
Abstract
Rodents are associated with the spread of deadly zoonotic and emerging infectious diseases. Their widespread distribution is strongly linked to their ability to thrive in human-altered environments with access to food and shelter. Thus, humans are exposed to zoonotic agents that can spread and infect them directly and indirectly. We sourced published literature on ectoparasites associated with Rattus rattus, R. norvegicus, and Mus musculus from various academic databases such as Google Scholar, PubMed, Web of Science, and Scopus. The relevant literature was screened and selected in R statistical software using the packages ‘‘metagear’’ and ‘‘revtool.’’ We reviewed and analyzed a total of 53 articles published between 1994 and 2024. The literature review covered studies from 29 countries, represented by six continents, namely, Africa, Asia, Europe, North America, Oceania, and South America, with the majority of the studies from Asia. R. rattus had the most published studies. Our literature review identified 87 species of zoonotic ectoparasites, with Xenopsylla cheopis, Polyplax spinulosa, and Laelaps echidninus being the most reported across all host species. Mites were the most reported ectoparasites (40.83%), followed by fleas (25%), ticks (20%), and lice (14.17%). Most of these ectoparasites were reported from R. rattus, with the highest number of documented ectoparasites. Investigating the prevalence, distribution, and potential risks of alien invasive rodents and their ectoparasites is paramount for public health and improving integrated pest management control strategies.
Introduction
Invasive murid rodent species have expanded beyond their native ranges over time (Dahmana et al., 2020; Measey et al., 2020; Witmer and Shiels, 2017). Their global distribution has been facilitated by a commensal relationship with humans, particularly through international trade and human migration patterns (Banks and Smith, 2015; Lucaccioni et al., 2016). Alien invasive rodents, including Norway or brown rat (Rattus norvegicus); the black, roof, or house rat (R. rattus); and the house mice (Mus musculus) have been associated with the spread of deadly zoonotic diseases (Akhtar et al., 2023; Reis Da Silva Fernandes et al., 2021) and emerging infectious diseases (Badou et al., 2024; Lucaccioni et al., 2016; Zhang et al., 2022). Zoonotic diseases, defined as infections transmissible between animals and humans (Balakirev and Van Chau, 2021; Ferreira et al., 2021; Franjić, 2022; Sanyaolu et al., 2016), have been extensively studied in the context of rodent–parasite interactions (Islam et al., 2021a; Ng et al., 2017; Singla et al., 2008). Parasitism represents a complex symbiotic relationship where parasites evolve dependence on hosts for survival and/or reproduction (Betts et al., 2018; Gómez-Arreaza et al., 2017; Zuzarte-Luís and Mota, 2018). The parasite–host relationship focuses on more than the damage a parasite causes to its host; it also involves frequent interactions at the population level and the dynamic process of the parasite moving from one host to another (Buck et al., 2018; Papkou et al., 2016; Tompkins et al., 2002). Substantial evidence indicates that parasites significantly impact host fitness in the wild and account for a selective force in community structure (Hudson et al., 2002; Papkou et al., 2016; Tao et al., 2016).
Parasites can manipulate the host, changing it in ways that create a more suitable habitat for other parasite species and ease the transmission of the parasite to its next host (Binning et al., 2017; Hafer, 2016; Mabbott, 2018; Poulin and de Angeli Dutra, 2021). While some parasites successfully establish in new environments, most fail to persist in invaded ecosystems (Gehman et al., 2017; Gunn and Pitt, 2022; Miura et al., 2006). Humans living in close proximity to rodents face significant risks from zoonotic agents through multiple pathways, which include direct contact through contaminated food or water, rat bites, or urine or indirectly through other vectors (Esposito et al., 2023; Hamidi and Bueno-Marí, 2021; Menona and Jolly, 2020). An example of a zoonotic disease that an infected rat can transmit directly through biting is leptospirosis, with approximately one million cases and 58,000 deaths worldwide (Mohd Zain et al., 2015; Murray et al., 2020; Sykes et al., 2022). Rodents serve as reservoirs and immediate hosts for emerging pathogens (Battersby, 2015; Davoust and Laidoudi, 2023; Devanathan et al., 2024) and host various arachnids species, such as mites, fleas, ticks, lice, and chiggers, collectively known as ectoparasites (Benedek et al., 2024; Hamidi and Bueno-Marí, 2021; Obiegala et al., 2021). Ectoparasites are organisms that inhabit the external surface of a host (Hinkle and Corrigan, 2020; Islam et al., 2021a; Shiferaw, 2018). A single rodent can carry more than one ectoparasite species (López-Pérez et al., 2022; Salas et al., 2019; Shilereyo et al., 2022). The Oriental rat flea (Xenopsylla cheopis) is an example of an ectoparasite flea species responsible for the transmission of Yersinia pestis, the causative agent of plague, through indirect transmission (Cross et al., 2021; Mohd Zain et al., 2015).
