
Abstract
In bovine Mx1, only an amino acid substitution between Ile and Met at position 120 was detected by the nucleotide sequence and mismatched PCR-RFLP technique. The Ile variant was assumed to distribute mainly in the bovine population since the gene frequency was 79.3%. Furthermore, we cloned water buffalo Mx1 cDNA, which showed 51 nucleotide and 20 amino acid substitutions in comparison with that of the cow. Another kind of Mx1 cDNA, bovine Mx1B cDNA, was found and it was deduced to cause 27 amino acid substitutions at the N-terminus compared to the original Mx1 by alternative splicing. However, no variation was detected in 27 amino acids specific for Mx1B among 29 cows and a water buffalo. We established four kinds of mRNA-expressing 3T3 cells and Vero cells. When infection experiments were performed using recombinant vesicular stomatitis virus (VSVΔG*-G), bovine Ile and Met types and water buffalo Mx1 mRNA-expressing cell lines showed equally positive antiviral activities (p < 0.05). On the other hand, bovine Mx1B mRNA-expressing cell lines did not have activity against VSVΔG*-G. Intracellular localization of bovine Mx1 and Mx1B proteins was examined by a transiently GFP-fused expression system in 3T3 cells. Bovine Mx1 was localized in the cytoplasm, while bovine Mx1B was mainly localized in the nucleus. An arginine-rich nuclear localization signal was found in 27 amino acids specific for Mx1B. N-terminus-deleted Mx1B was only localized in the cytoplasm, and the deleted Mx1B-expressing cell lines showed significantly positive antiviral activities (p < 0.05) against VSVΔG*-G.
Introduction
In our previous study, an Ile variant at amino acid position 120 (T at nucleotide position 458) in the bovine Mx1 protein was shown to be frequent in 11 cows, while a Met variant (G at this position) was observed in only one Hereford and one Brahman as a heterozygote (17). In that study, we compared the antiviral activities against recombinant vesicular stomatitis virus (VSVΔG*-G) of the Ile and Met variants at amino acid position 120 of bovine Mx1. Furthermore, during the course of that study, a splicing variant of bovine Mx1B (11), which codes 27 amino acid residues different from the 24 residues in the N-terminus corresponding to Mx1, was constantly detected in several breeds. Therefore, in the present study, we also compared the antiviral activities of bovine Mx1 and Mx1B. In addition, we cloned a water buffalo Mx1 cDNA in order to check the homology of amino acid sequences and the antiviral activity in related ruminant animals.
Materials and Methods
Breeds and DNA extraction
Genomic DNAs were obtained from 58 cows of 7 bovine breeds. Some genomic DNAs were kindly provided by Prof. T. Amano (Tokyo University of Agriculture, Kanagawa, Japan), Dr. E. Kobayashi (National Livestock Breeding Center, Fukushima, Japan), and Dr. N. Yamamoto (National Agricultural Research Center for the Western Region, Shimane, Japan). Otherwise, the DNAs of cows as well as a swamp water buffalo were extracted from blood samples in this study. Briefly, leukocytes were separated from the blood samples by centrifugation at 1800 rpm for 15 min, washed with 8 mL of phosphate-buffered saline (PBS), transferred to a tube containing 2 mL HISTOPAC-1077 (Sigma, St. Louis, MO), and centrifuged at 1800 rpm for 30 min. The obtained pellet of leukocytes was seeded on a 10-cm tissue culture dish (Falcon Labware, Franklin Lakes, NJ) containing 10 ml of RPMI 1640 medium (Sigma) supplemented with 10% fetal bovine serum (FBS), 5 μg/mL of phytohemagglutinin (Sigma), 1 μg/mL of pokeweed mitogen (Sigma), 100 IU/mL of penicillin (Nacalai Tesque, Kyoto, Japan), and 100 μg/mL of streptomycin (Meijiseika, Kyoto, Japan). The cultured leukocytes were incubated for 42 h at 37°C in 5% CO2. The DNA was extracted from the leukocytes by the phenol/chloroform method, and was finally dissolved in TE (10 mM Tris-HCl at pH 8.0 and 1 mM EDTA).
