
Abstract
Natural infection with Marek's disease virus occurs through the respiratory mucosa after chickens inhale dander shed from infected chickens. The early events in the lung following exposure to the feather and squamous epithelial cell debris containing the viral particles remain unclear. In order to elucidate the virological and immunological consequences of MDV infection for the respiratory tract, chickens were infected by intratracheal administration of infective dander. Differences between susceptible and resistant chickens were immediately apparent, with delayed viral replication and earlier onset of interferon (IFN)-γ production in the latter. CD4+ and CD8+ T cells surrounded infected cells in the lung. Although viral replication was evident in macrophages, pulmonary B cells were the main target cell type in susceptible chickens following intratracheal infection with MDV. In accordance, depletion of B cells curtailed viremia and substantially affected pathogenesis in susceptible chickens. Together the data described here demonstrate the role of pulmonary B cells as the primary and predominant target cells and their importance for MDV pathogenesis.
Introduction
Most studies of MDV pathogenesis have involved cell-associated MDV, and cell-free MDV or MD vaccines administered by a parenteral route. Although these approaches have elucidated the various phases of MD pathogenesis, inadvertently they exempt crucial events that occur directly after natural MDV infection via the respiratory tract, and as a result little is known about this early phase. In the present study, we investigated early events after intratracheal infection to identify infected cells and the factors contributing to early MDV pathogenesis. The infection method used is reliable and allows a precisely-timed, measurable dose of native MDV to be delivered to the natural portal of entry. We show increased and earlier viral replication in the lungs of susceptible line 7 chickens compared to resistant line 6 chickens. IFN-γ expression in the lung increased after infection, but was delayed in susceptible chickens. The predominant target cells for viral replication are pulmonary B cells, although macrophages can also become infected later in the response.
Materials and Methods
Experimental animals
Chickens were obtained from parent flocks of outbred Rhode Island Red (RIR), inbred White Leghorn subline 72 (line 7, MD-susceptible), or inbred White Leghorn subline 61 (line 6, MD-resistant), and maintained in disease-free conditions at the Institute for Animal Health. The parent flocks were not vaccinated and tested negative for antibodies to a range of poultry pathogens, including MDV, infectious bursal disease virus (IBDV), and chicken infectious anemia virus (CIAV). Their progeny were free of maternal antibodies to MDV. Experimental birds were housed in wire cages in isolation rooms entered through an air lock. Each room was provided with high efficiency particle air (HEPA)-filtered air under positive pressure. The rooms were hosed down daily to prevent the build-up of dander on the floor and walls. Food and water were available ad libitum throughout. All animal procedures were performed in accordance with local ethical regulations, and were approved by the U.K. Home Office. Unless otherwise stated, the chickens were 2 wk of age when infected with MDV.
Dander isolation for infecting chickens
Thirty RIR chickens were infected by intra-abdominal injection of 1000 pfu of a very virulent strain of serotype-1 MDV, RB1B (46), from the 11th passage in primary chicken kidney cells. Three weeks after infection, dander was collected from the extract filters of the isolation room. This dander was coarsely filtered using a household flour sieve followed by 315-μm steel mesh, desiccated overnight and stored in 2-g quantities in sealed tubes below −70°C. Control dander was obtained from the extract filters of a similar room housing uninfected RIR chicks and treated in the same way. The same batches of dander were used for all of the experiments.
Experimental design
Known quantities of a suspension of dander were delivered into the lower trachea by carefully inserting the delivery tube of a PennCentury microaerosilizerTM (PennCentury, Philadelphia, PA) through the glottis and insufflating the dander using 2–3 mL of air. Infected chickens were killed when clinical signs of MD reached a predetermined clinical end-point: unable or reluctant to walk (paretic limbs), disinterested in food or water, assuming a hunched posture with drooping wings and head, displaying loss of weight, and cloudy partially-closed eyes.
Titration of infective dander
The minimum infecting dose of the batch of MDV-infective dander needed to induce clinical signs of MD was determined by insufflating a standard amount (5 mg) of dander containing different proportions of the MDV-infective material: 5, 2.5, 1, 0.1, 0.01, 0.002, or 0.001 mg dose (n = 8). Another 8 chickens were treated with 5 mg of the uninfected dander. Blood samples (usually 50 μL) were obtained by venipuncture at 3, 7, 10, 14, 21, 35, 48, and 62 d post-infection (dpi) to monitor MDV viremia by quantitative PCR, and on some occasions test for antibodies against IBDV and CIAV by ELISA (Idexx, Westbrook, USA). The chickens were monitored for clinical signs of MD for 63 d, after which the experiment was terminated. At 63 dpi all remaining birds were killed, and liver, heart, and brachial/sciatic nerves were removed and fixed in buffered formaldehyde-saline (pH 7.5) prior to histological processing and staining with hematoxylin and eosin. Tissue sections were examined and scored blindly for lymphocyte infiltration using the MD lesion scoring system described by Burgess et al. (13).
Early events after natural infection
Thirty chickens from each of the resistant (line 6) and susceptible (line 7) lines were infected by intratracheal insufflation of 2.5 mg of dander. Thirty age-matched birds from each line were used as controls and given 2.5 mg of noninfective dander by the same route. Spleen and lung samples were collected post mortem from six infected and six uninfected chickens of each line at 1, 2, 3, 4, and 5 dpi. Leukocytes were isolated from the spleen and one of the lungs from each chicken and isolated for analysis by flow cytometry and for DNA/RNA extraction. The remaining lung was removed and prepared for cryosectioning and immunostaining.
Depletion of B cells and cell sorting after natural infection
Sixty chicks from each of the resistant (line 6) and susceptible (line 7) lines were surgically bursectomized at day 18 of embryonation and allowed to hatch. At 2 wk of age, blood B-cell levels were checked, and only those whose B cells were depleted were infected by intratracheal insufflation of 2.5 mg of dander. In addition, an age-matched control group was infected with 2.5 mg of dander. Blood samples (50 μL) were collected from the wing veins at 3, 7, 10, 16, 23, and 41 dpi for DNA isolation. At 5 and 20 dpi, spleens (n = 3) were removed and single-cell suspensions were stained with CT4-FITC (CD4 cells), AV20-PE (B cells), AV14-AlexaFluor-646 (CD8 cells), and KUL-01-biotin/SA-PerCP (macrophages). Cells were sorted four ways using a MoFlo (Dako Cytomation, Glostrup, Denmark), and RNA was extracted from the individual populations.
Preparation of cell suspensions
Spleens were extracted and homogenized over cell sieves (Becton Dickinson, Frnaklin Lakes, NJ) and blood was drawn from the jugular vein using a 23-gauge needle. Peripheral blood leukocytes (PBL), lung leukocytes, and spleen leukocytes were purified by density gradient centrifugation using Ficoll-Paque Plus (Amersham Pharmacia Biotech, Little Chalfont, UK). Leukocytes were prepared from the spleens by passing through a 70-μm cell strainer, and lung leukocytes were further purified by enzymatic digestion of the lung. Briefly, lungs were washed in PBS and injected in several places with a total of 3–4 mL of enzyme cocktail (556 μg/mL DNase I [Boehringer Mannheim, Bracknell, U.K.] and 2.2 mg/mL collagenase D [Boehringer Mannheim] in HBSS [Sigma, Gillingham, U.K.]). After incubation at 37°C for 15 min, the first suspension was discarded. Digestions were repeated until the lung tissue had completely disintegrated and subsequent cell suspensions were transferred to 10 mM EDTA in HBSS without calcium. The collected cell suspension was filtered using a 70-μm cell strainer and layered onto Ficoll-Paque Plus.
