
Abstract
Background:
Zoonotic diseases pose a significant global health challenge, accounting for over half of all newly emerging infectious illnesses. One such pathogenic agent is the bacterium Francisella tularensis, which causes tularemia, a zoonotic disease spread in the Northern Hemisphere through various means, including tick bites. Unfortunately, there is limited literature on tick-borne bacteria affecting human and animal health. This research confirmed the presence of Francisella spp. in ticks in the Urmia region, highlighting the potential for ticks to act as vectors for transmitting the bacteria to livestock and humans.
Materials and Methods:
A total of 397 ticks, 315 blood samples, and 241 milk samples were gathered from sheep and goats to screen for the existence of Francisella spp. The nested PCR technique revealed the existence of Francisella spp. 16S rRNA gene in the samples.
Results:
Blood and milk samples from small ruminants did not show the presence of Francisella spp., but the bacteria were detected in 6 tick pools (n = 79; 7.59%; 95% CI: 3.52%–15.59%). The positive ticks identified were Hyalomma asiaticum 2 (n = 24; 8.33%; 95% CI: 2.31%–25.84%), Hyalomma eagyptum 3 (n = 9; 33.33%; 95% CI: 12.06%–64.58%), and Rhipicephalus sanguineus 1 (n = 15; 6.67%; 95% CI: 1.19%–29.82%) and the presence of Francisella tularensis subspecies holarctica was found to be positive in 3 out of 79 tick pools (3.80%; 95% CI: 1.3%–10.58%). The positive ticks identified in this experiment were female Hyalomma aegyptium (1 out of 3; 33.33%; 95% CI: 6.15%–79.23%), male Hyalomma asiaticum (1 out of 14; 7.14%; 95% CI: 1.27%–31.47%) and female Rhipicephalus sanguineus (1 out of 7; 14.29%; 95% CI: 2.57%–51.32%).
Conclusion:
The study explored the presence of Francisella species, especially the most dangerous, F. tularensis, in small ruminants and the ticks that attach to them using molecular techniques in the Urmia region of northwestern Iran. The research seeks to improve knowledge of Francisella spp. epidemiology. Understanding the risk of human-livestock transmission of this pathogen is crucial for public health, particularly for individuals working in animal-related fields.
Introduction
Recently, there has been an increased focus on Francisella spp. Particularly in the field of vector-borne and zoonotic diseases. Tularemia is caused by the bacterium Francisella tularensis. Francisella spp. is a nonmotile, Gram-negative, pleomorphic coccobacillus and a facultative intracellular pathogen, commonly associated with zoonotic and tick-borne diseases (Maurin and Gyuranecz, 2016). Tularemia could be spread through direct contact, ingestion, and inhalation. Tularemia could be spread via ingestion, inhalation, and direct contact. Tularemia is deemed a waterborne disease owing to its ability to survive for long periods in aquatic environments and the suspected involvement of protozoans in maintaining its aquatic cycle (Hennebique et al., 2019). The bacteria continue to distribute in nature through transfer via rodents or blood-sucking arthropods such as mosquitoes, flies, and ticks (Yeni et al., 2021). Flies are hardly ever involved in Francisella spp. spread to humans, while mosquitoes are primarily involved in specific nations such as Sweden and Finland (Petersen et al., 2009). Ticks can reserve Francisella spp. in nature via transovarial and transstadial transmission. Livestock are possible hosts for infected ticks and may play a role in the cycle of disease (Hestvik et al., 2015).
Tularemia is endemic in some countries of the Northern Hemisphere (Giircan, 2014). Studies on arthropods, human beings, animal populations, and environmental specimens have shown the existence of Francisella spp. in Iran’s neighboring countries, such as Kazakhstan, Turkmenistan, Turkey, Georgia, Pakistan, and Afghanistan (Zargar et al., 2015). Evidence of Francisella spp. infection in humans, wild mammals, and livestock (including sheep, cows, hedgehogs, rodents, and hares) has been recorded in different parts of Iran, including the west, northwest, southwest, north, northeast, and southeast.