Mismatched polymerase chain reaction-restriction fragment length polymorphism
A mismatched forward primer of either Bomx428SspF (5′-AGGCAAAGTCAGCTTCCTGGACAAAAATAT-3′, 428–457) or Bomx428NlaF (5′-AGGCAAAGTCAGCTTCCTGGACAAAGACAT-3′, 428–457), and reverse primer BomxNTR (5′-AGACCCCATCTATTTCACCTTGTT-3′, intron 5) were constructed for PCR-RFLP to identify the nucleotide substitution encoding the amino acid variation at 120. Mismatched PCR was performed using GoTaq® DNA polymerase (Promega, Madison, WI) with a DNA Thermal Cycler (Perkin-Elmer, Waltham, MA). The cycling profile comprised an initial denaturing step at 94°C for 5 min, followed by 5 cycles at 94°C for 30 sec, 53°C for 3 min, and 72°C for 30 sec, and 30 cycles at 94°C for 30 sec, 59°C for 1 min, and 72°C for 30 sec. The PCR products obtained using the Bomx428SspF and BomxNTR primers were digested with restriction endonuclease SspI (Toyobo, Osaka, Japan) at 37°C for 2 h, and those obtained using Bomx428NlaF and BomxNTR were digested with NlaIII (Toyobo) at 37°C for 2 h. The digested DNA fragments were electrophoresed through 3% agarose gel and visualized with ethidium bromide.
RNA extraction and reverse transcription-polymerase chain reaction
Leukocytes were separated from blood samples obtained from 8 animals, including two Holstein cows, and one cow each of the breeds Jersey, Ayrshire, Charolai, Limousine, and Japanese brown, and one water buffalo. The cultured leukocytes and cultured Mardin-Darby bovine kidney (MDBK) cells were incubated for 42 h at 37°C in 5% CO2, and then treated with 1000 U/mL of recombinant human interferon-α-2b (IFN) (Intron-A; Schering-Plough, Kenilwoth, NJ) for 12 h to stimulate Mx mRNA expression. Subsequently, total RNA was extracted using ISOGEN (Nippongene, Tokyo, Japan), and the first double-stranded cDNA was synthesized from the total RNA by reverse transcription (RT) using a ReverTra-Ace kit (Toyobo). Primers used for amplification of the bovine Mx1 cDNA were designed from the nucleotide sequence reported by Ellinwood et al. (5). A 2280-bp fragment, which contained almost the full-length Mx1 cDNA, was generated using the primers Bomx1F (5′-GCCAACTAGTCAGCACTACATTGTC-3′, 44–68) and Bomx5R (5′-ATGGGGTGTGAAAGGCTATT, 2343–2324) as previously described (17). A 2404-bp fragment of the full-length Mx1B cDNA was also amplified using the primer BomxB1F (5′-TGGGTGGTTACTCCTCCCTTTAG-3′, 283–305), which was designed on the basis of the nucleotide sequence reported by Kojima et al. (11) (GenBank accession no. AB060169), and the primer Bomx5R. PCR was carried out using KOD DNA polymerase (Toyobo) on a DNA Thermal Cycler (Perkin-Elmer). The cycling profile consisted of an initial denaturing step at 94°C for 5 min followed by 33 cycles at 94°C for 1 min, 58°C for 1 min, 68°C for 1 min, and a final extension step of 68°C for 5 min. The RT-PCR products of Mx1 and Mx1B cDNAs were cloned into pGEM-T Easy vector (Promega). The nucleotide sequences of bovine Mx1 cDNAs were determined using an ABI310 sequencer (Applied Biosystems, Foster City, CA).