Quantification of MDV and cytokine transcripts
The amount of MDV present in the DNA isolated from PBL (DNeasy© 96 tissue kit; Qiagen, Crawley, U.K.) was quantified using a real-time quantitative PCR assay (TaqmanTM; Applied Biosystems, Foster City, CA) for the MDV Meq gene, essentially as described by Kaiser et al. (33). Meq reaction mixtures included primers and probe to an avian housekeeping gene, ovotransferrin (Ovo), labeled with a second fluorescent dye (VICTM; Applied Biosystems). Raw data were normalized to a positive control included in every run to exclude plate-to-plate variation (14).
Reverse transcription of isolated RNA (RNeasy© 96 tissue kit; Qiagen) was performed using a single-tube single-enzyme system reaction (Taqman EZ RT-PCR Kit; Applied Biosystems), essentially as described by Kaiser et al. (33). RNA was analyzed using primer and probe for cytokine transcripts (24) and viral transcripts (14). All cytokine/MDV reactions were corrected for the amount of cellular RNA by using 28S RNA (housekeeping gene) as described for quantitative PCR reactions.
Isolation and quantification of MDV in dander
DNA was extracted from 2.5 mg of infective dander and 1000 pfu cell-associated MDV for comparison of viral load. A sample (2.5 mg) of the dander was added to 10 mL of TE buffer (1 mM EDTA, 10 mM Tris-HCl [pH 8.0], 2% v/v nonidet P40, and 0.05% v/v Tween-20) and incubated overnight, and after sonication (3 × 30 sec, Soniprep, London, UK), was digested in proteinase K (1 mg/mL) in 1% (w/v) SDS at 45°C for 8 h. The supernatant was aspirated after centrifugation (300 × g for 5 min) and DNA was extracted using a DNeasy tissue kit (Qiagen). Viral DNA was quantified using real-time quantitative PCR assay using the Meq gene.
Fluorescent labeling of cell subsets and flow cytometry
Immunofluorescent staining of leukocytes for flow cytometry analysis was used to quantify relative changes in lymphocyte subpopulations. All antibodies were diluted 1:200 in PBS-BSA (0.4% w/v)-azide (0.1% w/v), and 1 × 106 leukocytes were stained with anti-CD8β-biotin (51), followed by streptavidin-APC (Southern Biotechnology Associates, Birmingham, AL). Following incubation, washing, and blocking with 5% normal mouse serum, the cells were incubated with directly-conjugated anti-chicken monoclonal antibodies CD4-FITC (19), CD3-PE (20), and CD8α-PerCP (51). In separate tubes, duplicate samples were stained for macrophages (KUL-01) (37), followed by Ig(H+L)-FITC (Southern Biotechnology), and then B cells (Bu1-PE) (43). Lymphocyte suspensions were analyzed using a FACScalibur (Becton-Dickinson) and FCSExpress software version 1.0 (DeNovo Software, Los Angeles, CA).
Immunostaining of tissue sections
Tissues sections (6 μm) were cut using a cryostat (Leica CM1900; Leica, Bannockburn, IL), air-dried overnight, and fixed in acetone for 10 min. Slides were stored airtight at −70°C until immunohistochemistry. All antibodies were diluted in 0.4% (w/v) BSA-PBS and incubated for 60 min. CD8α+ T cells were labeled (AV14, 1:25) and visualized using the Vectastain Elite ABC kit (Vector Laboratories, Peterborough, U.K.), and the Vector NovaRed substrate kit (Vector Laboratories), according to the manufacturer's instructions. Sections were counterstained with hematoxylin (Gill no. 3; Sigma) and permanently mounted in Clearium (Surgipath, Peterborough, U.K.).
Cryosections for immunofluorescent staining were prepared as above. Nonspecific reactivity was blocked by incubation with 5% (v/v) normal mouse serum for 20 min. Macrophages (KUL-01, 1:30) (37), B cells (Bu1, 1:200) (43), CD4+ cells (AV29, 1:30), and CD8α+ T cells (AV14, 1:25) were visualized by Alexa568-conjugated goat anti-mouse IgG H/L (Molecular Probes, Eugene, OR). Where indicated, sections were double-stained with the viral protein pp38 using Alexa488-conjugated BD1 (1:100) (36). Nuclei were stained by incubation with 1.09 μM DAPI (Sigma) for 10 min. Sections were mounted in Vectashield (Vector Laboratories) and analyzed using a confocal microscope (Leica SP2 with 405-, 488-, and 568-nm lasers). Control sections were treated similarly, but were incubated with isotype control antibodies.
Statistical analysis
Mean lesion scores between multiple groups were compared using the Kruskal-Wallis test. Where an overall level of significance was observed, Dunn's test was conducted to determine significance between groups. Mortality data were analyzed by the Kaplan and Meier method. Significance in viral load was analyzed using analysis of variance followed by Tukey's test, where overall significance was observed. Statistical analysis was performed using Graphpad Prism software version 4 (GraphPad Software, San Diego, CA).
Results
Intratracheal administration of MDV-infective dander to mimic the respiratory infection route
An infection model was established to study the early events following pulmonary exposure to MDV particles in their natural form. Infective or control dander were obtained from the extract vents of rooms housing chickens either infected with cell-associated RB1B virus or control chickens, respectively. The dander consisted of dried sloughed flakes of skin and fragments of feather barbules, and contained small numbers of micrococci (gram-positive cocci in clusters) and gram-negative lactose-positive and lactose-negative rods (data not shown). Overall there were no major differences between the two preparations. The presence of viral particles in the infective dander was confirmed by extracting MDV DNA and analysis by real-time PCR (data not shown). Insufflating this material deep into the trachea using a dry-material microaerosolizer appeared to cause only minor interference with respiratory function, and some nonspecific inflammatory changes from which the chicks quickly recovered. Preliminary experiments using a range of doses (2.5–20 mg) of MDV-infective dander demonstrated that susceptible RIR chickens developed MD (data not shown). No clinical signs were observed when noninfective dander was administered. After insufflation with either infective or noninfective dander, no clinical or serological manifestation of any other lymphotrophic virus (IBDV or CIAV) infection was observed. MDV ICP4 DNA could readily be detected in PBL as early as 5 d after instillation of 2.5–20 mg of dander.
The initial experiments did not take into account the inflammation caused by the bolus of dander administered and its potential effect on MD pathogenesis. Different doses of MDV-infective dander were tested for their ability to induce MD in susceptible line 7 chickens while standardizing the total administered bolus, and 5 mg of total dander containing different ratios of MDV-infective and noninfective dander was administered to the chickens, which were monitored for clinical signs, MD lesions, and viremia (Fig. 1). More than 50% of chickens given the higher doses of infective dander (1, 2.5, and 5 mg) developed clinical signs of MD by 10–12 dpi, whereas these were significantly (p < 0.001) delayed in chickens given the lower doses (0.001, 0.002, and 0.01 mg), which survived to the end of the experiment at 63 dpi.

