
Abstract
Toll-like receptors (TLRs) recognize an increasingly broad range of pathogens, thus demonstrating the importance of these pattern-recognition receptors (PRRs) in host defense. Here, the role of TLR3 in the interaction of monocyte-derived dendritic cells (moDCs) with human cytomegalovirus (HCMV) was investigated by using the TB40E strain, which actively replicates in moDCs. Microarray analysis and quantitative real-time PCR revealed that TB40E infection of moDCs led to changes in the gene expression pattern. A variety of proinflammatory cytokines and chemokines (CXCL10, CXCL11, and CCL5), TLR3, and genes whose products function downstream of the TLR3 signaling pathway (e.g., IFN-α and IFN-β) were significantly upregulated. By silencing TLR3 expression with short interfering RNA (siRNA), and subsequent stimulation with TLR3 ligand poly I:C, expression of IFN-β was markedly reduced compared to cells transfected with a non-silencing control siRNA. However, expression of IFN-β induced by HCMV was not diminished when TLR3 was silenced first. Thus the early HCMV-triggered immune response of human moDCs appears to be independent of TLR3 signaling.
Introduction
Attachment of HCMV to the cellular plasma membrane is undetected by the host, and leads to cellular activation and expression of proinflammatory genes (15). Compton et al. identified toll-like receptor 2 (TLR2) as a central receptor for recognition of HCMV virions (4).
Toll-like receptors (TLRs) are evolutionarily-conserved receptors that are homologues of the Drosophila toll gene. TLRs play a crucial role in the early innate immune response by detecting highly-conserved pathogen-associated molecular patterns comprising viral structures (1,28). For example, TLR3 is a known receptor for poly I:C, a double-stranded RNA (dsRNA) homologue, which triggers a MyD88-independent pathway associated with expression of interferon-β (IFN-β) (26).
Tabeta et al., who investigated the interaction between TLRs and murine cytomegalovirus, found that the TLR3 signaling pathway is activated in vivo upon viral inoculation, leading to the production of class I interferons as an essential part of the innate immune defense against systemic viral infection (38).
In human monocyte-derived dendritic cells (moDCs), TLR3 is localized in intracellular endosomal compartments, thereby detecting pathogens after penetration. DCs are the sentries of the immune system, and are involved in the initiation of both the innate and the adaptive immune responses. In their immature state they rest in peripheral tissues, but undergo maturation upon direct contact with pathogens, resulting in improved antigen presentation, expression of co-stimulatory molecules, and subsequent stimulation of naive T cells in lymphoid organs by secreting various cytokines (3).
In this study, gene expression profiling was used to characterize subsets of genes induced by HCMV in human moDCs, and to identify whether the immune response is mediated by TLR3. We used the endothelial cell–propagated HCMV strain TB40E, which actively replicates in moDCs (20). TLR3 expression was silenced with TLR3-specific short interfering RNA (siRNA) to further examine the role of the TLR3 signaling pathway in moDCs.
Materials and Methods
Cells
Peripheral blood mononuclear cells (PBMCs) were isolated from 50-mL of buffy coat blood (Blutspendedienst Tübingen, Frankfurt, Germany) by Ficoll-Hypaque density gradient centrifugation.
CD14+ cells were isolated from PBMCs by magnetic-associated cell sorting using paramagnetic microbeads conjugated to monoclonal anti-human CD14 antibodies (Miltenyi, Bergisch Gladbach, Germany). Monocytes were seeded onto 6-well plates (2.5 × 106 cells in 3 mL/well) in RPMI-1640 (plus GlutaMAX) medium containing 10% fetal calf serum (FCS) and 100 mg/mL gentamicin sulfate (Refobacin®; Merck, Darmstadt, Germany), supplemented with 100 ng/mL of recombinant human granulocyte-macrophage colony-stimulating factor (GM-CSF ) (Leukomax; Novartis, Basel, Switzerland) and 20 ng/mL interleukin-4 (IL-4) (R&D Systems, Wiesbaden, Germany). The cultures were fed with fresh medium and cytokines every other day. DC differentiation was monitored by light microscopy, and purity of the moDC culture was analyzed after 5 days by flow cytometry (FACSCalibur; Becton Dickinson, Heidelberg, Germany).