In 1972, the first documented cases of Francisella spp. infection in Iran involved 8 sheep, 3 cows, and 1 hedgehog (Arata et al., 1973). Ticks can not only pose a threat to livestock, wild animals, and humans, but they may also serve as a reservoir for Francisella spp. in nature for extended periods (Telford and Goethert, 2020). Research on tick-borne pathogens is crucial to stop and limit the occurrence of relevant diseases in both animals and humans. Tularemia diagnosis cannot rely solely on clinical examinations; microbiological and serological tests are also necessary. These methods, however, pose risks of infection and require extended periods to obtain results (Maurin, 2020). This study aims to use molecular techniques to detect Francisella spp. in ticks as well as in the blood and milk samples of livestock in the Urmia region of Iran.
Materials and Methods
Study area
Urmia, a city in the northwest region of Iran (37° 33′ 10.08″ N, 45° 4′ 33.24″ E), is known for its continental climate, featuring extremely hot, dry summers and cold, icy winters. Both Mediterranean and semiarid climates have an impact on the city’s surroundings. The moderate precipitation during the spring and fall seasons is crucial for the region’s ecosystems and farms. Due to the vast countryside, raising animals is common, and local livestock breeds have adapted well to the climate and geography.
Collection of samples
The specimens were collected from local flocks (sheep and goats) in the counties of Noshin, Talatapeh, Jarchlo, Dastjerd, Yaghmorali, and Gharabghlo from April 19, 2024, to September 19, 2024, in the Urmia region (Fig. 1) (Alizadeh Dizaj et al., 2019). Using a random sampling method, 315 blood samples were obtained from sheep (n = 236; 57 males and 179 females) and goats (n = 81; 19 males and 62 females). All the ruminants were confirmed to be healthy and had not received any treatment with acaricides or antibiotics before the study.
Schematic map of the study areas in West Azerbaijan, Iran, including Urmia.
The blood samples were added to tubes containing ethylenediaminetetraacetic acid and stored at 4°C–8°C until being moved to the laboratory. To maintain sanitary standards and avoid secondary contamination, the animals’ udders were cleaned with warm water and soap, and the teats were sterilized with 70% isopropyl alcohol and dried with a sterile cloth. The first milk obtained was discarded, and 10 milliliters from subsequent milkings were collected in 15-milliliter Falcon tubes. The sum of 241 milk samples were gathered from the examined animals (179 sheep and 62 goats). The milk specimens were divided into three age groups (≤6, 7–10, and >10 years) stored at 4°C–8°C until being sent to the laboratory. For the final sampling, the animals’ skin was thoroughly examined, including the removal of ticks. A small amount of alcohol was sprayed on the area of interest to safely remove ticks for more accurate identification. After a few minutes, ticks were removed from the skin using rotating metal tweezers and placed in tubes with 96% ethanol. In total, 397 ticks were collected from various parts of the animals’ bodies, such as the ears, belly, hind legs, and anal region. These tick samples were then sent to the parasitology lab, where a loupe microscope and dependable diagnostic keys were used to determine the species and genus of adult ticks. Finally, all samples were sent to the Microbiology Laboratory in the Department of Microbiology at the Faculty of Veterinary Medicine, Urmia University, for DNA extraction and molecular testing. The samples were transported in polystyrene coolers filled with ice.
The protocols for all animal experiments were conducted in accordance with the Guidelines for the Care and Use of Laboratory Animals and were supervised by the Animal Care and Use Committee of the Urmia University.