Natural infection of susceptible line 7 chickens with MDV induces morbidity and pathology in a dose-dependent manner. Natural infection was titrated by intratracheal instillation of 5, 2.5, 1, 0.1, 0.01, 0.002, or 0.001 mg of RB1B-infective dander bulked up to 5 mg with noninfective dander into susceptible line 7 chickens. Controls received 5 mg of noninfective dander. (
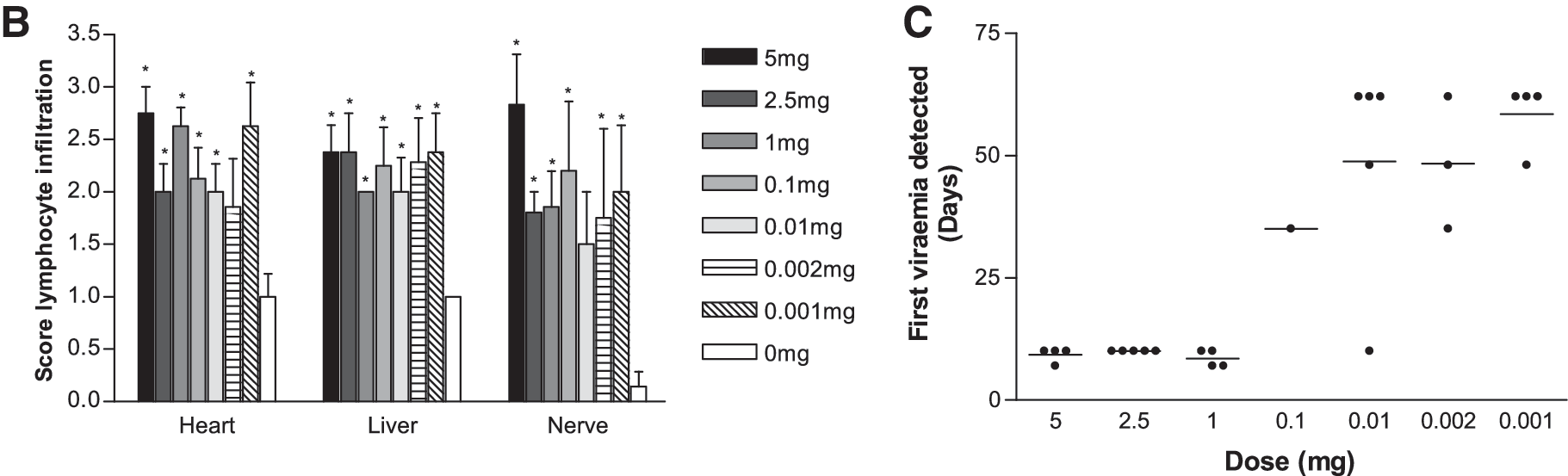
Tissues removed post-mortem were examined for the presence of lymphoid lesions by histology. Histology was evaluated at the onset of clinical signs rather than at set time points. All chickens treated with the MDV-infective dander had significantly (p < 0.05) greater lymphocyte infiltration into the liver, heart, and nerve post-mortem compared to uninfected birds, but there was no clear evidence that this infiltration was dependent on the treatment dose (Fig. 1B).
MDV levels in PBL were determined by quantitative PCR assay. Viral DNA was not detected in the samples from chickens treated with the noninfective dander (data not shown). The onset of a detectable viremia in infected groups was delayed in chickens given smaller doses of infective dander (Fig. 1C). MDV was detected from 7 dpi onwards in the chickens receiving the higher doses (5, 2.5, and 1 mg), but later (from 21 dpi onwards) in chickens receiving the lower doses of MDV-infective dander. In all future experiments 2.5 mg of infective dander was used for intratracheal infection, as this dose reliably and reproducibly induced MD in susceptible (line 7 and RIR) chickens.
Early viral replication in lung leukocytes of susceptible chickens
The viral events during the first 5 d after pulmonary exposure to 2.5 mg MDV-infective or noninfective dander were investigated in chickens susceptible and those resistant to MD. DNA was isolated from lung and spleen leukocytes daily after insufflation and tested for the presence of the Meq gene as an indication of the viral load present (Fig. 2A). Viral DNA was detected as early as 2 dpi in lung leukocytes from susceptible chickens, and was delayed by 1 d in the resistant chickens, and both increased with time. The earliest evidence of MDV in spleen leukocytes was not until a day later in susceptible chickens, at 3 dpi, with a 1-d delay in the leukocytes from the resistant chickens.

Early MDV replication in lung and spleen after natural infection. Resistant line 6 and susceptible line 7 chickens were infected by intratracheal instillation of 2.5 mg of RB1B-infective dander and harvested daily for 5 d. DNA and RNA were isolated from lung and spleen leukocytes. (
The presence of MDV genomes was indicative of viral particles, but did not demonstrate viral replication. Therefore, DNase-treated RNA from lung and spleen leukocytes was analyzed by quantitative RT-PCR assay using specific primers and probes for immediate early (ICP4), early (pp38), and late (gB) gene transcripts. Viral RNA (pp38) was first detected in lung leukocytes of susceptible chickens at 2 dpi, and of resistant chickens at 3 dpi (Fig. 2B). Similarly to the viral load, detection of pp38 transcripts in spleen leukocytes was delayed by 1 d compared with the respective values for the lung lymphocytes in both genotypes, which increased with time. From the time of detection, viral replication in the resistant leukocytes remained lower than that in the susceptible leukocytes (significance at 5 dpi was p < 0.001). Similar patterns were observed for the gB and ICP4 transcripts (data not shown).
