
Abstract
It is unknown why the influenza B virus causes less severe clinical signs than the influenza A virus in humans. Here we show that influenza B virus induces a lower levels of inflammatory cytokines in the lungs of infected ferrets, and causes less pathological damage to their lung tissues than does influenza A virus. The copy numbers of inflammatory cytokine genes, such as TNF-α and IFN-α, was significantly lower in the lungs of ferrets infected with influenza B virus than in those infected with influenza A virus. There were also significantly lower viral titers in the lungs of ferrets infected with the influenza B virus than with the influenza A virus. In addition, the duration of viral presence was shorter in the lungs of ferrets infected with influenza B virus than with influenza A virus. Taken together, our results suggest that the lower induction of inflammatory cytokines and lower viral titers in the lungs may be responsible for the milder clinical signs seen in ferrets or humans infected with influenza B virus.
Introduction
Influenza B virus is not subtyped. It circulates among humans and it causes clinical signs similar to those of influenza A viruses. Influenza B virus causes morbidity and mortality, and has been responsible for 16 epidemics during the past 70 years (35). The frequency of hospitalization of humans infected with influenza B virus is approximately one-fourth that of those infected with influenza A virus (16). Myositis and gastrointestinal signs are more frequent in humans infected with influenza B virus than in those infected with influenza A virus (4,13). Generally, influenza B virus causes milder clinical signs in infected humans than influenza A viruses.
The pathological signs caused by influenza viruses may be due to inflammatory cytokines, including tumor necrosis factor-α (TNF-α), that are released during the inflammatory response (3,17). In a study of the 1918 pandemic influenza A virus in macaques it was reported that aberrant innate immune responses were responsible for the high mortality rate (17). Infection of cultured human macrophages from monocytes with H5N1 influenza A virus produced proinflammatory cytokines, such as TNF-α, interleukin-1 (IL-1), IL-6, interferon-α/β (IFN-α/β), and cc chemokines, suggesting potentially high mortality (3). TNF-α is a proinflammatory cytokine that is involved in the enhancement of macrophage chemotactic protein 1 (MCP-1) and in MCP-3 gene expression in tissue macrophages, leading to enhanced inflammatory responses. IFN-α/β acts as a major antiviral cytokine by inducing the expression of double-stranded RNA-dependent protein kinase (PKR), RNAase L/2′-5′oligoadenylate synthetase (OAS), and Mx proteins (26,27).
Domestic ferrets (Mustela putorius furo) are the most relevant small animal model for human influenza infection, and have been used to assess influenza pathogenesis and vaccine efficacy (8,19,20). The course of infection and the clinical signs in ferrets infected with influenza A virus closely resemble those in humans.
Influenza B virus causes relatively milder disease in humans than does influenza A virus (16), and one study showed that ferrets were less responsive to infection by influenza B virus (22). Information on the mechanisms of influenza B virus that lead to lower pathogenicity in humans than influenza A virus is limited. To compare the pathogenesis of influenza A and B viruses, we used ferrets that can be readily infected with influenza B and A viruses, because they show clinical signs that are similar to those seen in humans infected with influenza viruses (18 –20). We also compared the induction of inflammatory cytokines in the lungs of ferrets infected with influenza A viruses and influenza B viruses.
Materials and Methods
Viruses
An influenza B virus (B/Malaysia/2506/04, B/Shanghai/ 361/2002) and an influenza A virus, H1N1 (A/New Caledonia/20/99, A/Puerto Rico/8/34), were amplified in 10-day-old hen eggs before they were used in this study. B/Malaysia/2506/04 and A/New Caledonia/20/99 (H1N1) were used as representatives of viruses with high pathogenesis, and B/Shanghai/ 361/2002 and A/Puerto Rico/8/34 (H1N1) were used as representatives of low pathogenesis, based on the clinical signs seen in the infected ferrets in this study.