DNA extraction
DNA extraction from blood and milk samples
DNA was extracted from animal blood and milk samples by the Blood Genomic DNA Extraction mini kit Farvogen (Taiwan, 50 preps), following the manufacturer’s instructions carefully. To start the procedure, 200 μL of blood was cautiously transferred into a 1.5 mL microtube. Following the method outlined by Parisi et al. (Parisi et al., 2006). Milk samples that had previously been centrifuged at 5000 rpm for 10 min were used for DNA extraction. The samples were used in the DNA extraction after removing the fat. The extraction was completed utilizing the Blood Genomic DNA Extraction Mini Kit Farvogen (Taiwan, 50 preps) as per the manufacturer’s instructions.
Obtaining DNA from tick samples
To remove any alcohol residues, the ticks were washed three times with phosphate-buffered saline (PBS). After the washing process, they were subjected to air drying, positioned on a sterile textile, and exposed to laboratory atmospheric conditions. Subsequently, according to their respective gender and species classifications, the ticks were grouped into sets of five and rapidly frozen in liquid nitrogen at −196°C. The frozen ticks were then homogenized in 200 µL of PBS using sterile pestles in a sterile environment. The homogenate was transferred into 1.5 mL conical microtubes for DNA extraction. A 100 µL aliquot of the homogenate was used for DNA extraction with a column-based DNA extraction kit, the Tissue Genomic DNA Extraction Mini Kit (Farvogen, Taiwan, 50 preps), following the manufacturer’s instructions. The extraction procedure was carried out as directed by the manufacturer. The NanoDrop spectrophotometer 2000c (Thermo Scientific, USA) was used to measure absorbance at 260 and 280 nm to evaluate the measure and quality of the extracted DNA from all three samples (blood, tick, and milk). The extracted DNA was then kept at −20°C until PCR was conducted.
Molecular detection of Francisella spp
For the molecular detection of Francisella spp., a nested PCR targeting the16S rRNA gene was utilized (Zargar et al., 2015). The primers presented by Tukmechi et al. (2024) were employed for both the PCR and nested PCR, respectively (Table 1).
The PCR Approach Used to Identify Francisella Species Involves Specific Primers, Product Sizes, and a Thermal Program, Which Are Explained below
During the initial step of the nested-PCR, Taq DNA Polymerase Master Mix RED from Amplicon in Denmark was used. The PCR process was conducted at a 25 μL capacity, with 5 μL of extracted DNA, 2 μL for both the forward and reverse primers, and 12.5 μL of the master mix included. To improve specificity and sensitivity, as well as reduce the probability of contamination and inhibitors in the reaction, the touchdown PCR technique was employed (Don et al., 1991). The touchdown and normal PCR thermal programs were set in the thermal cycler device (Quanta Biotech, England) as described previously (Tukmechi et al., 2024). The PCR experiment was carried out employing a nested PCR method, which operates similarly to conventional PCR yet utilizes different DNA elements. Products of the PCR for both stages on a 2% agarose gel were electrophoresed with 1 μL of safe stain (Labnet, ENDURO™, USA) and visualized using Genius Gel Documentation (Syngene Bio-Imaging, UK) (Figs. 2, 3).