Significance of the Study
Understanding rodent ecology and associated diseases is important for mitigating public health risks (Han et al., 2016; Menona and Jolly, 2020; Murray et al., 2020). This enhances identifying biological factors that can predict the high success rates in colonizing and inhibiting various environments (Gardner‐Santana et al., 2009; Puckett et al., 2016; Russell et al., 2019). Human activities, including deforestation, urbanization, agriculture, domestication of livestock, and climate change, put the ecosystem in continuous turmoil with the changes in the assemblage and distribution of parasites and their vectors (Esposito et al., 2023; Hamidi and Bueno-Marí, 2021; McMichael, 2004). These land-use changes represent primary drivers of emerging infectious diseases and rodent infestation as they share dwellings with humans (Castillo et al., 2003; Ecke et al., 2022; Patz et al., 2008). The three synanthropic species, R. norvegicus, R. rattus, and M. musculus, are considered commensal rodents due to their close association with humans (Allam et al., 2019; Battersby, 2015; Islam et al., 2021b; Puckett et al., 2020). These murid rodents are prevalent in urban areas, inhabiting buildings, destroying property, and endangering the health of occupants by spreading diseases (Chelule and Mbentse, 2021; Julius et al., 2021; Sodikoff, 2019). Several studies have shown that areas that lack adequate sanitation, public services, and building maintenance have the highest population of rats and mice (Julius et al., 2021; Langton et al., 2001; Rafique et al., 2021). As the public hygiene standards decline, the rodent population increases (Iles et al., 2006; Kunene, 2016; Rafique et al., 2021). Their populations grow rapidly because of their high reproductive capacity to utilize available habitats as shelter and breeding grounds, advantageously self-regulating their populations in stable environments (French et al., 2021; Russell et al., 2019; Tobin and Fall, 2004). The most important environmental component for any organism is its habitat (Bastille‐Rousseau and Wittemyer, 2021; Boyce et al., 2016). Various control methods have been implemented to target specific damage, focusing on environmental controls to manage rodent populations at economically and socially acceptable levels (Nembudani, 2017; Parsons et al., 2017; Tobin and Fall, 2004).
Aim of the Study
This study presents a systematic literature review examining commensal rodents and their role in disease ecology. Through comprehensive analysis of existing research, we aimed to (a) assess the global distribution of ectoparasites carried by these rodents, (b) estimate the prevalence of ectoparasites among commensal rodents, (c) identify rodent species harboring the highest diversity of ectoparasites, (d) determine common ectoparasite species found across various rodent populations, and (e) to highlight research gaps that call for additional research for ectoparasites associated with zoonotic diseases in murid rodents. This study’s findings will help identify key research gaps and guide future strategies for managing the risks associated with ectoparasites in invasive alien murid rodents in South Africa.
Materials and Methods
Literature search
We sourced published literature from multiple academic databases to gather information on the global zoonotic ectoparasites associated with invasive rodent species. The databases used included (i) Google Scholar (https://scholar.google.com/), (ii) PubMed (https://pubmed.ncbi.nlm.nih.gov/advanced), (iii) Web of Science (https://www.webofscience.com/wos/), and (iv) Scopus (https://www-scopus-com-s.web.bisu.edu.cn). To widen the search of the published literature across multiple platforms, including PubMed, Web of Science, and Scopus, the following keywords were used to determine zoonotic ectoparasites associated with the three murid rodents: “zoonotic ectoparasites,” “Rattus rattus,” “Rattus norvegicus,” and “Mus musculus.” For Google Scholar, we used the following phrases: “zoonotic ectoparasites associated with Rattus rattus,” “zoonotic ectoparasites associated with Rattus norvegicus,” and “zoonotic ectoparasites associated with Mus musculus.” We also used SciSpace (https://typeset.io/) to search for the additional relevant literature we could have missed. The research articles were selected according to the relevance of the current study and exported as RIS, TXT, and BIB files readable in R statistical software version 4.3.3 (R Core Team, 2022). The flowchart depicting the process of conducting the Boolean search following the Preferred Reporting Items for Systematic Reviews and Meta-Analyses (PRISMA) (Moher et al., 2009) guidelines was created using the yEd Graph Editor application (version 3.23.2; yEd Graph Editor, 2023).