Infection of ferrets with influenza viruses
Ferrets were purchased from Marshal Farms (North Rose, NY). The ferrets were approximately 7 wk old and were serologically negative for the human influenza viruses H1N1, H3N2, and B by hemagglutination inhibition assay. The ferret studies were performed in a BSL-2+ animal facility. The ferrets were sedated with Zoletil (2 mg/kg; Virbac Laboratories, Carros, France) before they were infected with influenza viruses. The ferrets were intranasally (IN) infected with 1 mL of 6 log10EID50/mL of influenza B virus or influenza A virus. The uninfected ferrets were used as control. The animal experiments were approved by the Animal Experimental Ethics Committee at the Chungnam National University.
Quantification of in-vivo inflammatory cytokines by quantitative real-time PCR
Total RNA was extracted from tissues collected at the left cranial, left caudal, right cranial, or right caudal lung lobes of the infected ferrets (n = 15). To collect total RNA, a 0.1-g segment of lung tissue was added to 1 mL of TRIzol (Invitrogen, Carlsbad, CA) in a 1.5-mL tube and homogenized. The samples were incubated at room temperature for 5 min. Chloroform (200 μL) was then added and the solution was mixed by vortexing. Each tube was then centrifuged for 15 min at 12,000 rpm at 4°C. The upper band containing RNA was collected and placed in another 1.5-mL tube with 500 μL of isopropanol (Sigma, St. Louis, MO). Each tube was centrifuged for 10 min at 10,000 rpm at 4°C. The pellet containing RNA was washed with 100 μL of 75% ethanol in water by centrifuging it for 5 min at 10,000 rpm at 4°C. The washed RNA was resuspended in 50 μL of diethyl pyrocarbonate-treated water.
To quantify the inflammatory cytokines in tissues, real-time PCR was performed as previously described (28), with some modifications. cDNA was made using 10 ng of RNA as previously described (28), and 1 μL of oligo dT primers (0.5 μmol) (Promega, Madison, WI) in a total volume of 9 μL in a 0.05-mL tube. The mixture was incubated for 5 min at 70°C, and then for 5 min at 4°C. Then 4 μL of 25-mM MgCl2, 4 μL of 5X RT enzyme buffer, 1 μL of RNase inhibitor, 1 μL of RT enzyme, and 1 μL of dNTP (10 mM) were added to each reaction tube. They were incubated for 5 min at 25°C, for 60 min and at 42°C, and for 15 min at 70°C.
SYBR green-based real-time PCR using Corbett Roto-Gene 6000 (Mortlake, Australia) and Quantace SensiMix Plus SYBR (London, U.K.) were performed for mRNA targets according to the manufacturer's recommendations. Duplicates of each sample from the ferrets were also run using SYBR green-based real-time PCR.
A total of 20 μL of reaction volume consisted of 2 μL cDNA, 10 μL SYBR mixture (Quantace), 1 μL forward primer (20 pmol), 1 μL reverse primer (20 pmol) (Table 1), and 3 μL water. Forty cycles of PCR were performed, for 5 sec at 95°C, 15 sec at 60°C, and 25 sec at 72°C. To calculate copy numbers, serial dilutions of amplified and eluted ferret glyceraldehyde-3-phosphate dehydrogenase (GAPDH) were performed to generate standard curves from which the concentration of each sample was deduced. All quantifications were normalized to the expression of ferret GAPDH. The copy number of the induced genes was calculated using the formula: number of copies = [template amount (ng) * 6.022 × 1023/template length (bp) * 1 × 109 * 650] (URI Genomics & Sequencing Center, University of Rhode Island, Kingston, RI). The detection limit for the cytokine-specific real-time PCR was 100 copies.
Staining ferret lung tissue with hematoxylin and eosin (H&E)
Each infected ferret (n = 15 per group) was euthanized with a high dose of Zoletil (Virbac Laboratories) before each lobe of the lung was collected. The lung tissues were fixed by submerging them in 10% neutral buffered formalin and embedding them in paraffin. Then 5-μm sections were made and stained with H&E, as previously described (1). The stained tissue sections were evaluated under an Olympus DP70 microscope (Olympus Corporation, Tokyo, Japan).