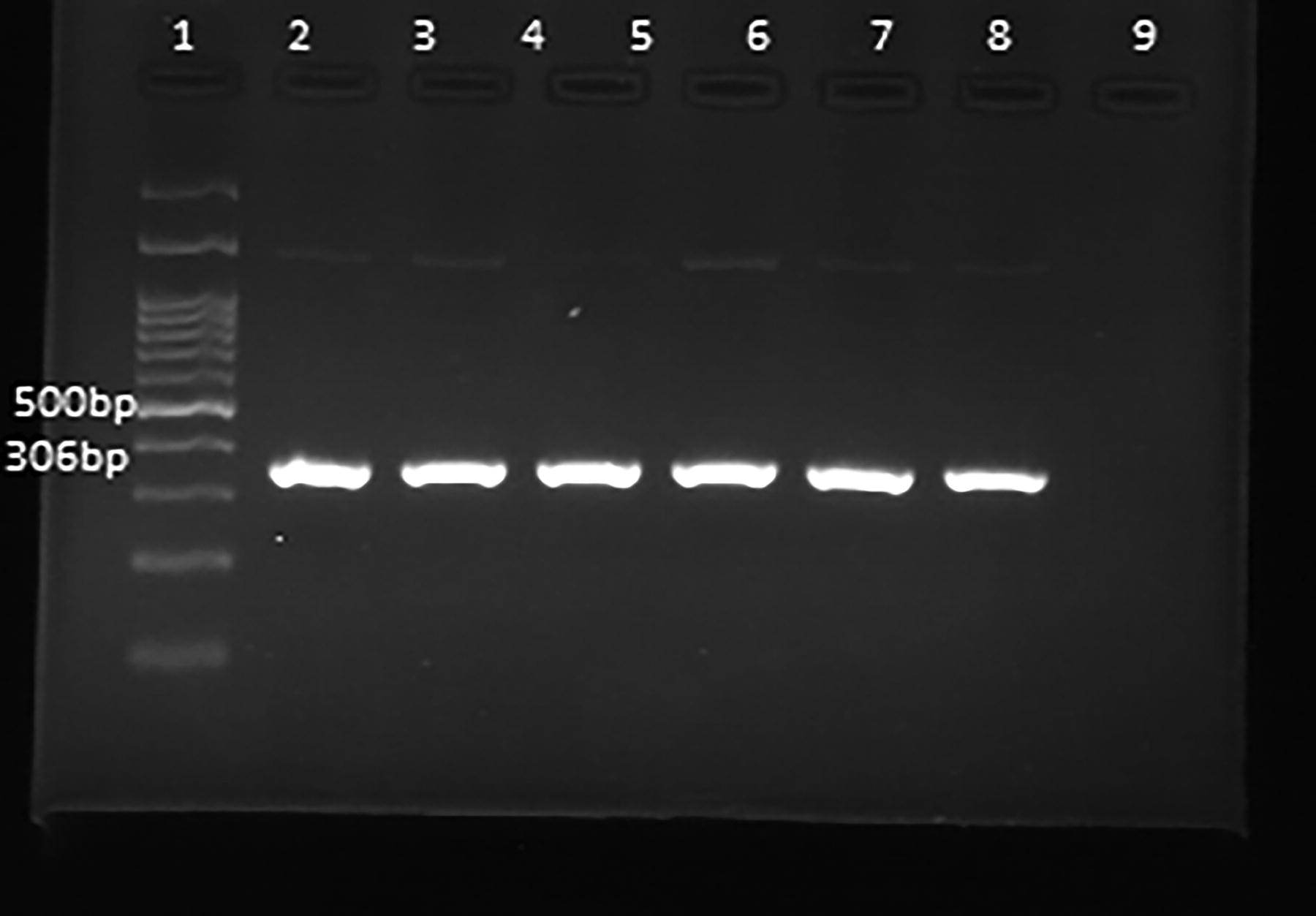
Visualization of amplified Francisella spp. 16SrRNA gene (306 bp) via agarose gel electrophoresis of PCR products from ticks. Lane 1: 100 bp DNA marker (Smobio Technology Inc, Taiwan), Lanes 2,3,4,5,6,7, and 8 positive samples, 9 Lane of negative control.