At least 95% of the moDC population stained was positive for CD1a, CD40, CD80, and CD86, and was negative for CD14 and CD83 (data not shown).
Virus preparation and infection of moDCs
Human foreskin fibroblasts (HFFs) were isolated from the foreskins of male neonates by trypsin treatment and were used for experiments between passages 10 and 25. Alternatively, MRC-5 fibroblasts were purchased from Sigma-Aldrich, Taufkirchen, Germany.
The fibroblasts were cultured in MEM containing glutamine (2.4 mmol/L), gentamicin (100 μg/mL), and 5% fetal calf serum. For high-titer virus preparations, the fibroblasts were infected at a low multiplicity of infection (MOI) with TB40E. Six days after infection, 100 mL of cell-free supernatant was concentrated by ultracentrifugation at 80,000 × g for 70 min, and pelleted virus particles were resuspended in appropriate volumes of RPMI.
Then 1 × 106 moDCs were incubated with the virus preparation at 37°C (MOI = 50) in complete RPMI medium supplemented with IL-4 and GM-CSF. After 2 h, the cells were washed once with HBSS and resolved in fresh medium. For mock-infection, cells were exposed to fresh medium as an appropriate control for cell-free virus concentrates.
TB40E-infected or mock-infected moDCs were collected after 24 h to assess the efficiency of infection. The cells were washed in PBS plus 2% FCS and 2 mmol EDTA, cytocentrifuged for 5 min at 550 rpm, and fixed on glass slides with acetone for 10 min. HCMV immediate early (IE) antigen expression was detected by indirect immunofluorescence staining. The slides were incubated with monoclonal antibody directed against the non-structural proteins IE72 and IE86 (pUL122/123; Biosoft, Paris, France) at the appropriate dilution for 90 min at 37°C, then were incubated with Cy3-conjugated Fab'2 goat anti-mouse IgG (Jackson ImmunoResearch Laboratories, West Grove, PA) for 45 min at 37°C. Nuclei were counterstained with DAPI. The slides were washed in PBS for 5 min after each incubation step. The percentage of infected cells was determined by immunofluorescence microscopy. As previously reported by Grigoleit et al., IE antigen was detected in 35–75% of the moDCs infected with TB40E (11).
Isolation of RNA
RNA was isolated and purified using the RNeasy Minikit and QiaShredder spin columns (Qiagen, Hilden, Germany) according to the manufacturer's protocol; 35 μL of eluate was obtained and stored at −80°C for further use. The concentration of total RNA was quantified with a Nanodrop spectrophotometer (Peqlab, Erlangen, Germany).
Microarray analysis
Double-stranded cDNA was synthesized from 5 μg total RNA by using a Superscript Choice kit (Invitrogen, Karlsruhe, Germany) with a T7-(dT)24 primer incorporating a T7 RNA polymerase promoter (Metabion, Martinsreid, Germany). cRNA was prepared and biotin-labeled by in-vitro transcription (Enzo Biochemical, New York, NY). Labeled RNA was fragmented by incubation at 94°C for 35 min in the presence of 40 mM Tris-OAc (pH 8.1), 100 mM KOAc, and 30 mM MgOAc. Labeled, fragmented cRNA (15 μg) was incubated for 16 h at 45°C with a HG-U133A human genome array (Affymetrix, Santa Clara, CA). The gene chips were then automatically washed and stained with streptavidin-phycoerythrin on a fluidics station. The probe arrays were scanned at 3-μm resolution using a Genechip System confocal scanner made for Affymetrix by Agilent, Boeblingen, Germany.
Affymetrix Microarray Suite software (version 5.0) Micro-DB, and a data mining tool were used to scan and analyze the relative abundance of transcripts of each gene on the basis of the intensity of the signal from each probe set, and to generate the cluster analysis. The software's analysis parameters were set to values corresponding to moderate stringency (statistical difference threshold = 30, and statistical ratio threshold = 1.5). Output from the microarray analysis was merged with the Unigene or GenBank descriptors and saved as an Excel data spreadsheet. We ran arrays, analyzing infected moDCs from three donors and three controls, for a total of nine comparisons (3 × 3 matrix). For each comparison, the Affymetrix software generates a “difference call” of no change, marginal increase/decrease, or increase/decrease.