Literature inclusion and exclusion criteria
The exported data were screened in R statistical software using packages ‘‘metagear’’ and ‘‘revtool’’ (Lajeunesse, 2016; Westgate, 2019). The data extracted from the academic databases were merged into one file. Relevant articles were identified by reviewing their titles and abstracts, focusing on those containing similar keywords to the current study. Duplicate articles were excluded to ensure data quality and reliability before analyzing individual abstracts.
Data extraction
We filtered studies that met our criteria by analyzing the full texts of all the included articles. The review encompasses articles that employ diverse methodologies for the detection of zoonotic pathogens, including both indirect (e.g., serology) and direct (e.g., isolation) approaches. The data extracted included the identity of ectoparasite species, the rodent species associated with these ectoparasites, sample sizes, the prevalence of rodents infested with ectoparasites, publication years, countries of study, and article titles. The geographical regions of study locations were analyzed using R statistical software version 4.3.3 (R Core Team, 2022).
Results
PRISMA study selection
We initially identified a total of 450 articles through academic literature databases (Fig. 1). After removing duplicates, 411 articles underwent screening. We then excluded 283 articles after evaluating titles and abstracts as they were outside the scope of the current study, and 73 articles were further removed for focusing on rodent-borne zoonosis without including ectoparasites of the alien invasive host species we were targeting. Most of the studies were excluded because they focused on host species carrying nonzoonotic ectoparasites. Thus, a total of 53 articles were reviewed to fulfil the criteria of the synthesis (Fig. 1).
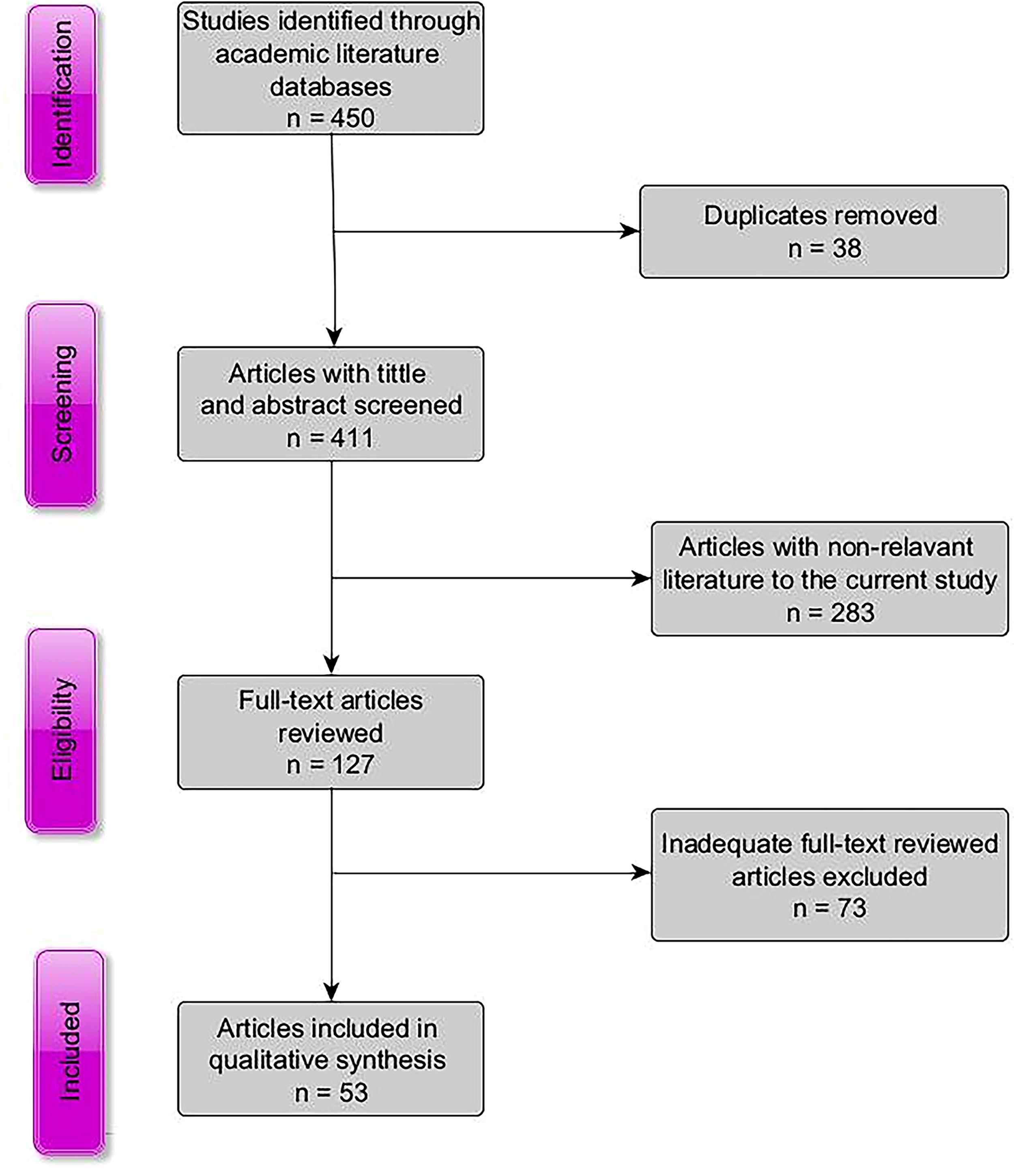
PRISMA flow diagram of the literature review screening process of articles included in the study add the reference of the PRISMA guidelines. PRISMA, Preferred Reporting Items for Systematic Reviews and Meta-Analyses.
Number of identified studies
Among the 54 identified articles, publication years ranged between 1994 and 2024 based on studies conducted on zoonotic ectoparasites associated with commensal invasive murid rodents (Fig. 2). Most studies were published between 2015 and 2021, followed by 2009 and 2023. The 19th century had the least number of publications compared to the 20th century. The first study that met the criteria was published in 1994 in Angola, reporting data on some ectoparasites of commensal rodents from Huambo. No studies were identified between 1998 and 2004 and 2011 (Fig. 2). R. rattus had the most published studies with M. musculus having the least published studies over the years.