Viral titration in ferret lung
A 0.05-g section of each lung lobe from the same ferret as that used for staining was homogenized in 0.5 mL of PBS (pH 7.4) supplemented with 2X antibiotic-antimycotic solution (Sigma). The prepared samples were serially diluted 10-fold in PBS (pH 7.4) before each diluted sample was inoculated into 10-day-old embryonated hen eggs. The presence of virus in the inoculated eggs was determined by a hemagglutination assay using 0.5% turkey red blood cells. Viral titers were determined by 1og10 egg infectious dose 50/mL (log10EID50/mL), as previously described (23).
Statistical analysis
Statistical analysis was done using SPSS version 10.0 (SPSS, Inc., Chicago, IL). For comparison of body temperatures, clinical scores, induction of cytokine genes, and viral titers between influenza A virus–infected and influenza B virus–infected ferrets, we used Student's t-test. A p < 0.05 was considered statistically significant.
Results
Observation of body temperatures, body weights, and clinical signs in the infected ferrets
The ferrets were IN inoculated with influenza B virus (B/Malaysia/2506/04 and B/Shanghai/361/2002), or influenza A virus (A/New Caledonia/20/99 [H1N1] and A/Puerto Rico/8/34), and were observed for body temperature, body weight, and clinical signs. Lower body temperatures, body weights, and clinical scores were seen in ferrets infected with influenza B virus compared to those infected with influenza A virus (Fig. 1). The mean peak body temperatures in ferrets infected with influenza B viruses B/Malaysia/2506/04 and B/Shanghai/ 361/2002 were 38.6°C and 38.5°C at 2 d post-infection (p.i.), respectively, while those in ferrets infected with influenza A viruses A/New Caledonia/20/99 (H1N1) and A/Puerto Rico/8/34 (H1N1) were 39.2°C at 3 days p.i. and 39°C at 1 day p.i. (Fig. 1A). The rate of weight gain in ferrets infected with influenza B or influenza A virus was delayed compared to that of uninfected ferrets, but the percentage of weight loss was less in ferrets infected with influenza B virus than in those infected with influenza A virus (Fig. 1B). At 5 days p.i., the mean body weights of ferrets infected with both influenza B viruses (B/Malaysia/2506/04 and B/Shanghai/ 361/2002) were 98% of original body weights, and the mean body weights of ferrets infected with the influenza A viruses (A/New Caledonia/20/99 [H1N1] and A/Puerto Rico/8/34 [H1N1]) were 92% and 96% of their original body weights, respectively. The uninfected ferrets were 120% of their original body weights. The clinical scores were lower in ferrets infected with influenza B virus than in ferrets infected with influenza A virus (Fig. 1C). The peak clinical scores were seen in ferrets infected with both influenza B and influenza A at 5 days p.i. The mean clinical scores of the ferrets infected with the influenza B viruses (B/Malaysia/2506/04 and B/Shanghai/361/2002) were 1.6 and 1.5, respectively (p < 0.05), while those of the ferrets infected with the influenza A viruses (A/New Caledonia/20/99 [H1N1] and A/Puerto Rico/8/34 [H1N1]) were 3.4 and 2.1, respectively, at 5 d p.i.

Change of body temperatures, body weights, and clinical signs in the infected ferrets. Ferrets (n = 5 per group) were IN infected with 1 mL of 6 log10EID50/mL of influenza B virus (B/Malaysia/2506/04 and B/Shanghai/361/2002) or influenza A virus H1N1 (A/New Caledonia/20/99 and A/Puerto Rico/8/34). We recorded rectal temperatures, clinical signs, and body weights daily until 15 days post-infection. Clinical scores were the sum of activity and respiratory signs. The activity was graded as: 0 = normal activity, 1 = calm demeanor, or 2 = depression and loss of interest in play. The respiratory signs were graded as: 0 = normal state, 1 = occasional sneezing and serous node exudates, and 2 = frequent sneezing, coughing, and mucous nose exudates. Data shown are the mean of 5 ferrets ± SD (*p < 0.05). (
Determination of viral titers and histopathology of lung tissues of infected ferrets
We observed milder clinical signs in ferrets infected with influenza B virus than in those infected with influenza A virus (Fig. 1). To determine a possible mechanism that leads to the lower pathogenicity seen in ferrets infected with influenza B virus, we determined the viral titers and pathological damage present in the lungs of the infected ferrets (Fig. 2). The viral titers were determined by log10EID50/mL (Fig. 2A). Viruses were detected in the lungs of ferrets infected with influenza B and influenza A virus up to 5 and 10 days p.i., respectively. The mean viral titers in ferrets infected with influenza B viruses (B/Malaysia/2506/04 and B/Shanghai/361/2002) were 2.2 and 2.5 log10EID50/mL (p < 0.05), respectively, while those in ferrets infected with influenza A viruses (A/New Caledonia/20/99 [H1N1] and A/Puerto Rico/8/34 [H1N1]) were 5.2 and 4.5 log10EID50/mL, respectively, at 3 days p.i.