The magnitude and direction of change were estimated as the signal log ratio (log scale base 2), and are indicated as calculated expression values in Table 1. The NetAffx database (
Selection of immunorelevant genes differentially regulated after infection of DCs with HCMV TB40E for 24 h. The table contains the mean fold upregulation of three independent moDC cultures. The complete data set can be accessed at the Gene Expression Omnibus (
cDNA synthesis
For cDNA synthesis, 500 ng of RNA was analyzed with the Quantitect Reverse Transcription (RT) Kit with an elongated DNase digestion time of 5 min, and RT reaction for 25 min (Qiagen).
Quantitative real-time PCR assays
Amplification and quantification of cDNA was performed using a standard LightCycler protocol (Roche, Mannheim, Germany) as described elsewhere (24). To confirm upregulated gene expression as detected by array technology, selected genes were analyzed using real-time PCR. The results were normalized against the expression of the housekeeping gene 5-aminolevulinate synthase (hALAS) by evaluating the crossing point (CP) values for the unknown samples using the equation 2 (CP h-ALAS – CP target gene) (18). Sequences of primers and probes are shown in Table 2.
Sandwich ELISA assays
To confirm array data, selected protein levels were measured in the culture supernatant 24 h after infection with TB40E. We used a commercially available colorimetric sandwich enzyme immunoassay technique (Quantikine; R&D Systems, Minneapolis, MN) to analyze 100 μL of the culture supernatant for the presence of IFN-α, IFN-β, CXCL10, and CXCL11, according to the manufacturer's recommendations.
Design of siRNAs and moDC transfection
The 21-nt siRNA duplexes with two 2-end overhang nucleotides have been reported to knockdown expression of a specific gene (6). The siRNAs used for targeted silencing of TLR2 and TLR3 were designed by Qiagen (Table 2). A non-silencing siRNA with random nucleotides and no known specificity was used in each experiment as a negative control.
Determination of transfection efficiency revealed that at least 85% of the cells harbored Cy3-labeled siRNA as demonstrated by FACS analysis and fluorescence microscopy.
Then 1 × 106 moDCs were mixed with no siRNA, with TLR3 siRNA, or non-silencing siRNA (133 nM) in a 4-mm electroporation cuvette in a volume of 100 μL of RPMI 1640 medium. Cell transfection was accomplished with an electroporator that used a rectangle pulse of 340 V for 10 msec (Fischer, Heidelberg, Germany). The cells were incubated for 15 min at room temperature, and then cultivated in 1 mL of RPMI 1640, refobacin, and 10% FCS containing GM-CSF and IL-4. Isolation of RNA, cDNA synthesis, and real-time PCR assays were performed as described above. For statistical evaluation, gene expression after TLR3 siRNA treatment was compared with expression change of the non-silencing control by pairwise comparison using the Student's t-test.
Results
Gene expression profiling
Genes whose expression levels changed in response to HCMV were identified on the basis of repeated differences in their expression levels in TB40E-infected and uninfected samples. In general, expression of 33.5–43.5% of all genes spotted on the array was detected. Only genes with at least twofold change of expression and statistical significance as calculated by t-test (p < 0.05) were considered for analysis.
moDCs incubated with the DC-tropic HCMV strain TB40E for 24 h showed statistically significant upregulation of 466 genes. Among these, a wide range of genes involved in the immune response, such as genes encoding for class I interferons (IFN-α and IFN-β), chemokines (CCL5, CXCL10, and CXCL11), and immunoreceptors (TLR3 and CCR7) were differentially regulated (Table 2). CXCL10 and CXCL11 showed the maximal expression (2040-fold and 993-fold, respectively), followed by interferon-stimulated gene 20 (ISG20), with 848-fold upregulation. Various genes showed strong induction, including tumor necrosis factor superfamily, member 10 (TNFSF10, 406-fold), interferon-induced transmembrane protein 1 (IFITM1, 631-fold), IFN-α (72-fold), and IFN-β (7-fold). Furthermore, many genes involved in the induction of class I interferons and genes, regulated themselves by IFN-α and IFN-β, were found to be strongly upregulated after viral exposure (e.g., IRF1, IRF7, IFI35, and STAT1) (Table 1).