Construction of expression vector and transfection into 3T3 cells
The 2280-bp fragment of Mx1 cDNA and the 2404-bp fragment of Mx1B cDNA from the pGEM-T vector of a Holstein cow and a water buffalo were religated into the NotI site of the pCI-neo expression vector (Promega), which contains the human cytomegalovirus immediate-early enhancer/promoter and the neomycin phosphotransferase gene. Embryonic 3T3 fibroblastic cells from a BALB/c mouse (Riken Cell Bank, Tsukuba, Japan) and Vero cells from an African green monkey kidney (Riken Cell Bank) were grown in DMEM containing 10% fetal bovine serum. The Mx1 and Mx1B constructs were transfected into 3T3 cells using FuGENE-6 transfection reagent (Roche Molecular Biochemicals, Basel, Switzerland) in accordance with the manufacturer's instructions, and the transfected clones were selected in medium containing 500 μg/mL of Geneticine (G418; Gibco-BRL, Carlsbad, CA). Individual clones of stably transfected cells expressing the Mx1 or Mx1B mRNA were tested by RT-PCR using the gene-specific primers described previously (24). Mouse β-actin mRNA was used as an internal control in the RT-PCR using primers mbActinF (5′-TGGAATCCTGTGGCATC-3′, 886–910) and mbActinF (5′-TAAAACGCAGCTCAGTAACAGTCCG-3′, 1234–1210). Mx1BΔ25, a construct of the pCI-neo vector of which 75 nucleotides coding the first 25 amino acids in the 5′-end specific for Mx1B were artificially deleted from 2404 bp of Mx1B cDNA, was made by RT-PCR using the primers Bomx1B25F (5′-ATGGCATCACTGACTCGATG-3′, 524–543), Bomx5R, and 2404 bp of Mx1B cDNA from the pGEM-T vector as a template.
Measurement of antiviral activity
Recombinant VSVΔG*-G carrying the green fluorescent protein (GFP) gene instead of the G protein gene was provided by Prof. A. Takada (Research Center for Zoonosis Control, Hokkaido University, Hokkaido, Japan). Stably transfected clones in 3T3 cells expressing bovine or water buffalo Mx1 mRNAs were subjected to experimental infection with VSVΔG*-G. Infectivity of VSVΔG*-G in 3T3 clones was determined by counting the number of GFP-expressing cells in 10–20 microscopic fields (10,17). At least three independent experiments were carried out for each clone.
Intracellular localization
For observation of intracellular localization of the bovine Mx1 and Mx1B proteins, we constructed GFP-fusion protein-expressing vectors using a pEGFP-N1 vector (Clontech, Mountain View, CA), which includes the GFP gene at the 3′-end. Primers containing recognition sites of some restriction enzymes (NheI, GCTAGC; XhoI, CTCGAG) were produced as follows: Bomx1NheF (5′-GTCAAGGCTAGCGAAGGTATATTGAGGAAGAAG-3′, 78–98), Bomx1BNheF (5′-CACCTCGCTAGCAGAGTTGACTCACTGGAAAAG-3′, 439–459), Bomx1BNhe25F (5′-ATGAGAGCTAGCGATGGCATCACTGACTCGATG-3′, 523–543), Bomx1XhoR (5′-AGGGCGCTCGAGGCCCGGGAACTTGGCCAGCC-3′, 2042–2023), and Bomx1BXho25R (5′-AATCTCCTCGAGCGAGTCAGTGATGCCATCCA-3′, 540–521). Then PCR was carried out using DNA from recombinant pCI-neo vectors with the 2280-bp fragment of Mx1 cDNA, or the 2404-bp fragment of Mx1B cDNA. The PCR products were adequately digested by NheI or XhoI, and religated into the pEGFP-N1 vector.
Embryonic 3T3 fibroblast cells were transiently transfected by using FuGENE-6 transfection reagent with each Mx1 cDNA construct into the pEGFP-N1 vector. In the construct, the GFP gene is connected with a carboxyl terminal of each Mx1 cDNA. 3T3 cultured cells were seeded at a density of about 2 × 104 cells/mL on a 24-well plate and allowed to reach 70–80% confluence. Then 1 μg of each plasmid was transfected into each cultured cell in the plate according to the manufacturer's instructions. Two days after transfection, intracellular localization of the Mx1 protein was performed by observation of GFP fluorescence in the transient cells through an appropriate filter using an Olympus IX 70-23 FL/DIC microscope (Olympus, Tokyo, Japan).