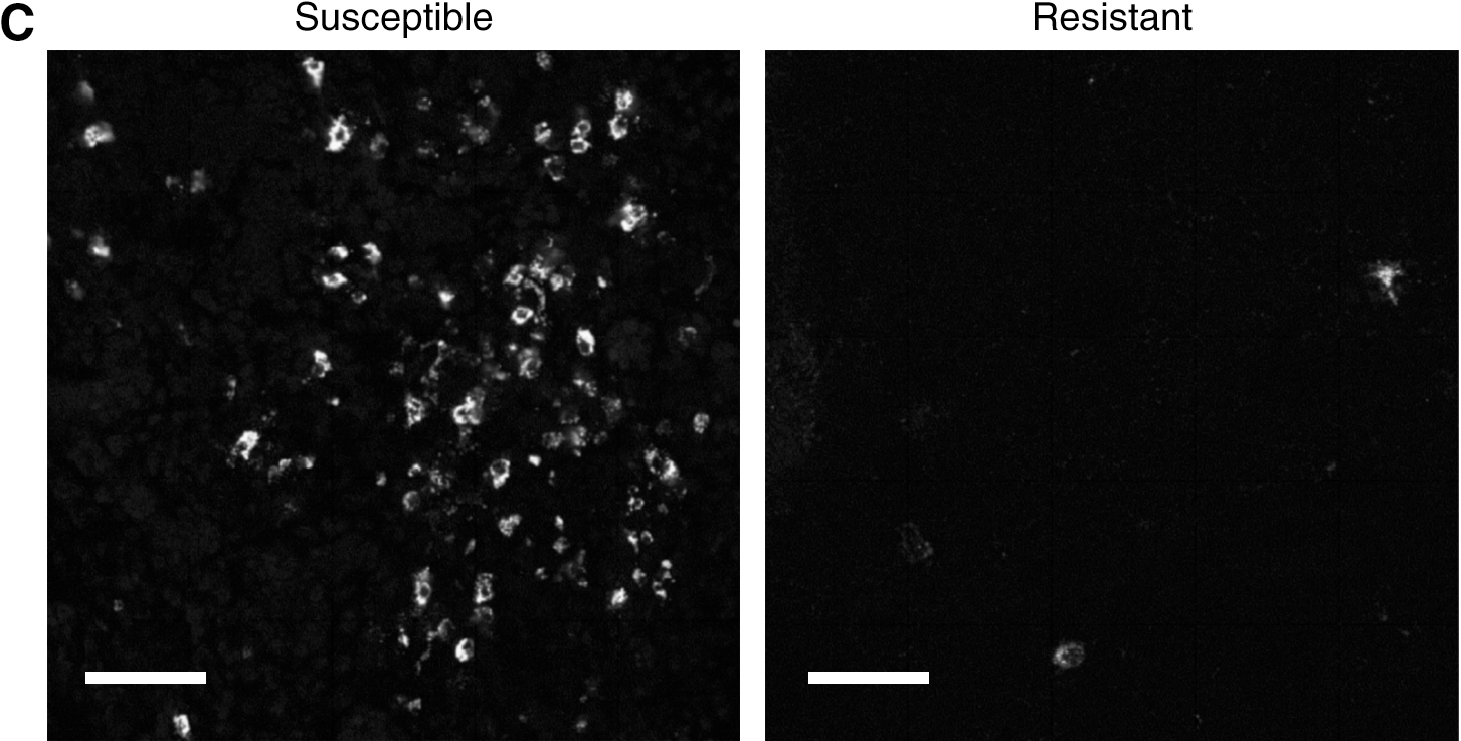
Viral replication was also evident when lung sections were stained for pp38 antigen using immunofluorescence (Fig. 2C). Five days after infection a substantial amount of viral antigen was present in the lungs of susceptible chickens, but not resistant chickens, confirming the results obtained using RT-PCR.
Inflammatory cytokine transcript levels in lung and spleen leukocytes
The effect of early viral replication on cytokine levels was assessed, as they could play an important role in MDV pathogenesis. Transcripts for IFN-α, IFN-β, IFN-γ, IL-4, IL-6, IL-18, and TGF-β in lung and splenic leukocytes were measured using quantitative RT-PCR assay. Intratracheal infection with MDV resulted in a substantial and sustained increase in IFN-γ transcripts as early as 3 dpi in the lungs of resistant chickens compared to controls, and was delayed in the susceptible chickens (Fig. 3A). No substantial differences based on either genotype or infection could be demonstrated for the other cytokines in lung leukocytes (data not shown). During the first 3 d after intratracheal infection, there was little effect on the transcript levels of any of the cytokines in spleen leukocytes compared with birds treated with noninfective dander (Fig. 3). However, by 4 dpi there was an increase (4–8 times) in the transcript levels of IFN-α, IFN-β, IFN-γ, IL-6, and IL-18 in susceptible leukocytes. This increase was also evident at 5 dpi, especially for IFN-γ and IL-6, for which a 16-fold increase was observed (p < 0.01 and p < 0.05, respectively). In the resistant spleen leukocytes this increase was minimal even at 5 dpi, and was restricted to IL-6, IFN-γ, and IL-18. The observed increase in spleen leukocytes from susceptible chickens was significantly different (p < 0.05) from that of resistant chickens. No differences based on genotype or infection were observed for either IL-4 or TGF-β in the spleen (data not shown). Thus, susceptible chickens seem to have a delay in IFN-γ production in the lung and a more profound inflammatory cytokine profile in the spleen.
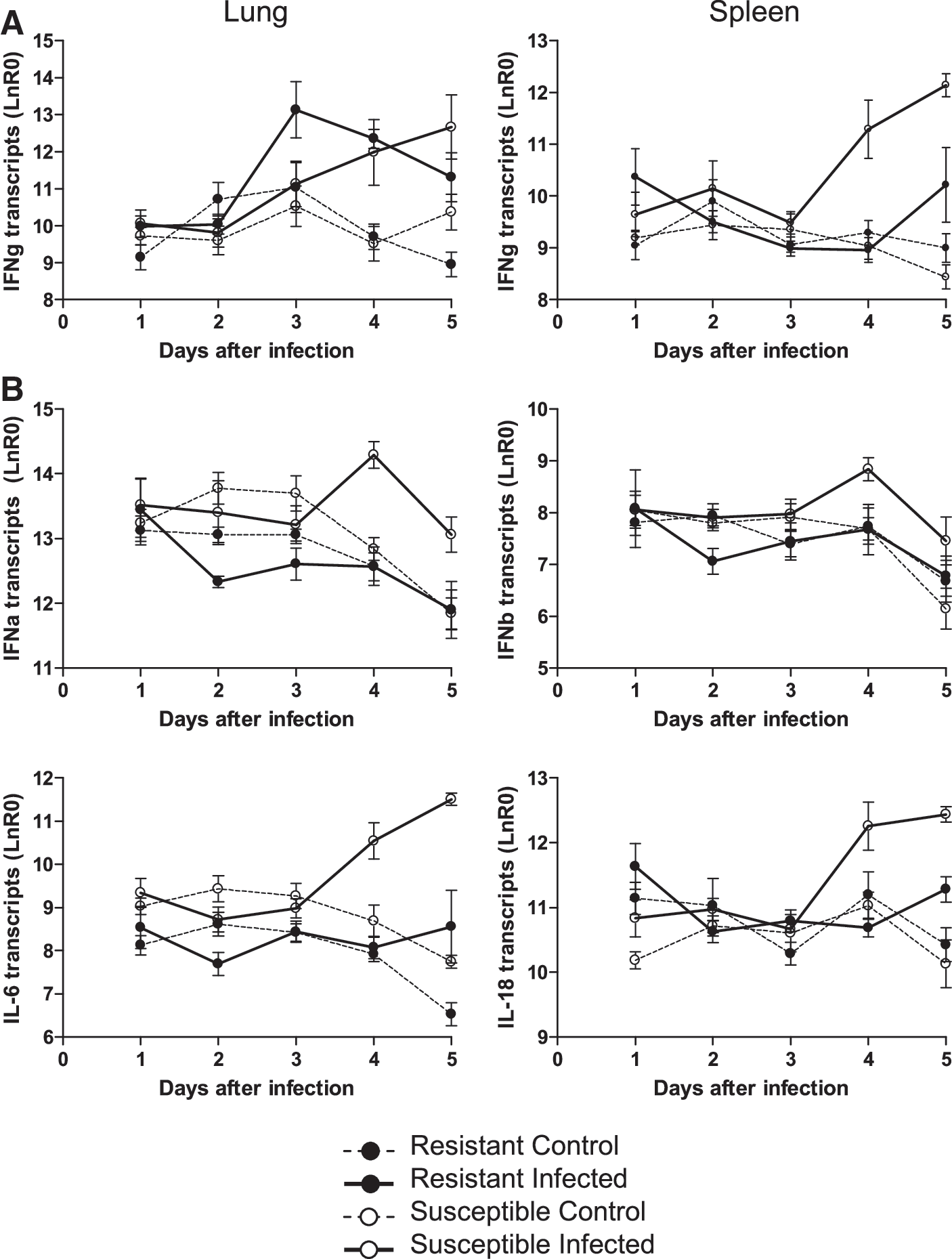
Cytokine transcript levels in lung and spleen leukocytes of resistant and susceptible birds after intratracheal insufflation with RB1B-infective or control dander. (
Increased CD8 T-cell population in the lung after infection
The effect of intratracheal insufflation of infective dander on proportions and location of lymphoid cells was investigated using flow cytometry. The percentage of T cells, B cells, and macrophages in the lungs and spleens of resistant and susceptible chickens during early viral replication was determined (Fig. 4). Overall, infection caused a significant increase (p < 0.01) in the ratio of lymphocytes in the lungs of both genotypes from 4 dpi onward, as measured by CD45 expression (data not shown). In the lung, the proportion of CD3+CD8αβ+ leukocytes increased from 4 dpi after insufflation in the infected compared to uninfected groups to around fourfold at 5 dpi in the susceptible chickens (p < 0.05). KUL-01+ macrophages displayed a similar profile, increasing to twofold over the uninfected controls by day 5 (p < 0.05). Pulmonary B cells were unaffected in number and remarkably similar between the groups. In the spleen, the percentage of B cells was significantly higher in resistant versus susceptible chickens, with the opposite being demonstrated for T cells. Genotype appeared to affect the cellular composition significantly more in the spleen. Other differences were restricted to a reduction in CD8αβ+ T cells at 4 and 5 dpi, and B cells at 5 dpi in the spleens of infected susceptible chickens.