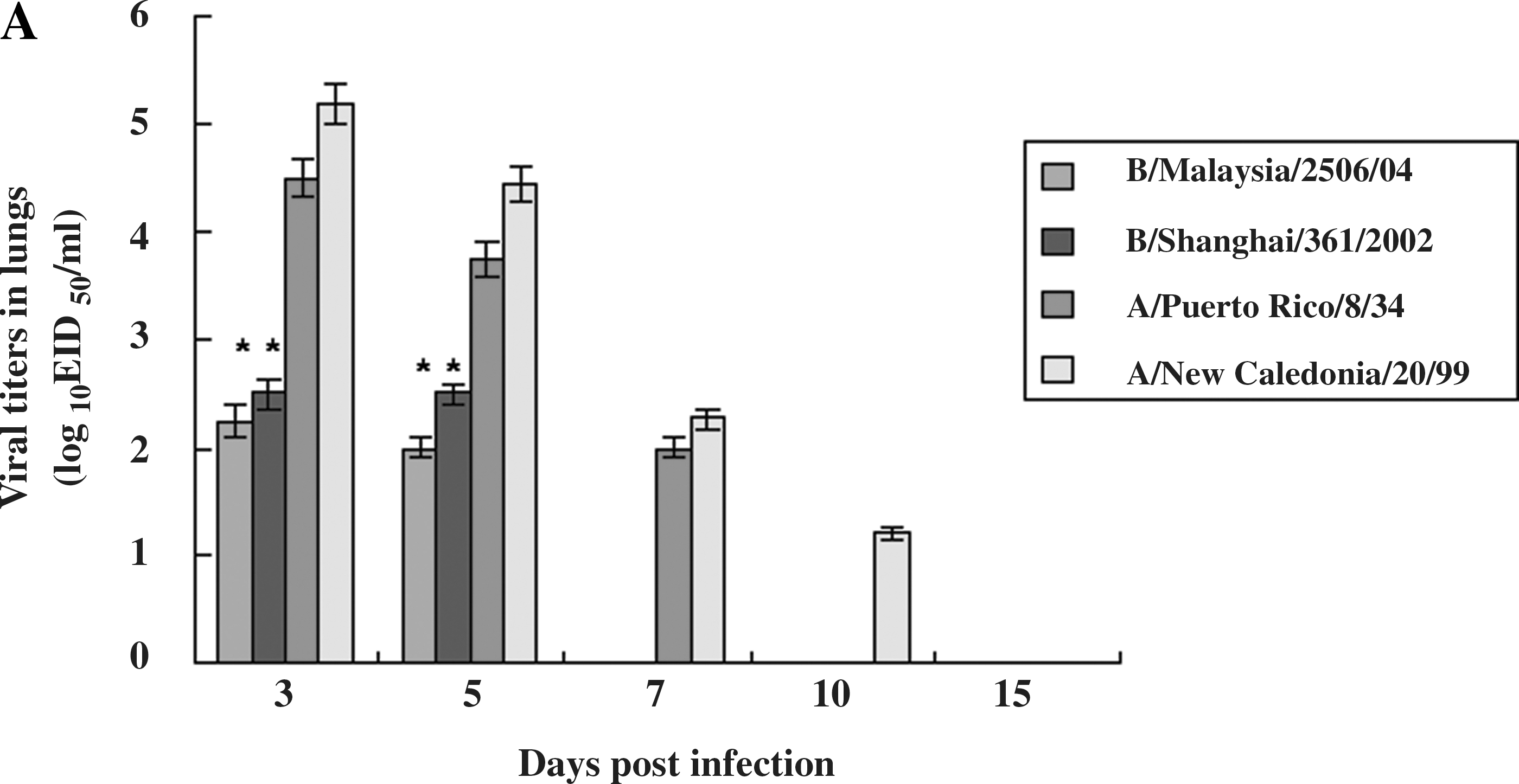
Viral titers and histopathology in the lungs of infected ferrets. (
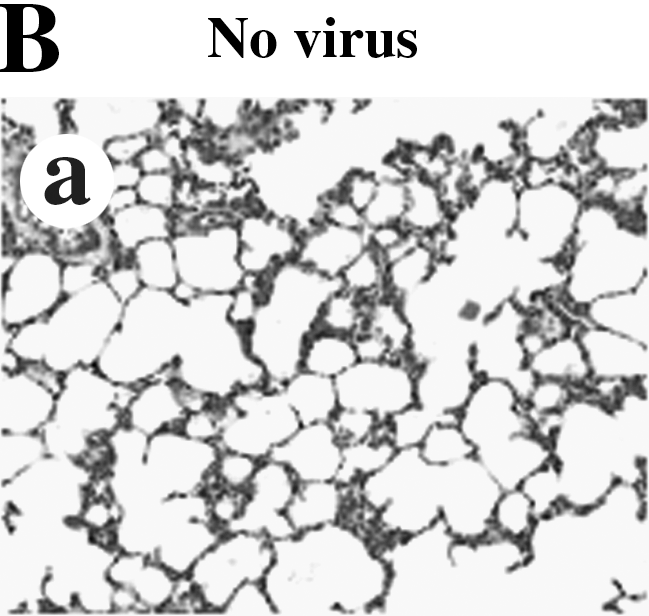

When we stained the lung tissues of ferrets, the lung tissues of ferrets infected with influenza B virus (B/Malaysia/2506/04) showed mild interstitial pneumonia with infiltration of inflammatory cells up to 7 days p.i., while the lung tissues of ferrets infected with influenza A virus (A/New Caledonia/20/99) showed severe interstitial pneumonia with the infiltration of inflammatory cells into alveoli up to 10 days p.i. (Fig. 2B)
Determination of inflammatory cytokines in the lungs of infected ferrets
To determine whether reduced inflammatory responses were induced in ferrets infected with influenza B virus, we measured the induced cytokine mRNA copy number by quantitative real-time PCR. We detected cytokines at the mRNA transcript level because the reagents needed to detect ferret cytokines at the protein level are not available.
Lower mean copy numbers of TNF-α and IFN-α were detected in the lungs of ferrets infected with influenza B virus than in the lungs of ferrets infected with influenza A virus. The induction of TNF-α in the lungs of ferrets infected with both influenza B and influenza A virus peaked at 5 days p.i. The mean