Results
Genetic variant causing an amino acid substitution in Mx1
An amino acid substitution was detected only at position 120 in bovine Mx1 protein (17). Ile and Met variations are derived from a nucleotide substitution between T and G at nucleotide position 458 of the Mx1 cDNA, respectively. In this study, we first aimed to distinguish this nucleotide substitution by PCR-RFLP using a mismatched forward primer of either Bomx428SspF or Bomx428NlaF, and reverse primer BomxNTR. A typical example of PCR-RFLP in the case of restriction enzyme SspI is shown in Fig. 1. Since the size of the PCR product is 105 bp when using Bomx1SspF and BomxNTR, the PCR product in an Ile variant corresponding to T at nucleotide position 458 was digested into both fragments of 77 bp and 28 bp, but a Met variant corresponding to G was not digested as a 105-bp fragment. Using this PCR-RFLP technique, bovine Mx1 alleles were examined in 58 cows from 7 breeds (Table 1), and the Ile variant was assumed to be distributed mainly in the bovine population because the gene frequency was 79.3%, whereas the frequency of the Met variant was estimated to only be 20.7%. We obtained exactly the same results of genotypes using primers of Bomx428NlaF and BomaxNTR and restriction enzyme NlaI, as well as those using primers of Bomx428SspF and BomxNTR and SspI.
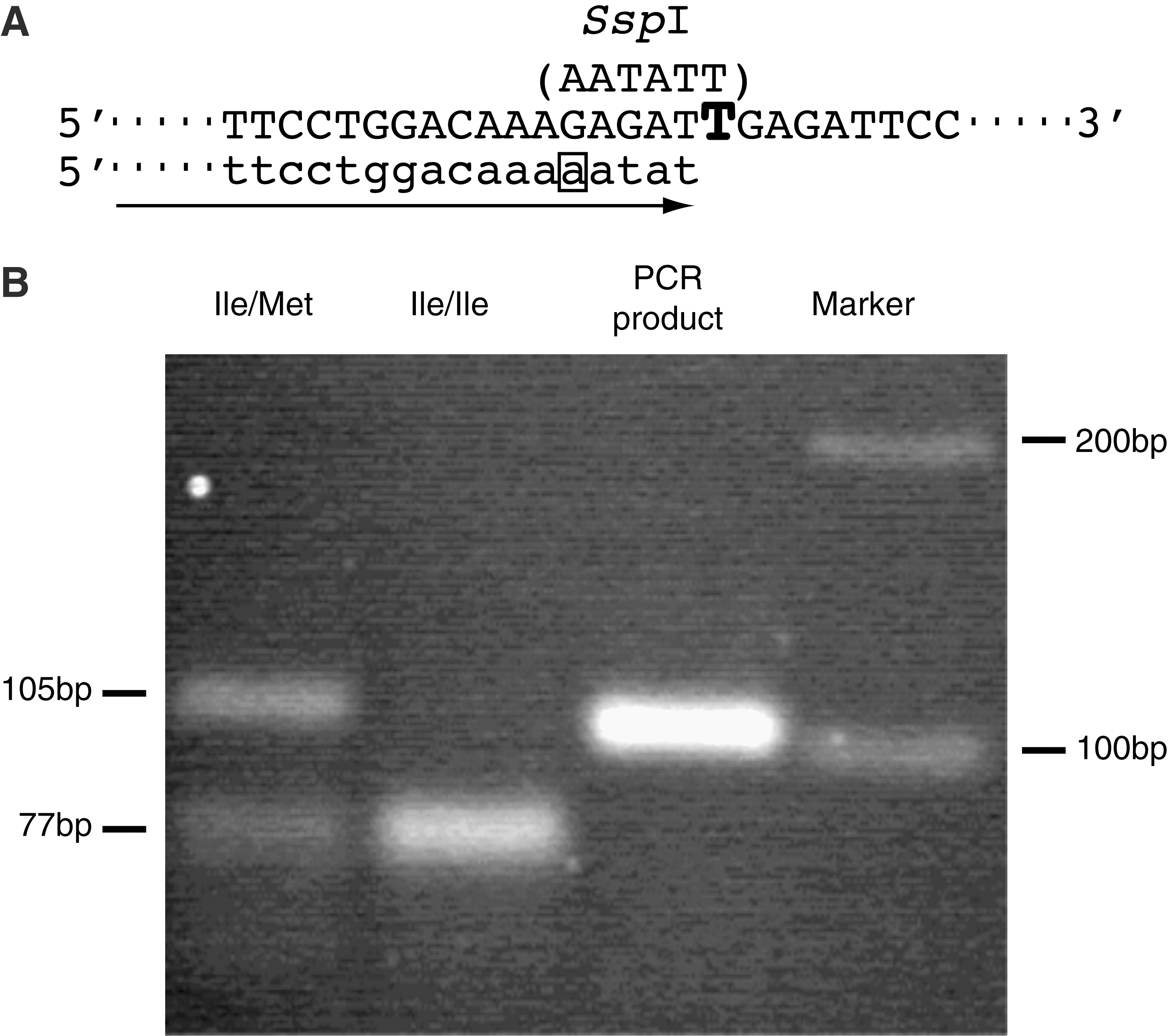
(
Splicing variant causing amino acid substitutions at the 5′-end in Mx1