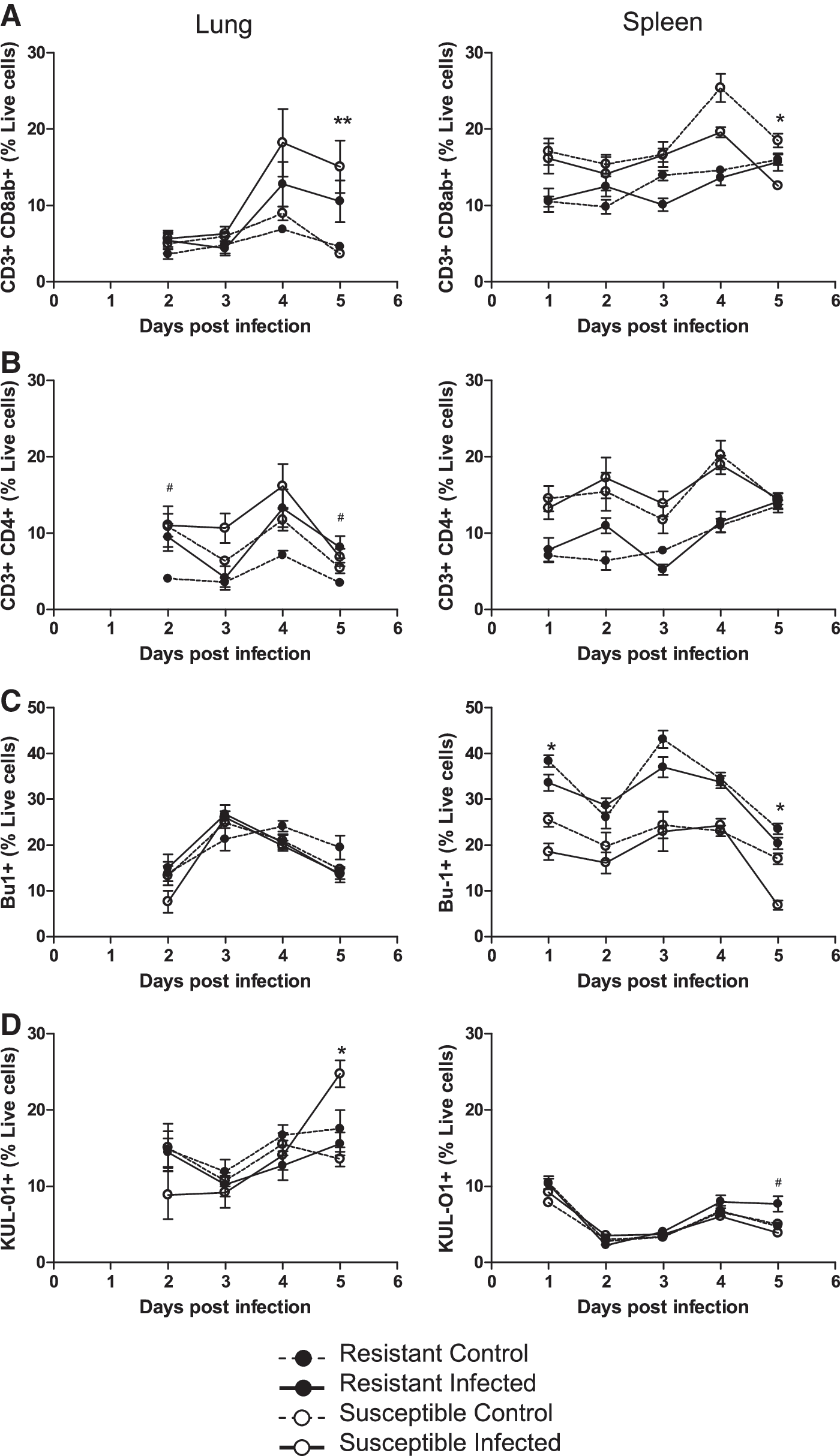
Early changes in cell populations in the lungs and spleens following pulmonary infection. Single-cell analysis of lung and spleen leukocytes from resistant line 6 and susceptible line 7 chickens following pulmonary exposure to RB1B-infective or noninfective dander. CD8 T cells (
As the predominant change in cellular ratio was observed in lung CD8αβ+ T cells, the location of these cells following intratracheal infection was investigated using immunohistochemistry (Fig. 5). An increase in the proportion of CD8α+ pulmonary leukocytes was evident in both chicken lines following infection. The number of positive cells increased with time, especially in susceptible chickens, in which the CD8α+ cells occupied a large proportion of the lung from 3 dpi onward. The CD8 T cells were usually located adjacent to pulmonary spaces.