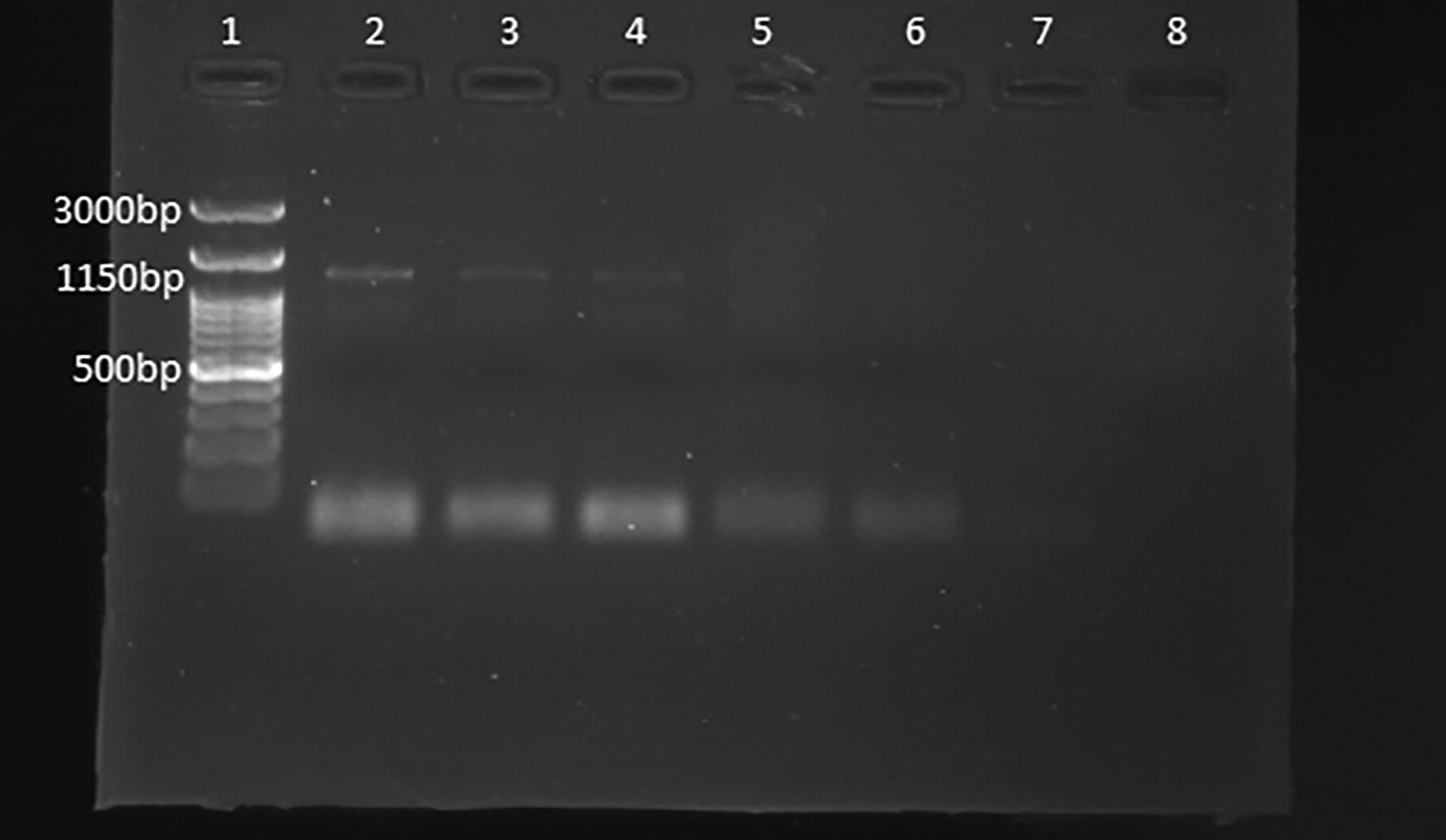
Visualization of amplified Francisella tularensis subspecies holarctica RD1 gene (1150 bp) via agarose gel electrophoresis of PCR products from ticks. Lane 1: 100 bp DNA marker (Smobio Technology Inc, Taiwan), Lanes 2,3 and 4 positive samples and Lanes 5,6, and 7 negative samples and Lane 8 of negative control.
Molecular detection of Francisella spp. subspecies
The presence of RD1 genes was assessed to identify Francisella species subspecies through PCR subtyping assays following WHO guidelines (Alim Aikimbayev et al., 2007). The primers for RD1 genes, as previously described by Rahravani et al. in 2022 (Rahravani et al., 2022), were used for normal-PCR. The PCR protocol for the RD1 gene included an initial denaturation at 95°C for 15 min, and then 36 cycles of 95°C, 57°C, and 72°C for 30 s, 45 s, and 60 s, respectively. Then, 72°C for 10 min (Table 2).