The complete data set can be accessed at the Gene Expression Omnibus (
Confirmation of array data
To confirm upregulation of gene expression patterns detected by array technology, we analyzed expression of some cytokines (CXCL10, CXCL11, IFN-α, IFN-β, and CCL5) and immunoreceptors (TLR2, TLR3, and DC-SIGN) by qRT-PCR in a time-dependent manner (1–6 h and 24 h after infection compared to mock-infected moDCs, respectively).
The PCR assay demonstrated upregulation of cytokines in moDCs infected with TB40E, and showed steep upward trends during the time of viral exposure, as compared to expression of mock-infected controls (Fig. 1). We found that expression of IFN-β was increased as one of the first genes analyzed, and it reached elevated expression levels within 2 h after exposure to HCMV. The strongest upregulation was detected for CXCL10 and CXCL11, supporting our observations obtained by microarray analysis.

Time-dependent expression profiling of DCs infected with HCMV. DCs were infected for 1–6h and 24 h with HCMV TB40E, and expression of CXCL10, CXCL11, IFN-β, IFN-α, and CCL5 (
Expression analysis confirmed that expression of TLR3 was increased after 6 h of infection with TB40E, whereas DC-SIGN and TLR2 expression remained unchanged after viral exposure.
Further verification of array data was done by measuring protein levels in culture supernatants by colorimetric sandwich ELISA. At 24 h after HCMV infection, markedly higher amounts of IFN-α (mean: ∼120 ×), IFN-β (∼8 ×), CXCL10 (∼100 ×), and CXCL11 (∼1000 ×) were observed in the supernatants of moDCs infected with TB40E (data not shown).
Previous studies in mice indicated a role of TLR3 as an important immunoreceptor for MCMC by subsequent triggering of the IFN-β response in murine bone-marrow derived DCs (38). Thus expression of TLR3 in human moDCs was targeted by siRNA prior to stimulation with TLR3 ligand poly I:C or exposure to HCMV TB40E virions.
TLR3 silencing by siRNA
To investigate the role of TLR3 in the HCMV-DC interaction, moDCs were electroporated with either no siRNA (I), with non-silencing siRNA (IV), or with two different TLR3 siRNAs (II and III). Gene expression was analyzed by qRT-PCR assay. Expression of TLR3 was significantly reduced after 24 h in cells transfected with TLR3 siRNA. In contrast, non-silencing siRNA transfection did not significantly change the expression of any of the genes analyzed, and no reduction was observed for the housekeeping gene hALAS. In addition, expression of IFN-α, IFN-β, CCL5, CXCL10, and CXCL11 was not significantly altered by TLR3 siRNA transfection (data not shown).
Stimulation of moDCs with dsRNA (poly I:C) induced strong expression of IFN-β. When moDCs were transfected with TLR3 siRNA prior to poly I:C stimulation, a marked reduction in the expression of TLR3 (siRNA 1: 81% ± 16%, p = 0.011; siRNA 2: 77% ± 21%, p = 0.029), as well as IFN-β (siRNA 1: 44% ± 21%, p = 0.019; siRNA 2: 34% ± 29%, p = 0.041) was detected (Fig. 2). Analysis of cell culture supernatants by ELISA confirmed qRT-PCR results, and showed reduced IFN-β release after silencing of TLR3 (siRNA 1: 62% ± 12%; siRNA 2: 61% ± 11%; p < 0.010). Control experiments, involving LPS stimulation after transfection of moDCs with TLR3 siRNA revealed that expression of IFN-β remained unaltered in comparison to untransfected cells. Thus human moDCs depend on the presence of TLR3 for triggering of poly I:C-mediated expression of IFN-β.