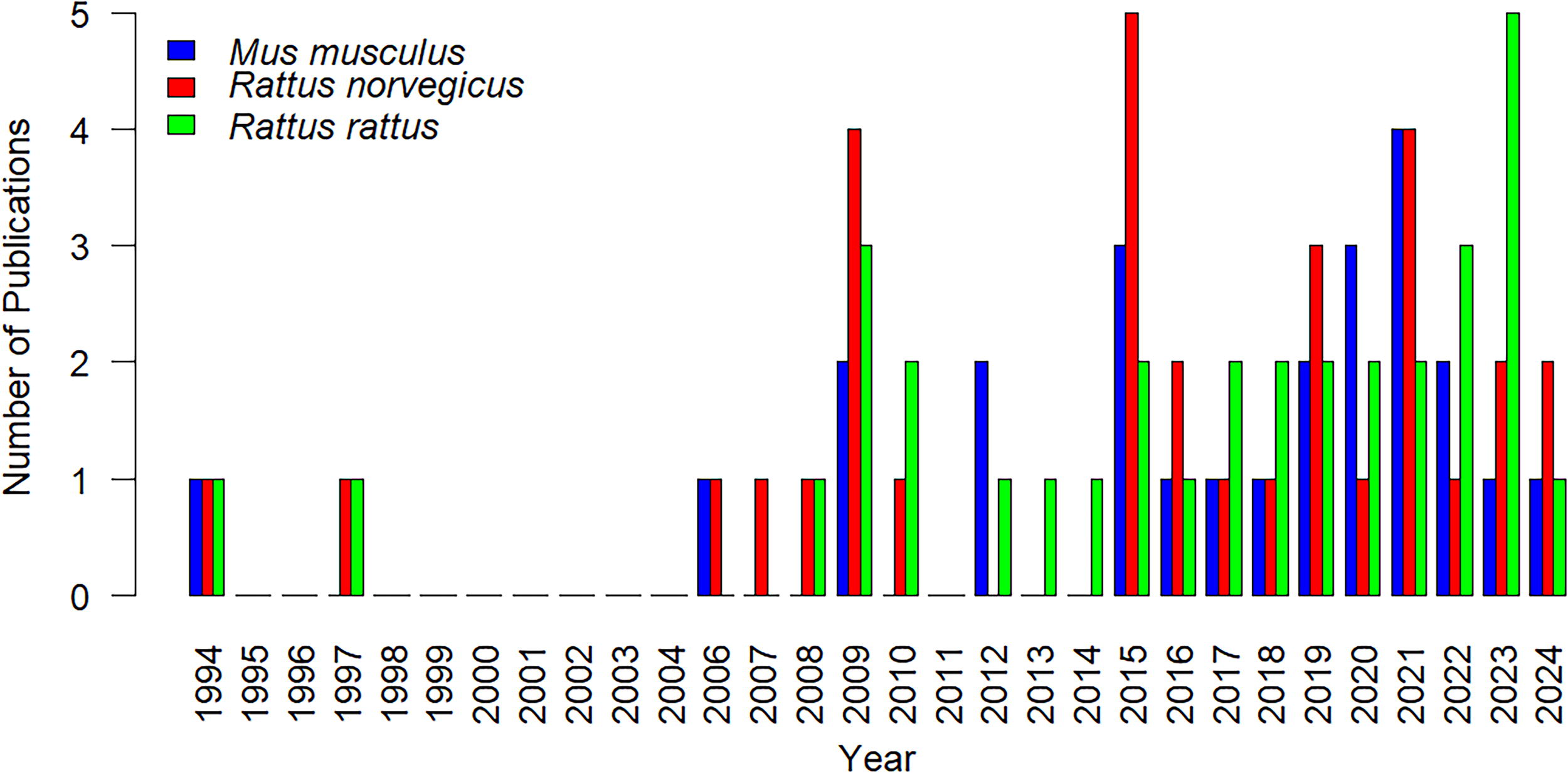
The number of publications from 1994 to 2024 (a 30-year period) associated with zoonotic ectoparasites of the three commensal alien invasive rodents.
Geographical locations
The literature review covered studies from 29 countries (Fig. 3), with Iran (n = 11, 21.3%) having the most published articles, followed by the United States (n = 5, 9.2%). The overall studies took place in six continents, with the majority undertaken in Asia (n = 29, 53.7%) followed by Africa (n = 13, 24%), North America (n = 6, 11.1%), Europe (n = 3, 5.5%), and South America (n = 2, 3.7%). Oceania was the only continent where a single study was documented, with Australia as the sole country involved. This suggests a research gap in the continent regarding rodent-borne diseases. Iran had the greatest number of ectoparasite species (n = 30, 32.6%). Lithuania, Mali, and Vietnam are countries with the least ectoparasite species, their conducted studies reported one pathogen each (n = 1, 1.08%).
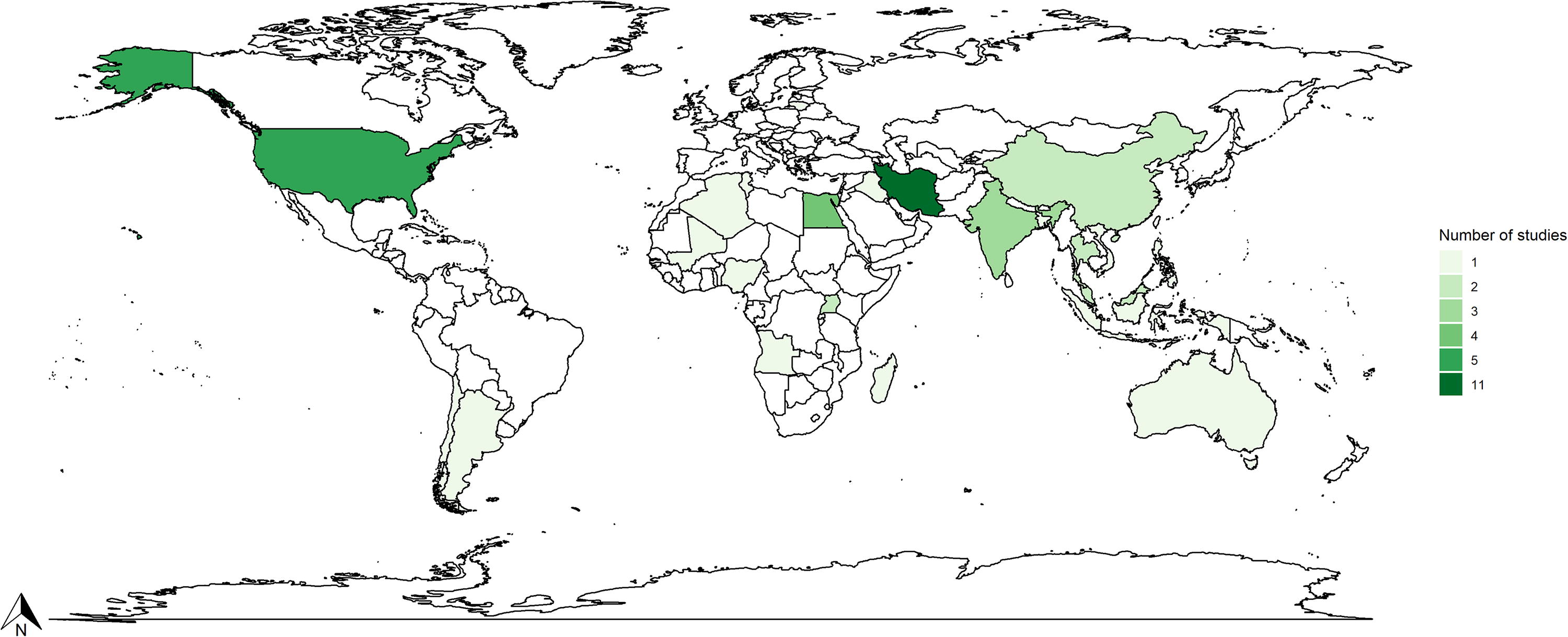
Map displaying the geographical distribution of the number of studies conducted on ectoparasites of the three commensal rodents between 1994 and 2024. (The colors represent countries, with the number of studies listed beside them).
Zoonotic ectoparasites
Each rodent displayed varied patterns of occurrences of parasites. R. rattus had the highest mite species (n = 21), with both R. norvegicus and M. musculus having the same mite species (n = 14) (Fig. 4). R. rattus had the highest flea species (n = 13) followed by M. musculus, having the lowest flea species (n = 7). Lice species were less prevalent compared to the other parasites, with seven lice species reported on R. norvegicus, followed by R. rattus (n = 5), and M. musculus (n = 5) (Fig. 4). Both Rattus species had the same number of tick species (n = 10) with the least number of tick species (n = 4) recorded on M. musculus (Fig. 4).

Sankey diagram showing the occurrence of the recorded ectoparasite pathogens among each rodent species.
Distribution of zoonotic ectoparasites among host species
The number of ectoparasite species found exclusively in each host species was 24 for R. rattus, 13 for R. norvegicus, and 8 for M. musculus, respectively (Fig. 5). Ectoparasites found in both R. rattus and R. norvegicus were 16 (Fig. 5). Those that occurred in both R. rattus and M. musculus were only three, while those found in both R. norvegicus and M. musculus were 8. Fifteen ectoparasite species were reported in all host species (Fig. 5).