When the nucleotide sequences of 2280-bp fragments of bovine Mx1 cDNAs from 7 cows, including two Holstein cows, and one cow of each breed of Jersey, Ayrshire, Charolai, Limousine, and Japanese brown, as well as previous data for 11 cows of two Holstein, three Japanese black, three Hereford, one Angus and two Brahman (17) were determined, there was no amino acid variation among the 18 cows except at position 120. However, another kind of Mx1 cDNA, Mx1B, that had a distinctly different sequence with 546 nucleotides in the 5′ end, was constantly observed by RT-PCR in all cultured leukocytes and a MDBK cell line using primers Bomx1BF and Bomx5R designed for Mx1B. This different type of Mx1 was deduced to cause 27 amino acid substitutions from 24 amino acids of the original Mx1, and had been reported as a splicing variant Mx1B found in a cDNA library from the endometrial tissue of a pregnant cow by Kojima et al. (11). In this study, no variation was detected in the 27 amino acid residues specific for Mx1B among the 29 cows examined.
Subsequently, we cloned Mx1 and Mx1B cDNAs from a water buffalo and determined the nucleotide sequences (GenBank accession no. AB462524). When the nucleotide sequence of Mx1 cDNA of a water buffalo was compared with that of a Holstein cow, 51 substitutions were found, and 20 amino acids were exchanged between them (Fig. 2). However, in this case also, no variation was observed in the 27 amino acids specific for Mx1B among the water buffalo and cows.
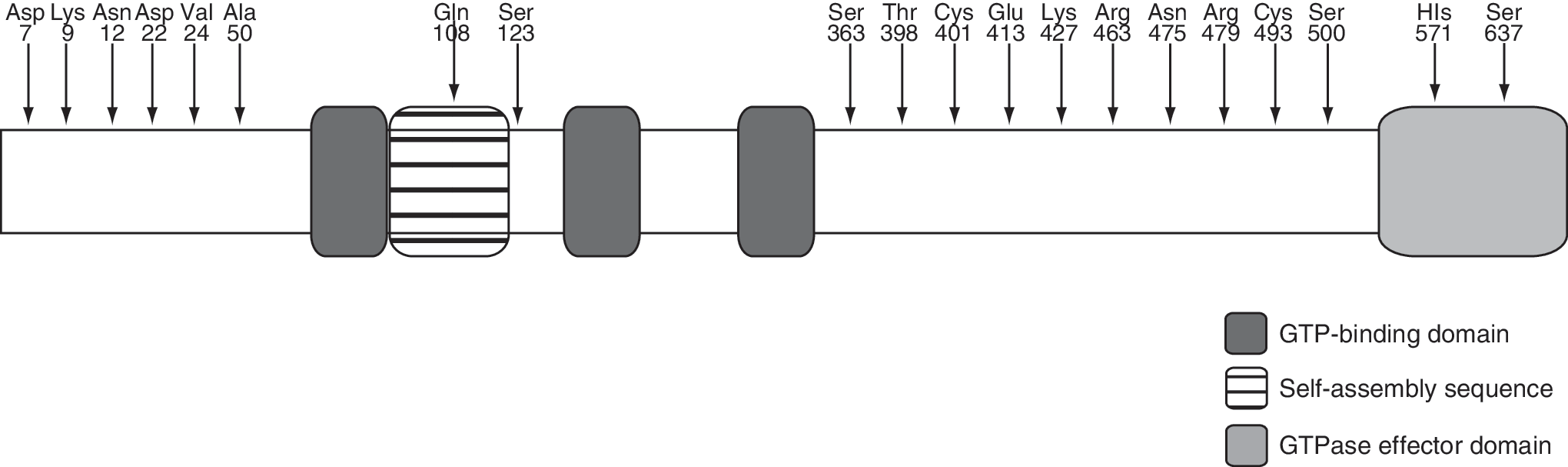