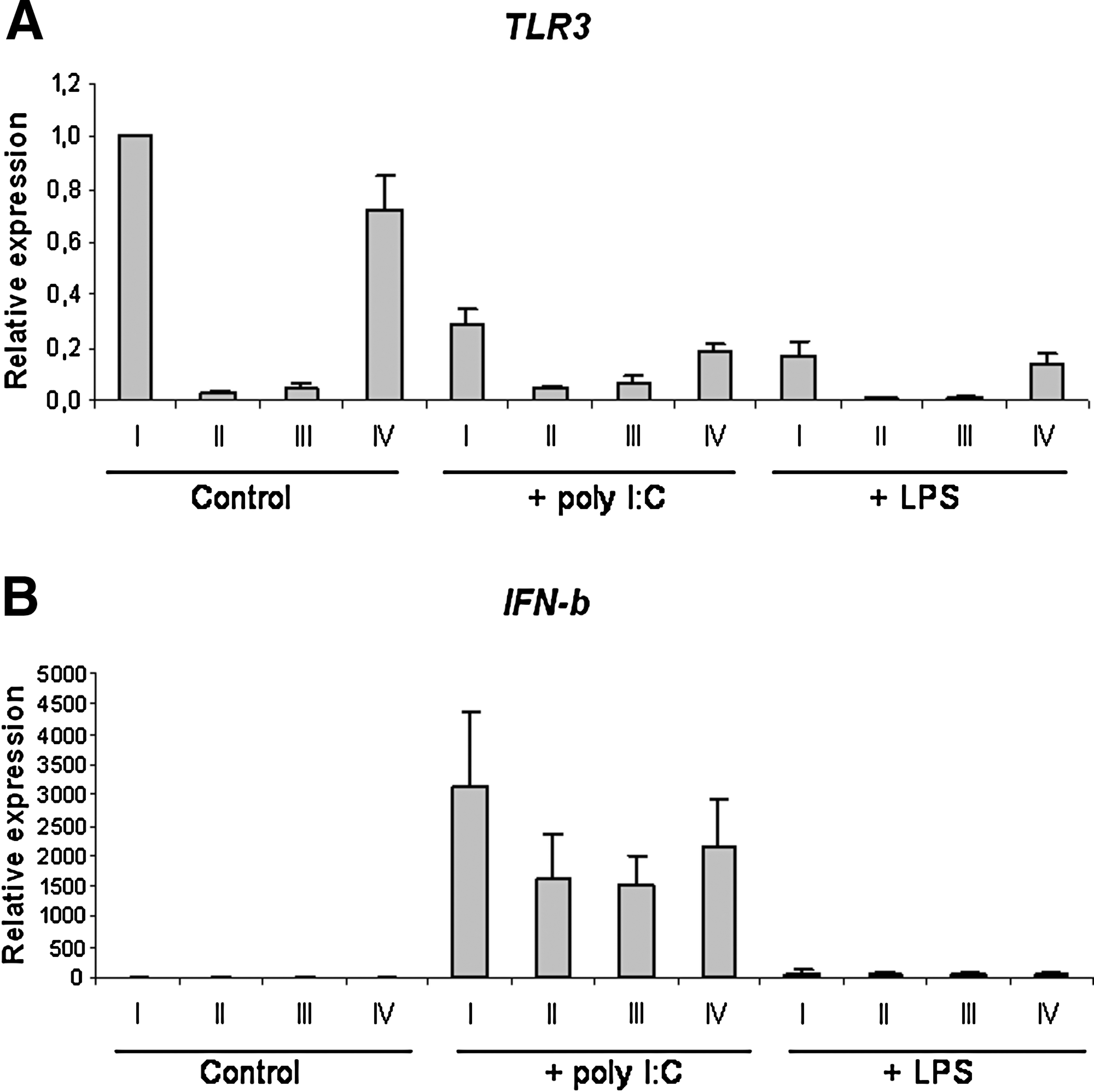
TLR3 mediates expression of IFN-β induced by poly I:C. At 24 h after electroporation with either no siRNA (I), TLR3 siRNA (II and III), or non-silencing siRNA (IV), DCs were stimulated with poly I:C (25 μg/mL) or LPS (10 μg/mL) for 6 h. Total RNA was isolated and reverse transcribed, and cDNA was analyzed by qRT-PCR. Expression of TLR3 (
In contrast to poly I:C stimulation, infection of moDCs with TB40E did not yield a reduction of IFN-β after TLR3 siRNA treatment (Fig. 3). Although a strong reduction of TLR3 was observed (siRNA 1: 84% ± 12%; siRNA 2: 82% ± 12%; p < 0.010), expression of IFN-β was equally upregulated in transfected moDCs as in cells without siRNA, or cells containing non-silencing control siRNA (siRNA 1: −8% ± 63%, p = 0.256; siRNA 2: 1% ± 42%; p = 0.417).