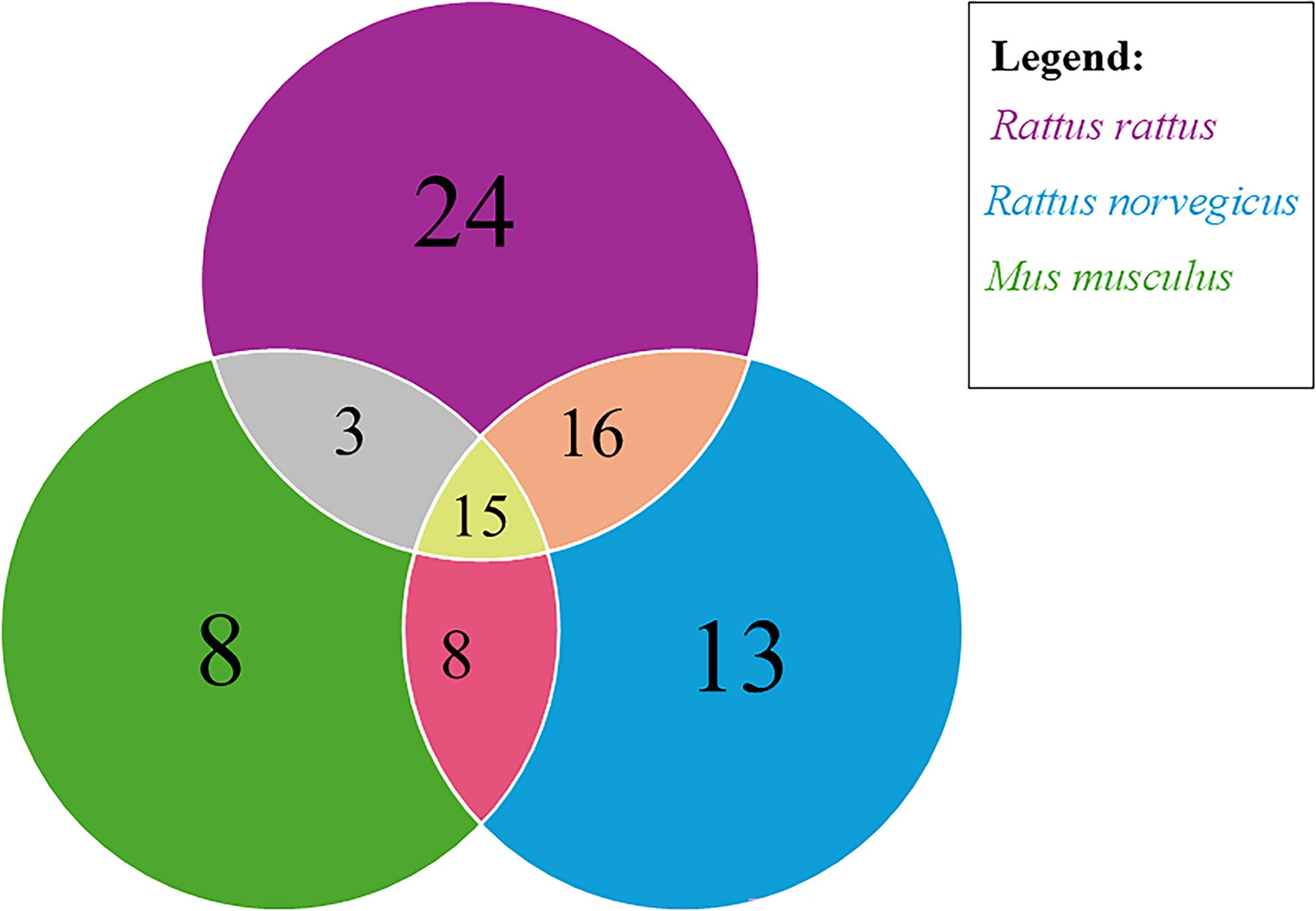
Venn diagram representing the distribution of zoonotic ectoparasites among the three invasive murid rodent species.
Fleas, lice, mites, and ticks carried by rodents
Our literature search found a total of 248 examined ectoparasites associated with the invasive murid rodents (Table 1). A total of 92 flea species from 18 species were examined, based on records from 53 articles (Table 1). X. cheopis was the most frequently reported species of flea, with 40 records. Four flea species were reported in all three host species, and this included Ctenocephalides felis, Leptopsylla segnis, Nosopsyllus fasciatus, and X. cheopis (Table 1). The included articles documented a total of 42 lice from 10 different lice species on rodents Polyplax spinulosa was the most documented lice species with 21 records, while Laelaps echinus was the most recorded mite species, with 14 records of the total 81 mites documented (Table 1). The reviewed articles documented 15 tick species from which Rhipicephalus sanguineus (seven records) was the highest.
Zoonotic Ectoparasites Associated with Alien Invasive Rodents
Represents families of ectoparasites, which were not identified to the species level.
The numbers represent the number of times ectoparasites were recorded in different articles per host species.
Discussion
Our study reviewed published literature on rodent ectoparasites to provide a global overview of zoonotic ectoparasites associated with R. rattus, R. norvegicus, and M. musculus. The review showed that most of the studies (11 of 53) were from Iran, likely due to historical rodent-borne epidemics such as the plague that was accountable for numerous deaths, wherein rodents were reported as natural reservoirs for carrying the disease (Azizi and Azizi, 2010; Khosravani, 2018; Shahraki et al., 2016). Research output increased substantially in the 20th century relative to the 19th century. This could be explained by the sudden interest in studying rodent-related pathogens can be explained by an increase in human-transmitted diseases relating to rodents and their ectoparasites and factors influencing the transmission of pathogens (Canavan, 2023; Dahmana et al., 2020; Islam et al., 2021a).