fold changes in TNF-α in the lungs of ferrets infected with the influenza B viruses (B/Malaysia/2506/04 and B/Shanghai/ 361/2002) were 30.5 and 31 (p < 0.05), respectively, and those seen in the lungs of ferrets infected with the influenza A viruses (A/New Caledonia/20/99 [H1N1] and A/Puerto Rico/8/34 [H1N1]) were 52.2 and 45, respectively, at 5 d p.i. (Fig. 3A). The induction of IFN-α also peaked in the lungs of ferrets infected with both the influenza B and A viruses at 5 days p.i. However, the induction was lower in the lungs of ferrets infected with influenza B virus than in the lungs of ferrets infected with influenza A virus (Fig. 3B). The mean fold changes in IFN-α in the lungs of ferrets infected with the influenza B viruses (B/Malaysia/2506/04 and B/Shanghai/361/2002) were 17.2 and 18 (p < 0.05), respectively, while those in the lungs of ferrets infected with the influenza A viruses (A/New Caledonia/20/99 [H1N1] and A/Puerto Rico/8/34 [H1N1]) were 114.8 and 88, respectively, at 5 days p.i. IFN-γ was detected in the lungs of ferrets infected with both influenza A and influenza B viruses, but the mean fold change in IFN-γ was lower than those of TNF-α and IFN-α (Fig. 3C). The cytokines IL-4, IL-6, and IL-10 were not detected in the lungs of infected or uninfected ferrets.