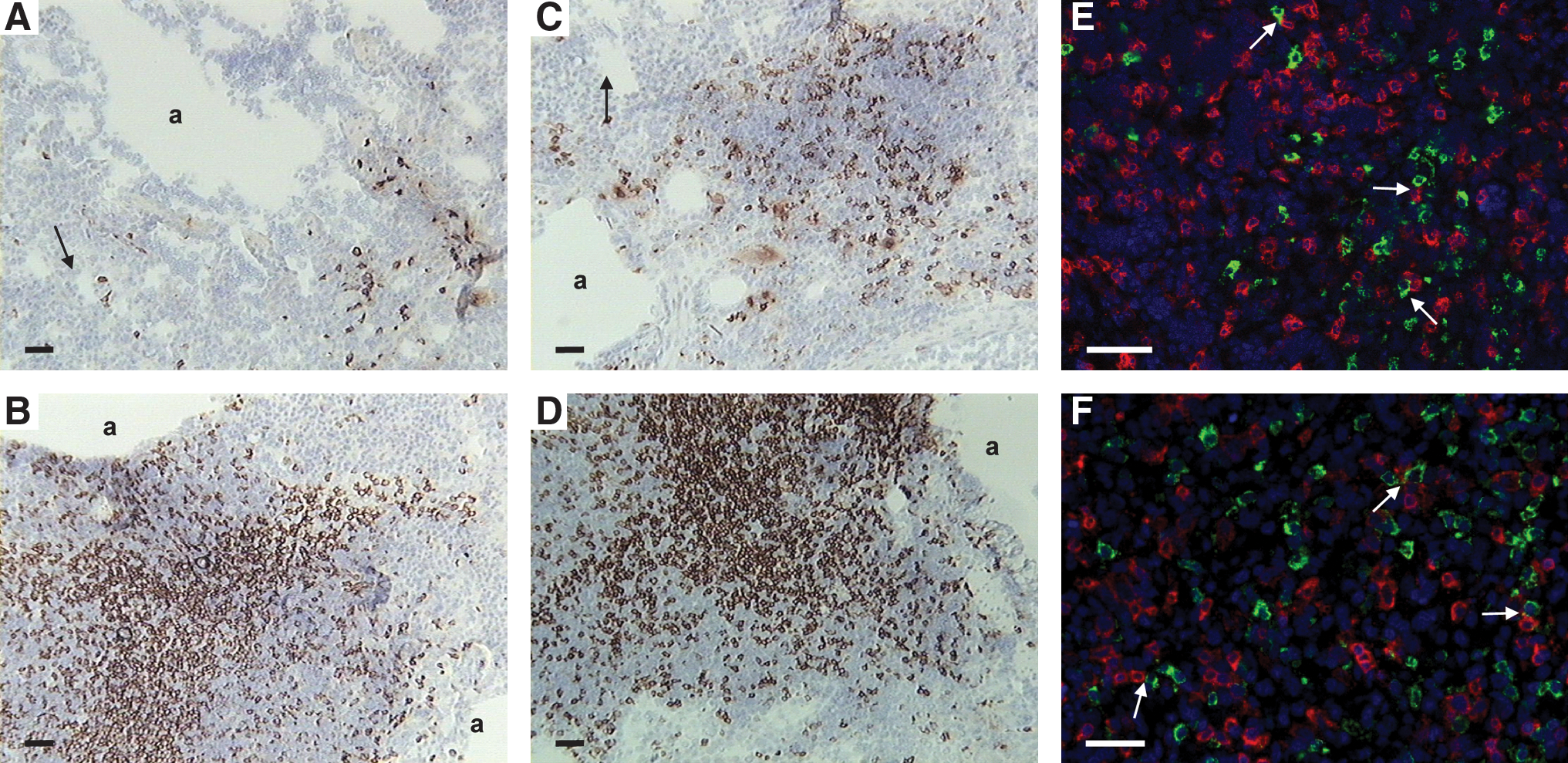
T-cell populations surround infected cells in the lung after intratracheal infection. Lung sections from resistant (
The spatial relationship between T cells and infected cells was investigated by immunofluorescence. No evidence of infected (pp38+) CD8 T cells was observed. Even at 5 d after insufflation no co-localization of the pp38+ and CD8+ cells could be distinguished (Fig. 5E). However, CD8+ T cells seemed to be surrounding the infected cells. A similar observation was made for CD4+ T cells (Fig. 5F); after extensive searches no infected cells could be distinguished, even during the highest levels of infection at 5 dpi in susceptible chickens.
Pulmonary B cells are primarily infected following MDV infection
Although crucial for pathogenesis, the cell types that take up MDV in the lung following natural infection remain unknown. A potential role of macrophages in the uptake and propagation of the virus has been proposed (1,8). In our experiments, macrophages were predominantly localized around the pulmonary spaces following intratracheal infection (Fig. 6A and B). The potential role of macrophages as carriers of MDV was tested by immunofluorescent double-staining. Viral pp38 antigen was observed in a small percentage of KUL-01+ macrophages in the lungs of susceptible chickens at 5 dpi, but this was by no means a frequent occurrence (Fig. 6D). Many non-macrophage-infected cells were present and were proposed to be B cells. Double staining of infected cells for the viral antigen pp38 and the B-cell marker Bu1 showed that B cells were the predominant infected cell type in the lung following infection (Fig. 6E and F). pp38-Specific staining of the infected cells was predominantly cytoplasmic, near the plasma membrane, and could be detected as early as 3 dpi.
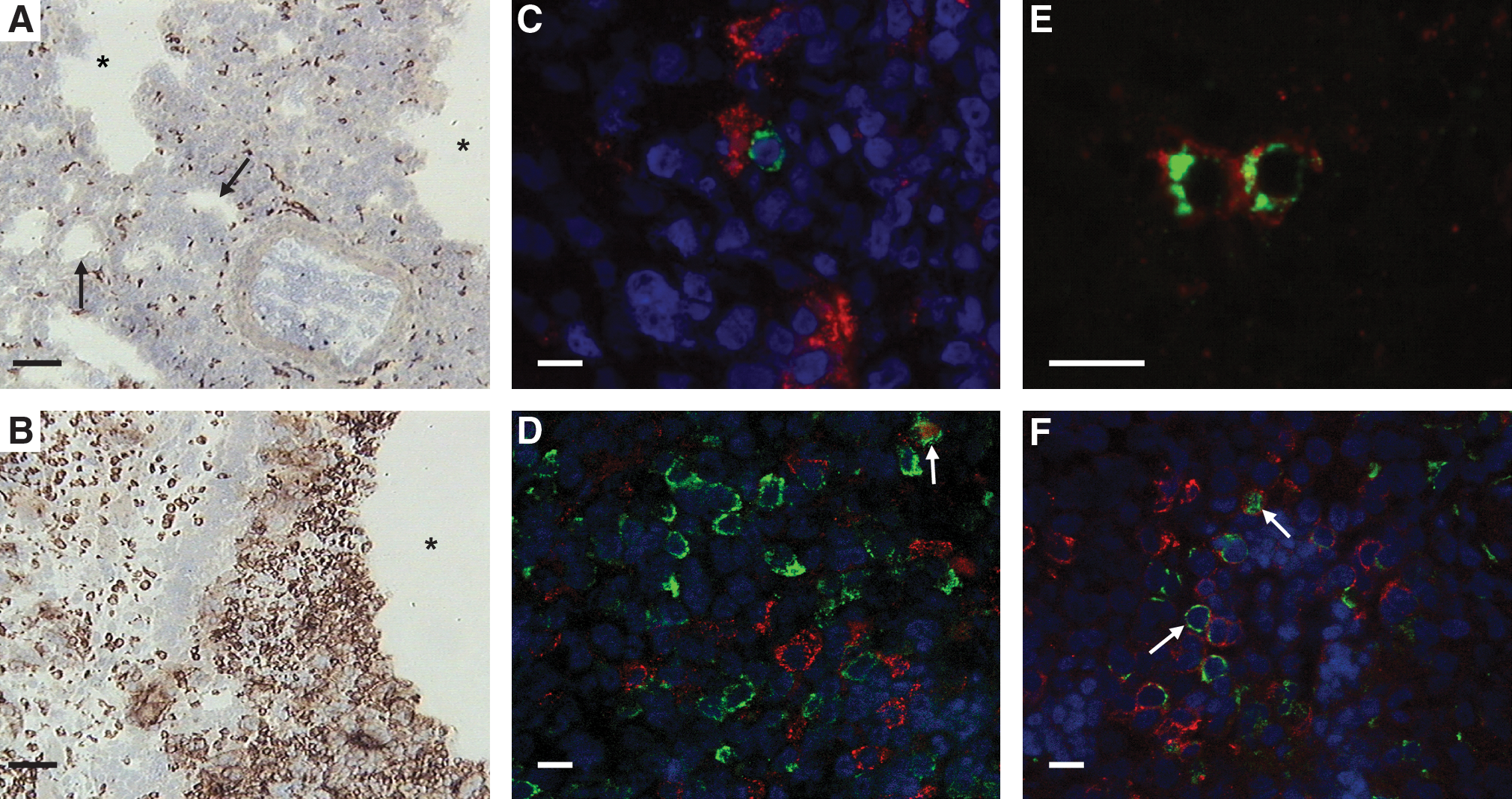
Viral replication in pulmonary B cells and macrophages of chickens after infection. The effect of intratracheal infection on macrophages in the lung was assessed by immunohistology. Lungs were removed from control (
The role of B cells in the pathogenesis after natural infection was investigated by depleting all B cells using embryonic surgical bursectomies. Resistant and susceptible chicks were bursectomized at embryonic day 18 before the efflux of B cells into the periphery (reviewed in 53). Control fertilized eggs were set under similar conditions. All chicks were allowed to hatch and were infected by the intratracheal route at 2 wk of age. Bursectomized susceptible line 7 chickens had a similar onset of clinical signs as intact resistant line 6 chickens (Fig. 7A). The resistant line 6 chickens that had been bursectomized were completely protected from any clinical signs for the duration of the experiment. These data were complemented by reduced viremia in the bursectomized chickens compared to their respective controls (Fig. 7B). In the susceptible line 7 chickens viremia was delayed.

Depletion of B cells delays viremia and clinical symptoms after intratracheal infection. Resistant and susceptible chickens bursectomized (Bx) at day 18 of embryonation or intact were infected at 2 wk of age. (
To identify how B cell depletion affected pathogenesis, B cells, macrophages, and CD4 and CD8 T cells were sorted from spleens of chickens infected by the natural route at 5 and 20 dpi (Fig. 7C and D). MDV replication was demonstrated in B cells and macrophages at 5 dpi in susceptible but not resistant chickens, and the cell types had very similar viral loads per cell. No viral replication was observed in T cells at this time point. Depletion of B cells by bursectomy prevented viral replication in KUL-01+ macrophages of susceptible chickens. At 20 dpi, predominantly CD4 and CD8 T cells were infected in the bursectomized susceptible chickens. No intact susceptible line 7 chickens remained for comparison at this time point. Intact resistant chickens showed only very low levels of viral replication in CD4 T cells, but did not have any detectable levels of viral replication once bursectomized.
Discussion