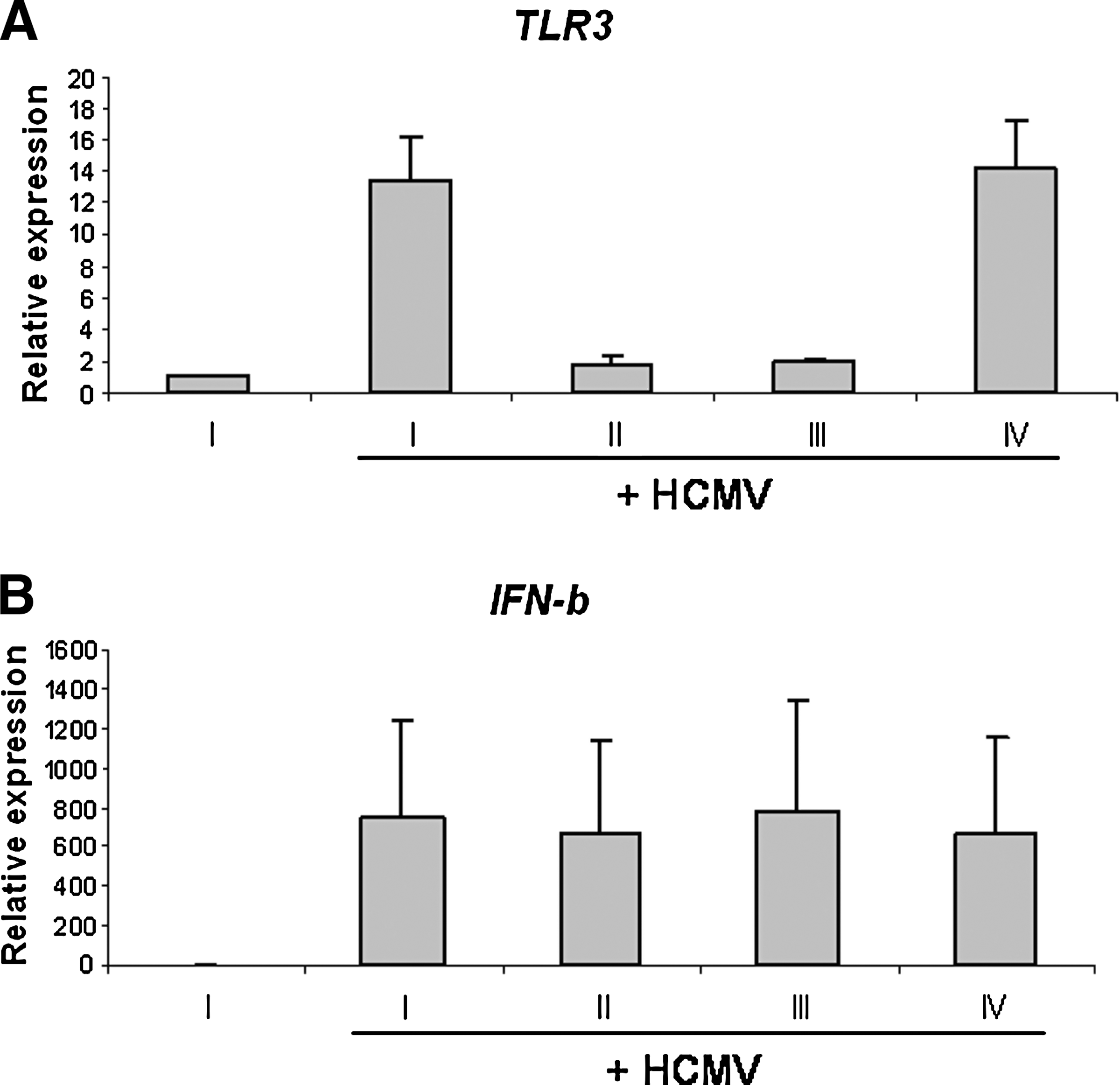
Role of TLR3 in the expression of IFN-β induced by HCMV. At 24 h after transfection with either no siRNA (I), TLR3 siRNA (II and III), or non-silencing siRNA (IV), DCs were exposed to HCMV TB40E (MOI = 50) for 24 h for further expression analysis. These results (TLR3 [
Thus TLR3 does not appear to be of primary relevance for the early induction of IFN-β in human moDCs.