Primer Sequences and Product Size to Recognize F. tularensis Subspecies Using the PCR Procedure
Phylogenetic analysis tree
To confirm the identification of Francisella species, the Sanger sequencing method was used to sequence the RD1 and 16S rRNA amplicons. The Mega (version 11) software was used to conduct phylogenetic analyses. The evolutionary history was inferred using the Neighbor-Joining method. The best-resulting tree is shown. Bootstrap values (1,000 replicates), indicating how often related bacteria grouped, are displayed next to the branches. Sequence divergence was measured using the Tamura 3-parameter method, yielding the number of base changes per site. The first, second, and third codon positions, as well as the noncoding regions, were checked. Unclear parts of the sequences were removed when comparing pairs through pairwise deletion.
Results
Among 317 small ruminants, 76.03% were female and 23.97% were male. In terms of the livestock species, 236 (74.45%) were sheep, and 81 (25.55%) were goats. Blood and milk samples were collected from different counties in the Urmia region, including Noshin (46 blood samples and 30 milk samples), Talatapeh (52 blood samples and 44 milk samples), Jarchlo (64 blood samples and 49 milk samples), Dastjerd (37 blood samples and 26 milk samples), Yaghmorali (50 blood samples and 39 milk samples), and Gharabghlo (68 blood samples and 53 milk samples). No positive cases of Francisella spp. were detected in either the blood or milk samples using the PCR method.
In addition, out of the 317 ticks collected, 56 (17.67%) were from livestock in Noshin, 79 (24.92%) in Talatapeh, 52 (16.40%) in Jarchlo, 23 (7.25%) in Dastjerd, 35 (11.04%) in Yaghmorali, and 72 (22.71%) in Gharabghlo (Table 3). A morphological analysis of the ticks revealed that the most prevalent species were H. asiaticum, accounting for 116 (29.21%), followed by Rhipicephalus turanicus with 97 (24.43%), R. sanguineus with 77 (19.40%), H. anatolicum with 59 (14.86%), and H. aegyptium with 48 (12.09%). All ticks collected were adults, and 54.15% were male. The presence of Francisella spp. was found to be positive in 6 out of 79 tick pools (7.59%; 95% CI: 3.52%–15.59%). The positive ticks identified in this experiment were H. asiaticum (2 out of 24; 8.33%; 95% CI: 2.31%–25.84%), H. aegyptium (3 out of 9; 33.33%; 95% CI: 12.06%–64.58%), and R. sanguineus (1 out of 15; 6.67%; 95% CI: 1.19%–29.82%) (Table 4).
The Number of Ticks Collected from Various Body Areas of Animals Under Study
Different capital letters in each row and small letters in each column showed significant differences (P ≤ 0.05).
Identification of Fransicella SPP. from Ticks Collected from Sheep and Goats
In samples where the 16S rRNA gene was positive, Francisella spp. was confirmed as having products size of 306 bp (Fig. 2). Then, to identify the subspecies of Francisella, PCR was used with the aim of identifying the RD1 gene.
However, the RD1 gene resulted in amplification, having a product size of 1150 bp (Fig. 3). Sequencing of the RD1 gene in tick samples confirmed the presence of holarctica. The presence of F. tularensis subspecies Holarctica was found to be positive in 3 out of 79 tick pools (3.80%; 95% CI: 1.3%–10.58%). The positive ticks identified in this experiment were female H. aegyptium (1 out of 3; 33.33%; 95% CI: 6.15%–79.23%), male H. asiaticum (1 out of 14; 7.14%; 95% CI: 1.27%–31.47%) and female R. sanguineus (1 out of 7; 14.29%; 95% CI: 2.57%–51.32%) (Table 4).