MD, a lymphoproliferative disease of chickens, is caused by a virus that is mainly spread by contact exposure. The ability to infect chickens via the respiratory tract has been demonstrated by studies performed as early as the 1970s; infection through contact exposure with airborne suspensions of infective dander (1,9,15,31,32) and with infected individuals (34,39,48), administration of feather tip extracts (16,21), as well as cell-associated virus (38) were all shown to cause MD. However, some characteristics of these approaches, such as reproducibility issues, inability to deliver a quantifiable dose of virus, or delivery to the gastrointestinal tract (1,9,29), precluded their use to investigate early events after infection. The approach to deliver infective dander via the trachea described here circumvents some of these issues, allowing for the exact timing of events following pulmonary infection. The microaerosolizer provides a simple and effective tool to introduce a standard dose of MDV in its native form into the trachea in a timed manner. We previously demonstrated the importance of the route and the form of the virus when parenteral infection with cell-associated virus was compared to intratracheal infection with native MDV for resistant and susceptible strains (14). Although 2.5 mg of dander contained 16 times more viral DNA compared to 1000 pfu of cell-associated virus (the doses used in the experiments), even increasing the infective dose of cell-associated virus 100 times could not replicate the timing of the clinical signs seen after intratracheal infection (14). In addition, preliminary data suggest that the form of the infective material is also crucial in determining clinical outcome (Staines and Butter, unpublished data). These data highlight the importance of the route of infection for pathogenesis and eventual clinical outcome.
Immune responses in line 6 and 7 genotypes have been investigated by several researchers and were shown to influence the outcome of infection (reviewed in 38). Here, the immune response after natural infection was investigated by quantitative analysis of cytokine transcript levels and cell subpopulations. We show that differences between genetically inbred strains that are susceptible or resistant to MD tumor formation may already be expressed during the early virological events in the pulmonary environment, and could therefore direct the ensuing pathogenesis. Viral replication could be detected as early as 2 dpi in the lung leukocytes of susceptible chickens, whereas that in the spleen was delayed by 1 d. Although cell-associated MDV was reported by Philips and Biggs (41) at 24 h following natural exposure infection, the length of contact with the infected chickens (3 d) did not allow for the determination of the precise time of infection (e.g., 24 h after the end of exposure could actually be 4 d after initial infection). The 1-d delay for detection of viral replication in the spleen could be a consequence of the transport of the virus from its site of entry. Although it is not clear if transport occurs through blood or lymph, Adldinger and Calnek (1) reported virus in the buffy coat of infected chickens as early as 18 h after infection. With a limited sensitivity of detection, these authors failed to identify any early replication in the lung. In spleen and lung leukocytes of resistant birds the 1-d delay of detection and significantly reduced viral replication at 5 dpi suggests that resistance is expressed at the replication level at this early stage. A correlation between viral load in the blood and tumor formation in line 6 and 7 chickens has been reported, although this was much later, at 14 dpi (12). That genetic resistance manifests itself so early with respect to viral load is remarkable, and the delay and lower viral load could be a distinct advantage to the resistant line 6, as viral and tumor load are correlated. The reduced viral replication could allow for more time to generate an effective immune response. For example, MDV-induced IFN-γ in lung leukocytes of resistant line 6 chickens is concomitant with viral replication, but delayed in those of susceptible chickens. The 2-d delay between viral replication and IFN-γ production in lung leukocytes of susceptible birds could provide a crucial time period for the virus to replicate uncontrolled, as IFN-γ is a potent inhibitor of MDV replication (22,56). It is unclear what cell population produces the IFN-γ in the resistant chickens; the influx of CD8 cells, potent secretors of IFN-γ, did not occur until 4 dpi. NK cells are a likely candidate and could potentially express IFN-γ. The mapping of MD resistance in line 6 and 7 chickens to the MDV1 locus (11), which shows synteny to the Cmv1 locus that controls susceptibility to infection with mouse cytomegalovirus and maps to the mouse NK gene complex (44), suggests a link between genetic resistance and NK cell activity. The importance of NK cells in MD pathogenesis was inferred from genetically resistant chickens that have earlier and stronger NK cell response in resistant (B21B21) compared to susceptible (B19B19) chickens (26,28,49). However, these cells were not examined in our experiments.
