive oxygen species and reactive nitrogen to eliminate parasites when ookinetes pass through them (Han et al., 2000; Singh et al., 2021). These mechanisms may explain the reduced prevalence of enema injection compared to hemocoel injection.
Having established that P. berghei infection can be induced through enema injection despite low survival rates, we employed this method to examine the influence of a bloodmeal during infection on oocyst growth. We compared the oocyst growth in mosquitoes infected by enema injection alone, hemocoel injection alone, each injection method combined with uninfected blood feeding, and infected blood feeding. Upon comparing these five groups, the mean oocyst diameter in mosquitoes with enema injection combined with uninfected blood feeding was higher than in the other four groups. However, the number of oocysts varied significantly among these five groups. Consequently, we analyzed the relationship between oocyst numbers and oocyst diameters, suggesting that a larger number of oocysts results in smaller diameters. This relationship has been reported in oocysts infected by blood feeding (Habtewold et al., 2021). Additionally, the present results show that this relationship could also apply to oocysts infected by hemocoel injection.
We reanalyzed oocyst growth while considering the number of oocysts per mosquito. The similarity in oocyst size between the enema- and hemocoel-injected groups suggests that passage through the midgut wall alone does not affect oocyst growth, consistent with previous studies (Carter et al., 2007; Nacer et al., 2008). Notably, oocyst diameters in enema-injected mosquitoes increased with a blood meal, while those in hemocoel-injected mosquitoes were unaffected by the blood meal. These findings suggest that a bloodmeal during infection can influence Plasmodium growth in the midgut but not in the hemocoel.
Previous studies indicate that oocysts utilize additional resources from the mosquito’s additional blood feeding (Habtewold et al., 2021; Kwon et al., 2021; Shaw et al., 2020). For instance, Kwon et al. showed that the size of P. berghei oocysts remains unaffected by additional blood feeding at 4 days postinfection (Kwon et al., 2021). However, Habtewold et al. found that P. berghei oocyst size increases with additional blood feeding at 7 days postinfection (Habtewold et al., 2021). These findings suggest that oocysts in the midphase or later utilize blood meal resources. Our results suggest that the blood meal in the midgut during infection might enable parasites to obtain necessary nutrients for their growth. Thus, we speculate that ookinetes might directly take nutrients from the blood in the midgut lumen, becoming nutrient-rich and promoting oocyst formation under the midgut’s basal lamina, leading to the possibility that Plasmodium parasites might have a cross-stage energy storage mechanism.
The use of mosquito-derived nutrition by Plasmodium parasites varies with the Plasmodium species, Anopheles mosquito species, and their combination (Shaw et al., 2022). Unlike P. berghei, P. falciparum oocysts enlarge with additional blood feeding at 3–9 days postinfection in An. gambiae and An. coluzzii (Habtewold et al., 2021; Kwon et al., 2021). The mosquito’s lipid transporter lipophorin and its transported lipids are essential for P. berghei oocyst growth, while other nutrients like amino acids and carbohydrates are more crucial in P. falciparum (Costa et al., 2018; Habtewold et al., 2021; Rono et al., 2010; Shaw et al., 2022, 2020; Werling et al., 2019). This suggests that the nutrient acquisition mechanism in Plasmodium parasites differs among species. Our study used P. berghei and An. stephensi as models, but future research should explore other combinations.
Conclusions
We initially demonstrated that enema injection can be applied to Plasmodium infection. Utilizing the enema injection method, it is suggested that the blood meal in the midgut during the Plasmodium infection might be vital for oocyst growth. Although the sample size was small and additional research is required, it is proposed that there is a new possibility related to nutrient acquisition of Plasmodium parasites.
Footnotes
Acknowledgments
The green fluorescent protein-expressing P. berghei ANKA strain was provided by Dr. Masao Yuda, Mie University, Japan. An. stephensi (STE2 strain, MRA-128) was provided by BEI Resources. This study would not have been possible without the help of our laboratory support staff member, Mai Tanaka.
Authors’ Contributions
A.H.: Conceptualization, funding acquisition, methodology, visualization, and writing—original draft. M.G.: Conceptualization, formal analysis, investigation, methodology, validation, and visualization. K.N.: Formal analysis, writing—original draft, and writing—review and editing. K.F.: Investigation and methodology. J.H.: Methodology and resources. S.N.: Resources. K.K.: Conceptualization and project administration. H.I.: Conceptualization, funding acquisition, project administration, supervision, and writing—review and editing.
Data Availability Statement
All data generated or analyzed during this study are included in this published article.
Author Disclosure Statement
The authors declare no conflicts of interest.
Funding Information
This study was supported by KAKENHI of the Japan Society for the Promotion of Science [grant numbers 17H04073, 24K01933, 22K19256 (to H.I.), and 23KJ1873 (to A.H.)].