A schematic diagram showing the amino acid exchanges detected in water buffalo Mx1 (GenBank accession no. BAG72219) protein in comparison with bovine Mx1 protein (GenBank accession no. AAC18655). The amino acids indicate substituted amino acids, and numbers denote the amino acid positions exchanged.
Antiviral activities of genetic and splicing variants of Mx1
Four kinds of cDNAs, bovine Mx1 cDNAs from Ile and Met types at amino acid position 120, a splicing variant of bovine Mx1B cDNA, and a water buffalo Mx1 cDNA, were transfected into 3T3 cells, and permanent mRNA-expressing cell lines were cloned. Infection experiments were performed in the mRNA-expressing cell lines using VSVΔG*-G to clarify the antiviral activities. Parental 3T3 cells and empty pCI-neo vector-transfected 3T3 cells were used as negative controls. The 3T3 cells established from embryonic cells of BALB/c mice carry non-functional Mx genes because the deletion types of alleles have been found in the laboratory mice (6,7). As shown in Fig. 3A, Ile and Met types of bovine Mx1 mRNA-expressing cell lines and water buffalo Mx1 mRNA-expressing cell lines showed significantly positive antiviral activities (p < 0.05) against VSVΔG*-G compared to the activities of both negative controls. On the other hand, bovine Mx1B mRNA-expressing cell lines did not show antiviral activities against VSVΔG*-G, as well as controls. We repeated the same infection experiment in the mRNA-expressing cell lines transfected into Vero cells, as well as into 3T3 cells. Completely identical results were obtained for transfected Vero cell lines (data not shown).
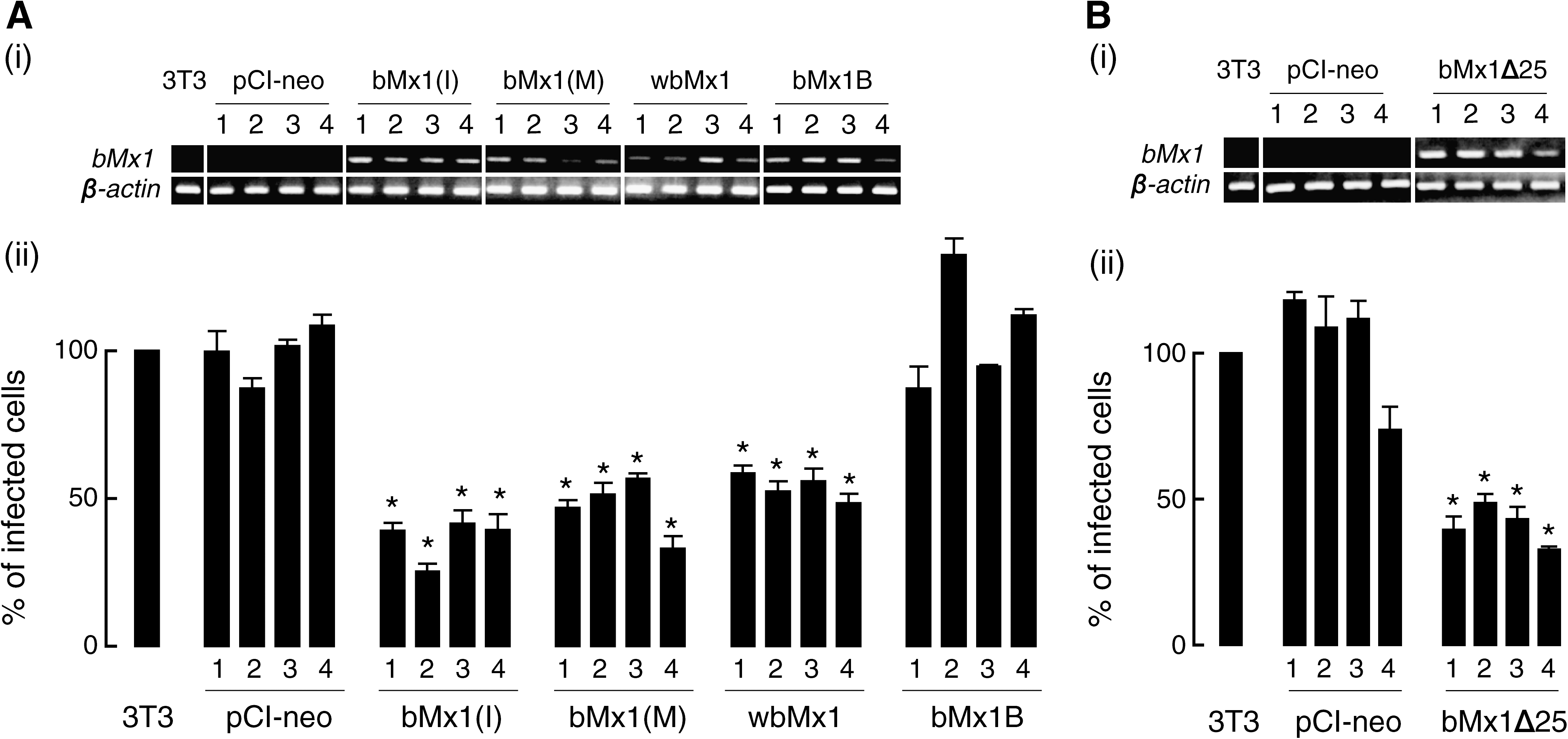