In the spleen, IFN-γ levels correlated with the onset of detectable viral replication and were higher in leukocytes of susceptible than resistant chickens at 4 and 5 dpi. Temporal expression studies in resistant chickens (B21B21) have revealed that IFN-γ transcription was upregulated in spleen extracts as early as 3 dpi following intra-abdominal infection with cell-associated virus (55). Similar to our data and those of Xing and Schat (55), Kaiser et al. (33) reported strong IFN-γ expression during the first week of viral replication in splenocytes of chickens with the resistant B21B21 or B2B2 haplotype. However, the delay in IFN-γ expression observed in susceptible B2B2 or B19B19 chickens in our experiments was not evident. The viral load between line 6 and 7 splenocytes was also not significantly different in the very first week and at odds with the data reported here. Both could be a result of the infection route.
The IFN-γ response in the spleen leukocytes corresponded with an overall increase in the levels of IFN-α, IFN-β, IL-6, and IL-18 inflammatory cytokine transcripts 4 and 5 days after intratracheal infection in susceptible birds. Comparable changes for IL-6 and IL-18 transcript levels between line 6 and 7 chickens have been described (33), although these were later in the infection. Consistent with earlier hypotheses (4,35), a strong increase in cytokines in the line 7 birds suggests a more profound inflammatory response, which potentially could attract more target lymphocytes. Interestingly, type I IFNs are upregulated in the spleens of susceptible chickens following infection. Hong and Sevoian (30), reported the opposite when using K and S strain chickens, and although a bioassay was used that could not discern between IFN-α, IFN-β, or IFN-γ, overall this was not expected, given the antiviral role of the IFNs.
Overall the differences we observed in the proportions of specific lymphocyte subpopulations in the spleen between the genotypes (more B cells and fewer T cells in the resistant chickens, independent of infection) were not observed in the lung. Thus a possible role for cell numbers in the lung to affect infection potential and influence genetic resistance, as postulated for the spleen (35), is unlikely. Infection caused an increase in CD3+CD8αβ+ T cells in the lungs of both susceptible and resistant chickens while viral replication was increasing. Although cytotoxicity was not addressed in this experiment, a cytotoxic T-cell population capable of lysing MDV-infected target cells is generated following MDV vaccination of line 6 chickens (35,42 and B.J.G. Baaten, unpublished data). A decrease in the CD3+CD8αβ+ T-cell population in the spleen coincided with extensive viral replication and is most likely a result of cytolytic infection.
We were unable to find infected T cells in the lung or spleen during the first 5 d. Flow cytometric approaches have shown that as early as 4 d after intraperitioneal infection T cells were pp38+ (7). The infection route may affect the cell tropism of the infection, and T cells may not become infected until after the 5 d investigated in our experiments. It has been postulated that phagocytic cells take up virus particles in the lung and transport these to the lymphoid organs, where infection of B and T cells occurs (1). Although infected pulmonary macrophages were identified, there were many KUL-01-negative virus-positive cells present, and thus macrophages were not the predominant infected cell type in the lung. Nevertheless, these do seem to be able to support viral replication, as sorted spleen macrophages from susceptible chickens were positive for MDV mRNA, confirming a report by Barrow et al. (8). However, since we show that a fully productive infection is already in progress in pulmonary B cells as early as 3 d after infection, when no infected macrophages can be identified, it seems unlikely that the macrophages are infected first. Although B cells have been shown to be the primary target cells in spleen following intraperitoneal infection with cell-associated virus, to our knowledge this is the first time that they have been shown to be infected in the lung after respiratory infection. Complete removal of B cells by surgical bursectomy before intratracheal infection greatly impacted pathogenesis, reducing viremia and clinical signs, especially in susceptible line 7 chickens. The substantial impact on pathogenesis is in accordance with results seen following parenteral infection with cell-associated virus (45). Further analysis of the cell types infected revealed that B cells and macrophages were infected, but that B-cell depletion did not only impact the B-cell compartment, but also prevented viral replication in macrophages of susceptible chickens. The absence of any detectable viral replication in sorted populations from line 6 spleens may be due to a lower level being undetected, and due to the timing of the event. Whereas secondary effects of bursectomy cannot be excluded, these data suggest that infection of B cells is required for macrophage infection, and provides more evidence for the B cells as the primary targets for MDV. These data confirm and extend earlier suggestions that MDV can replicate in macrophages (8), but challenge the paradigm that the macrophage's role in the pathogenesis is to transport MDV from the lung to the lymphoid organs, where the cytolytic phase ensues. Apparent differences between resistant and susceptible chickens do not seem to stem from a difference in the number of target B cells, as these were consistent over time. It is more likely that pulmonary B cells from line 7 chickens are more susceptible on a per-cell basis compared to those from resistant chickens, as reported by Lee et al. (35). It is not clear from our data whether B cells become infected directly, as is the case for other lymphotrophic herpesviruses (i.e., Epstein-Barr virus), or through an intermediate cell type, such as lung epithelium. Several researchers (23,41) have demonstrated that chicken lung epithelial cells themselves can harbor MDV antigen. Epithelial cells in the atria and infundibila have been shown to phagocytose small particles that enter parabronchi in other avian species (47). It is possible that epithelial cells become infected while translocating viral particles from the air-side of the lung to the parenchyma. Here, B cells could become infected directly by translocated viral particles, or indirectly through infected epithelial cells. Therefore genotypic variation at this level could account for the difference in viral replication demonstrated, and this is an area of great interest.
A better understanding of the early events following natural infection with MDV is crucial to investigate the possibilities of designing vaccines that block infection (reviewed in 2 and 27). Whereas current vaccines have been instrumental in controlling disease, they target viral replication. Challenge viruses still infect, replicate in, and are shed by MD-vaccinated chickens, providing a reservoir of MDV pathotypes that can survive vaccine-induced immune responses and are able to infect other chickens (10). Epidemiological modeling studies predict that vaccines that target pathogen replication tend to drive the pathogen to increased virulence (25). In contrast, vaccines that interfere at the infection stage, before amplification of the pathogen, may have a neutral or even negative effect on the evolution of virulence. Knowledge of the cellular and molecular events in the lung immediately following natural infection will aid in the eradication of the disease.
Conclusion
The data described here demonstrate that there is extensive viral replication in the lung within the first 5 d after intratracheal infection of susceptible chickens, which is complemented by an increase in inflammatory cytokines and the arrival of CD8 T cells. These events are delayed by 1 d in the resistant chickens, although interestingly, there is early increased and sustained IFN-γ expression in the lungs. Our findings suggest that pulmonary B cells sustain this early viral replication, whereas only a small number of infected macrophages can be identified.
Footnotes
Acknowledgments
This work was supported by DEFRA and the BBSRC. The authors would like to thank Dr. G.J. Underwood for help and guidance with the animal model, Dr. P. Barrow for bacteriological examination of dander samples, H. Davis for screening serum samples for IBDV and CIAV, Dr. D. Worth and J. Oldham for cell sorting, and Dr. P. Kaiser for generous donation of cytokine primers and probes.
Author Disclosure Statement
No competing financial interests exist.