TLR2/TLR9 interaction with HCMV
In addition to the quantification of TLR3, we considered analyzing the expression of two other potentially relevant PRR, TLR2 and TLR9, which might be both involved in the HCMV – DC interaction. However, it has been described previously that TLR9 is not expressed by moDCs (30). By the use of specific primers and probes, we could confirm the observation that TLR9 is absent in moDCs. In addition, stimulation of moDCs with 10 μg of the TLR9 ligand CpG DNA did not result in any activation of moDCs, as indicated by unchanged expression of type I interferons, as well as IL-12 and TNF-α (data not shown).
Furthermore, moDCs were electroporated with 2 different TLR2 siRNAs prior to exposure to HCMV TB40E. When moDCs were transfected with TLR2 siRNA, a marked reduction in the expression of TLR2 (siRNA 1: 92% +/− 3%, p = 0.013; siRNA 2: 87% +/−5%, p = 0.017; n = 4) was achieved. However, no statistically significant reduction for expression of IFN-β was observed in our experimental setting.
Discussion
The DC-tropic strain TB40E goes through the complete reproductive cycle, leading to the production of mature virions (17,34). In this context, our group was able to demonstrate IE antigen expression in moDCs, thereby confirming TB40E replication (11), and recently Kessler et al. (20) showed that IE antigen is expressed in human moDCs 24 h after exposure to TB40E. Furthermore, Jacobs and Langland proposed that HCMV may produce dsRNA as a consequence of bidirectional genomic transcription during the course of infection (16). These dsRNA molecules may induce TLR3-mediated interferon expression (27).
TLR3 was found to activate a MyD88-independent signaling pathway via interferon regulatory factor 3 (IRF3) (9). Activated IRF-3 forms homodimers and translocates to the nucleus, where it activates IFN-α and IFN-β (33,42). This initial burst of interferons acts in an autocrine and paracrine manner through the interferon-α/β receptor and the JAK-STAT pathway to induce a broader panel of ISGs, including another member of the IRF family, IRF-7 (36). Newly synthesized IRF7 is activated and promotes increased expression of IFN-α and IFN-β that cannot be induced by IRF3 alone (31). By using genome-wide arrays, we demonstrated that IRF7, IFN-α, and IFN-β were upregulated in moDCs after infection with TB40E.
Among the most effective innate antiviral responses is the production of IFN-α and IFN-β and the subsequent induction of interferon-stimulated genes (ISGs), such as ISG20, which our array experiments showed to be strongly upregulated in moDCs infected with TB40E. Espert et al. demonstrated that ISG20 is involved in several antiviral functions (7).
Type 1 interferons activate effector cells of the innate immune system, such as NK cells and macrophages, inhibit cell proliferation, and regulate expression of MHC class I antigens. An interferon class I signal provides unique regulatory mechanisms for its own IFN-α/β signaling systems, as well as for the TCR signaling system (39). Moreover, IFN-α and IFN-β have been shown to potently enhance the antibody response through DC stimulation (21).