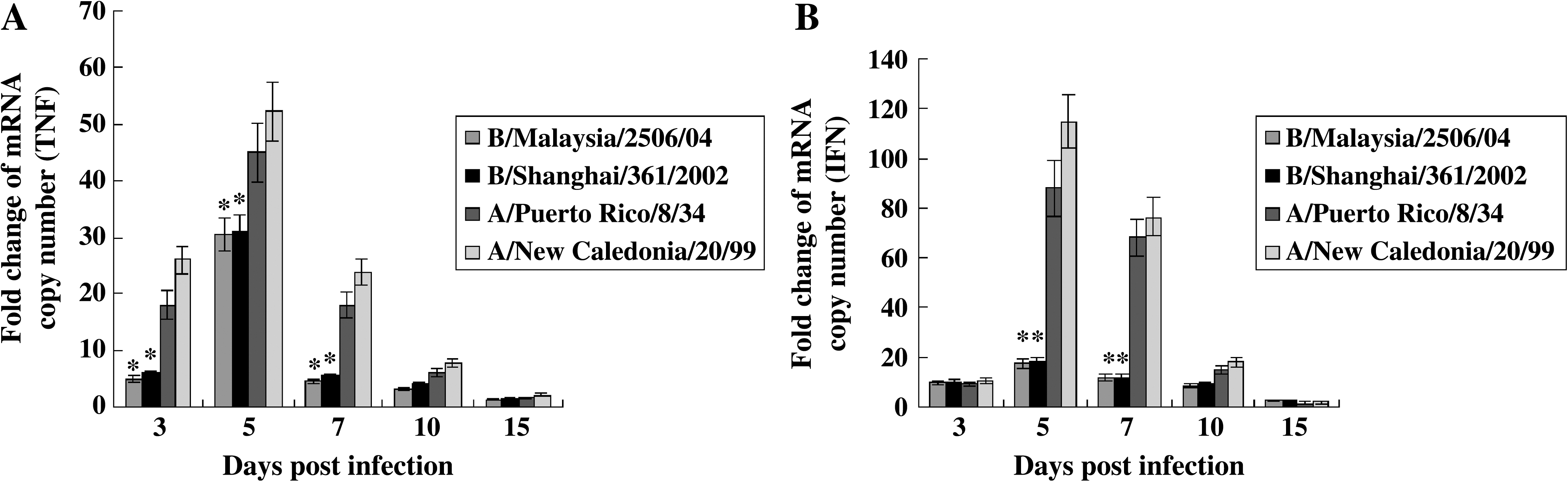
Quantitation of inflammatory cytokines in the lungs of infected ferrets. SYBR green-based real-time PCR was performed for the inflammatory cytokines IFN-α, IFN-γ, TNF-α, IL-4, IL-6, and IL-10. The primers were confirmed using mRNA of ferret peripheral blood lymphocytes stimulated with concanavalin A and lipopolysaccharide by real-time PCR. The lungs of the ferrets were divided into four parts (left cranial, right cranial, left caudal, and right caudal), and the cytokine copy numbers of the parts were combined and the mean copy number in each lung was calculated. The fold change of mRNA copy numbers in the ferrets was determined by comparison with those present in the uninfected ferrets. IL-4, IL-6, and IL-10 were not detected in the lungs of infected or uninfected ferrets. Statistical analysis was performed by comparing the cytokine fold change between influenza A virus–infected and influenza B virus–infected ferrets using Student's t-test. Data shown are the mean of three ferrets ± SD. (

Discussion
The ferret is a good animal model for studying the pathogenesis of influenza viruses because it shows similar clinical signs to those seen in humans (20). In this study, we showed that the clinical and pathological signs in ferrets infected with the influenza B virus were lower than in those infected with the influenza A virus, a pattern similar to that seen in humans. Reduced viral replication and lower induction of inflammatory cytokines were also detected in the lungs of ferrets infected with influenza B virus than in those infected with influenza A virus.