Although some countries had fewer studies or none reported in our systematic review, zoonotic ectoparasites have been identified in rodent species from these countries that were not investigated in the review. The review encompasses data from six continents, with most studies published in Asia and Africa. This may be explained by that these continents are experiencing outbreaks related to increased public health risks and infections (Allen et al., 2017; Blasdell et al., 2022). For example, in the last decade, leptospirosis has had numerous outbreaks and has been reported as a major emerging and reemerging disease among humans and animals, causing over 7000 reported fatal cases (Ganjeer et al., 2021). Oceania had only one published study in the present review. This implies a significant research gap in the country regarding rodent-borne diseases. A study by Brown and Henry (2022) investigated the available knowledge on the impacts of mice on sustainable fodder storage in Australia. They found that while growers were concerned about disease risk, they lacked knowledge about specific parasites or their impacts on livestock or humans. Hence, research focuses may differ among countries.
The current review indicated a notable proportion of studies were conducted between 2015 and 2021. A possible explanation for the high number of studies in recent years could be the rise in reports of zoonotic disease transmissions influenced greatly by climate change. Climate change affects the transmission of zoonotic diseases by arthropod vectors since they are ectothermic, such as ticks, lice, fleas, and mites (Morley and Lewis, 2014; Ogden, 2017). Over the years, there have been cases of a rise in global temperatures and expectations to directly influence the abundance, distribution, habitat suitability, and activity patterns of vector-borne zoonotic diseases risk (Banda et al., 2023; Rupasinghe et al., 2022). Another possible explanation could be the improvement of technology and accessibility of research tools, which are more advanced and reliable references.
Moreover, knowledge of clinical symptoms and finding ways to prevent and control the spread of diseases increases the number of publications (Adhikari et al., 2020). Our review found that R. rattus is the most common and frequently studied alien invasive rodent species. This may be due to its ability to colonize and adapt to a newly invaded habitat and being among the most notorious alien invasive species worldwide (Diagne et al., 2017; Hancke and Suárez, 2022; Hima et al., 2019; Konečný et al., 2013). However, R. rattus should not be singled out, as murid rodents of the genus Rattus are a common example of an invasive group of small mammals that are widespread and ecologically adaptable (Dammhahn et al., 2017). Their remarkable adaptability makes them of the highest importance for carrying zoonotic pathogens, consequently resulting in emerging infectious diseases (Diagne et al., 2017; Islam et al., 2021a; Roy et al., 2023).
The present literature search found that fleas were the most studied ectoparasites, whereby X. cheopis was the most frequent flea species reported. The X. cheopis is one of the species involved in the transmission of plague. For example, the plague is native to the highlands of Madagascar, where it has been found that X. cheopis is one of the two flea species primarily responsible for transmitting the disease (Kreppel et al., 2016). Furthermore, R. norvegicus harbored more flea species than R. rattus and M. musculus, due to the persistence of fleas in the wild and laboratory (Fitzgerald et al., 2023).
Conclusion
The data presented in our study reveal research gaps on commensal invasive murid rodents in several countries, indicating a need to conduct more studies on rodent-borne zoonosis and their public health importance. Although some studies were overlooked based on not being host-specific, the results show that further research can provide more insight into the relationship between the prevalence of host species and emerging zoonotic infections. Thus, it is important to investigate and identify the potential impacts that invasive rodents may pose and the risks available. Prevention is the best option for managing the overwhelming negative impacts caused by alien invasive species, which will require more relevant and accurate knowledge as the threats for epidemics increase, and it is almost impossible to predict when and where they will take place. These recommendations should be considered during rodent ectoparasite control programs.
Footnotes
Acknowledgments
The authors sincerely thank the Department of Nature Conservation and the Directorate of Research and Innovation at Tshwane University of Technology (ZA) for their support. Their contributions were instrumental in the successful completion of this research. The authors also sincerely thank the anonymous reviewers for their insightful comments and constructive feedback, which significantly strengthened this study.
Author Disclosure Statement
The authors have no conflicts of interest to declare.
Funding Information
This work was supported by funding from Stud Game Breeders.