Holarctica subspecies of F. tularensis were identified based on the results of 16S rRNA and RD1 gene sequences and phylogenetic analyses. Their sequences were then added to the Genbank database with the accession numbers PQ637477 and PQ805469, respectively (Figs. 4 and 5). The ticks of H. asiaticum, H. aegyptium, and R. sanguineus that parasitized sheep and goats were shown to harbor the F. tularensis subsp. holarctica.

Francisella spp. phylogeny based on the sequence of 16SrRNA gene. The phylogenetic tree is constructed by MEGA 11 software.

Francisella tularensis subsp. Holarctica phylogeny produced by the neighbor-joining method based on sequence of RD1 gene.
Discussion
F. tularensis is a zoonotic bacterium transmitted by ticks, which is responsible for tularemia in humans. In this study, we assessed the incidence of F. tularensis in the blood and milk samples of small ruminants (sheep and goats) and their ticks in the Urmia region by using a 16S rRNA gene of the Francisella genus via using the nested-PCR technique. These studies raise major concerns about the potential occurrence of tularemia in Iran (Tukmechi et al., 2024).
The detection of F. tularensis in blood samples from livestock is rare, as bacteremia caused by this pathogen is uncommon (Haristoy et al., 2003). The life cycle of F. tularensis may contribute to the difficulty in identifying it in blood samples from livestock (Esmaeili et al., 2014). Our study, consistent with earlier investigations, found F. tularensis not present in goat and sheep blood samples. For instance, a molecular study conducted in the Kurdistan area (Western Iran) using real time-TaqMan PCR similarly showed no positive F. tularensis in animal blood samples even when the infection was identified in tick samples (Rahravani et al., 2022). Correspondingly, research conducted in Iraq (Mohammed et al., 2023) and Egypt (Ghoneim et al., 2017) revealed no indication of F. tularensis in animal blood samples, hence verifying our conclusions.
This analysis did not find any positive samples in milk, even though F. tularensis is intracellular. The use of PCR has enabled the detection of intracellular bacteria, such as Brucella, in milk (Tantillo et al., 2001).
The results of this study revealed the detection of Francisella spp. among examined ticks with prevalence rate of 1.51%. The positive ticks identified in this experiment were H. asiaticum, H. aegyptium, and R. sanguineus, but the pathogen was not find in H. anatolicum and R. turanicus. The incidence of Francisella spp. in ticks indicated 1.7% in the Kurdistan region in Iraq, Iran’s neighbor, which was a little more than our result (1.51%) (Mohammed et al., 2023). The results of our study from research conducted using the real-time TaqMan PCR method in western Iran are slightly higher (0.82%) (Rahravani et al., 2022), which may indicate that we are closer to the source of contamination. In a separate genetic study in Egypt, 4.7% of ticks collected from camels tested positive for Francisella spp., consistent with our results showing the absence of these bacteria in camel blood and fecal samples. The investigation also disclosed a notable presence of F. tularensis antibodies in slaughterhouse workers, with those frequently exposed to tick bites exhibiting a higher seroprevalence (20.7%) compared to those with infrequent exposure (2.2%) (Ghoneim et al., 2017). Conversely, a Malaysian study presented unusual findings, indicating a limited distribution of the bacteria with no detection of Francisella spp. in ticks or animal samples from livestock farms. Transmission to other ticks or animals was not observed, with only Dermacentor questing ticks identified as carriers. Specifically, ticks attached to cattle, sheep, or goats, as well as animal blood samples, showed no signs of the bacterium, while 11.3% of questing ticks from forest reserves tested positive for the 16S rRNA of Francisella spp. (Koh et al., 2019). Recently, in Iran’s West Azerbaijan region, a molecular investigation identified F. tularensis in tortoise blood using the Nested-PCR method with a specific 16S rRNA gene linked to the Francisella genus. By conducting morphological examinations, the ticks were recognized as H. aegyptium. The findings revealed a presence of Francisella in 7% of animal blood samples, with a detection rate of 15% in tick samples (Tukmechi et al., 2024).