The inflammatory cytokines TNF-α and IFN-α were induced less in the lungs of ferrets infected with influenza B virus than in those infected with influenza A virus. Previous studies suggested that inflammatory cytokines may be the cause of the clinical signs observed in humans infected with influenza virus (6,9). When 19 volunteers were experimentally infected with influenza A virus (A/Texas/36/91 [H1N1]), IL-6 and IFN-α levels peaked in nasal lavage fluid at 2 d p.i., and the levels of these cytokines were correlated with viral titers, temperature, mucous production, and clinical scores. The induction of TNF-α peaked in the nasal fluids at 4 d p.i. (9). Similarly, another study showed that the induction of an inflammatory cytokine, IL-6, was related to the development of nasal symptoms and secretion in 17 volunteers experimentally infected with influenza A virus (A/Kawasaki/86 [H1N1]) (7). A recent study using ferrets infected with the influenza A viruses A/Puerto Rico/8/34 (H1N1), A/USSR/90/77 (H1N1), A/Port Chalmers/1/73 (H3N2), and A/Aichi/2/68 (H3N2), also suggested that virulent influenza viruses can induce higher amounts of IL-6 in nasal fluids (29). In cases of human infection with the highly pathogenic H5N1 influenza virus, increased mRNA levels of TNF-α were detected by reverse transcriptase PCR and immunohistochemistry in the lungs of patients who died of this virus strain (21,32,33). When reagents to detect cytokines on the protein level become available, more research will be needed to correlate cytokine expression with immune response or clinical symptoms in ferrets infected with influenza We did not detect IL-6 in the lungs of ferrets infected with either influenza A or B virus, whereas it was detected in another study (29). The discrepancy between our results and those of the previous study appears to be due to the different tissues we used to detect the inflammatory cytokines; our study used lung tissues and the other researchers used nasal fluids.
The mild pathogenesis seen in ferrets infected with influenza B virus may be related to the lower viral titers and shorter duration viral antigens are present in the lungs of infected ferrets. Our study showed that viral titers were significantly lower in the lungs of ferrets infected with the influenza B virus than in the those of ferrets infected with the influenza A virus. This higher viral replication may cause more pathological damage to lung tissues, resulting in more severe clinical symptoms.
Our results showed that the clinical signs seen in ferrets infected with A/Puerto Rico/8/34 (H1N1) virus were similar to those seen in ferrets infected with influenza B viruses. The reason that A/Puerto Rico/8/34 (H1N1) virus causes milder pathogenesis in ferrets may be due to the multiple passages seen in animals such as mice and ferrets. A previous study showed that A/Puerto Rico/8/34 (H1N1) virus had been passaged over 100 times in mice, ferrets, and embryonated eggs, resulting in attenuation (2).
Our results suggest that the lower induction of TNF-α and IFN-α seen in the lungs of ferrets infected with influenza B virus may be due to reduced replication of the viruses. In this study, the viral titers of ferrets infected with influenza A virus correlated with the induction of inflammatory cytokines such as TNF-α and IFN-α. It appears that inflammatory cytokines are important for controlling influenza virus, and that they cause pathological damage to the lung tissues, resulting in the severe clinical signs seen in animals and humans infected with influenza viruses. Previous studies have demonstrated the importance of inflammatory cytokines for clearing influenza viruses in animals (15,31). A study in mice infected with the 1918 pandemic virus (H1N1) indicated that the depletion of alveolar macrophages that produce TNF-α in mice caused higher mortality (31). A study using human H1N1 influenza virus in pigs also showed that the depletion of alveolar macrophages caused higher mortality and more severe clinical signs compared to controls (15).
Conclusion
The results of this study suggest that the milder pathological signs and symptoms seen in ferrets infected with influenza B virus than in those infected with influenza A virus may be due to lower rates of viral replication and lower levels of induction of inflammatory cytokines such as TNF-α and IFN-α in the lungs of the infected ferrets.
Footnotes
Acknowledgments
We wish to thank Keun Bon Ku for excellent technical support. This study was supported by a grant from the Korea Healthcare technology R&D Project, Ministry for Health, Welfare & Family Affairs, Republic of Korea. We also with to thank HARRISCO, an English editing company, for help editing the manuscript.
Author Disclosure Statement
No conflicting financial interests exist.