Infection of moDCs with TB40E induced upregulation of the chemokines CXCL10 and CXCL11. Both are secreted in response to interferons and act as chemoattractants for monocytes and T cells (25). Induction of CXCL11 also requires expression of the signal transducer and activator of transcription 1 (STAT1), which we found to be upregulated.
We also found an upregulation of CCL5 in HCMV-infected moDCs. CCL5 has potent chemotactic and activating properties for activated T cells, NK cells, and DCs. The promoters of CCL5 and CXCL10, like that of IFN-β, contain transcription factor binding elements for NF-κB (10,22).
In order to analyze the interaction between TB40E and TLR3, moDCs were transfected with TLR3-specific siRNAs. We were concerned that double-stranded RNA might induce a nonspecific type 1 IFN response in mammalian cells, leading to arrest of transcription and cell death, and that DCs might be particularly sensitive to double-stranded RNA via the expression of TLR3 (13). Kariko et al. reported that siRNA induced IFN-α, TNF-α, and IFN-β, mediated in part by signaling through TLR3 (19). In contrast, we did not observe any siRNA-mediated class I interferon activation. Additionally, we found no evidence that our siRNAs affected non-targeted gene expression.
Although the immune response to HCMV involves a much larger and more complex cascade, our findings indicate that TLR3 appears not to be the key element of the primary innate immune response to HCMV, as demonstrated by our TLR3 siRNA transfection, poly I:C stimulation, and TB40E infection experiments. This hypothesis is supported by the fact that augmented expression of class I interferon genes can be seen 2 h (IFN-β) and 4 h (IFN-α) post-infection (Fig. 1), whereas differential regulation of TLR3 was noticed only at 5 h post-infection. So we hypothesize that at least the early immune response may be independent of TLR3.
Moreover, TLR2 has been described as a central receptor for the recognition of HCMV virions in fibroblasts, independent of viral replication (4). We analyzed the role of TLR2 in the HCMV-moDC interaction; however, no statistically significant reduction in the expression of IFN-β could be observed.
It is assumed that moDCs must have additional mechanisms for the response to dsRNA, an idea supported by the finding that TLR3−/− cells still react upon poly (I:C) stimulation (32). These mechanisms comprise the dsRNA-dependent protein kinase (PKR), which inhibits host translation, and the IFN-inducible RNA helicase RIG-I as an intracellular PRR for dsRNA. Yoneyama et al. demonstrated that RIG-I is essential for IFN induction, independent of TLR3 (43). In contrast to TLR3, PKR and RIG-I are cytoplasmic, and therefore probably are better suited for the direct recognition of dsRNA produced during bidirectional viral replication. For future experiments, it would be interesting to see if PKR and/or RIG-I can be targeted by our established siRNA system, to uncover any possible involvement of these receptors in the immune response of moDCs to HCMV TB40E.
Conclusion
In conclusion, the DC-tropic HCMV strain TB40E activates and stimulates human moDCs; however, future work is needed to clarify the signaling pathways that ultimately lead to the induction of a class I interferon response in human moDCs. The siRNA system described here may be a valuable tool for further exploration of the HCMV-triggered immune response. The identification of pathways that enhance innate antiviral activity may provide new directions for developing antiviral therapies.
Footnotes
Acknowledgments
This study was supported by research funding from the National Genomic Research Network, the EU project The Development of Immunotherapeutic Strategies to Treat Haematological and Neoplastic Diseases on the Basis of Optimised Allogeneic Stem Cell Transplantation (Allostem; LSHB-CT-2004-503319), the Infectious Diseases Working Party of the European Group for Blood and Marrow Transplantation (EBMT-IDWP), and EuroNet Leukemia (contract no. LSH-2002-2.2.0-3), to strengthen and develop scientific and technological excellence in research, and for the therapy of leukemia by integration of the leading national leukemia networks and their interdisciplinary partner groups in Europe (project no. 503216).
Author Disclosure Statement
The authors have no conflicts with regard to financial interests. This material has not been previously reported and is not under consideration for publication elsewhere.