Ticks play a vital role in transmitting important pathogens to humans and animals, making them indicators of infections in the natural environment (Rizzoli et al., 2011) . Various tick species, which are diverse and adaptable, have recently expanded their geographic range and habitats. This expansion is greatly influenced by changes in land use, shifts in climate conditions, and global interdependence (Harrus and Baneth, 2005).
In this study, a greater number of male ticks were captured compared to females. However, among the 79 pools of ticks examined, four female ticks tested positive for Francisella spp., while only two pools of male ticks tested positive. Female ticks, due to their higher likelihood of feeding on various hosts throughout their lifespan and their increased reproductive capacity, may have more interactions with diseases. In addition, the stress of egg production could weaken their immune system, making them more vulnerable to pathogens (Wang et al., 2020). The significant presence of intracellular bacteria in female ticks suggests they may play a role in the transmission of tick-borne diseases, as these bacteria can be passed on to their offspring (Beninati et al., 2004; Lo et al., 2006).
The population of organisms rises as ticks progress from the larval to adult phases of their life cycle, with Francisella spp. being detectable in the gut and hemolymph of ticks (Petersen et al., 2009). While some research indicates that ticks could potentially serve as a reservoir for these bacteria, further investigation is needed to confirm this. It has been established that Francisella spp. can be transmitted between different life stages of ticks, but the transmission through eggs is still uncertain. Numerous studies support the role of ticks as biological vectors in transmitting Francisella spp. among mammalian hosts and maintaining these bacteria within populations during periods between outbreaks (Genchi et al., 2015). Based on extensive evidence, ticks are known as biological carriers that spread Francisella spp. among mammalian hosts, functioning as reservoirs during intervals between outbreaks (Gehringer et al., 2013). With potential habitat expansions for ticks due to climate change, the risk of increased tularemia cases may escalate, emphasizing the need for ongoing surveillance (Schutz et al., 2024).
The increased global processing and distribution of food has raised awareness regarding food safety (Lamont et al., 2014). The Centers for Disease Control and Prevention (CDC) classified F. tularensis subsp. tularensis as a class A agent due to its high infection rate, ease of dissemination, and its potential pathogenicity (Willke et al., 2009). In certain geographical regions, tularemia transmission has been linked to interaction with infected animals and the consumption of contaminated water or food, as well as transmission through arthropods (Duzlu et al., 2016). Additionally, certain occupations, including laboratory personnel, farmers, veterinarians, hunters, cooks, and individuals who handle meat, are also associated with increased risk (Nigrovic and Wingerter, 2008). There have been limited studies focused on recognizing F. tularensis in food and environmental samples, even considering the public health concerns (Lamont et al., 2014). It is believed that tularemia outbreaks in Turkey could have been spread through water rather than vectors (Duzlu et al., 2016).
Conclusion
The purpose of this research was to identify any Francisella spp. in small ruminant blood, milk, and ticks in the Urmia region in Iran. We used nested PCR to examine blood and milk samples from goats and sheep, and we failed to detect any Francisella spp. However, the bacteria were found in tick populations, specifically in R. sanguineus, H. aegyptium, and H. asiaticum. These results indicate that ticks may serve as potential vectors for Francisella spp. in this region, whereas the blood and milk of small ruminants do not appear to pose a substantial risk as sources of the pathogen. This study was almost a detection examination and did not examine the transmission dynamics of Francisella spp. among ticks, livestock, and people. Additional study is necessary to clarify the function of ticks in the transmission cycle of this infectious agent and to evaluate the likelihood of infection in both animal and human populations within the region. To decrease the risk of exposure to this zoonotic pathogen, public health authorities ought to think about the implementation of tick control programs and the promotion of awareness among individuals employed in animal-related professions.
Footnotes
Author Disclosure Statement
No competing financial interests exist.
Funding Information
Each author confirms that their research is supported by an institution that is primarily involved in education or research.