Bovine Mx1 mRNA and β-actin expression levels by RT-PCR in each clone (i) and infectivity of VSVΔG*-G (ii). Abbreviations: (
Subcellular localization of bovine Mx1 and Mx1B
Intracellular localization of the bovine Mx1 and Mx1B proteins was examined using each Mx1 cDNA construct into the pEGFP-N1 vector in transiently transfected 3T3 cells. In a control experiment, GFP fluorescence in the transfected cells with only the pEGFP-N1 vector was found in both the nucleus and cytoplasm (Fig. 4A). Subsequently, the fluorescence of transfected cells expressing GFP-fused bovine Mx1 was observed in the cytoplasm with a granular-like pattern (Fig. 4B), while that of transfected cells expressing GFP-fused bovine Mx1B was detected mainly in the nucleus with a granular-like pattern, and slightly in the cytoplasm with a diffused pattern (Fig. 4C). At that time, we noticed that 27 amino acids specific for Mx1B contained an arginine-rich sequence; the arginine-rich sequence was similar to a nuclear localization signal in Rev protein of human immunodeficiency virus (HIV) (23), as shown in Fig. 5B. Then we made a pEGFP-N1 construct of Mx1B(1–25) including only 75 nucleotides coding the first 25 amino acids specific for Mx1B, and the fluorescence of transfected cells expressing GFP-fused Mx1B(1–25) was clearly observed in the nucleus with a granular-like pattern (Fig. 4D), while that of transfected cells expressing GFP-fused Mx1BΔ25, which is a construct with deletion of the 75 nucleotides specific for Mx1B, was observed only in the cytoplasm with a granular-like pattern (Fig. 4E).
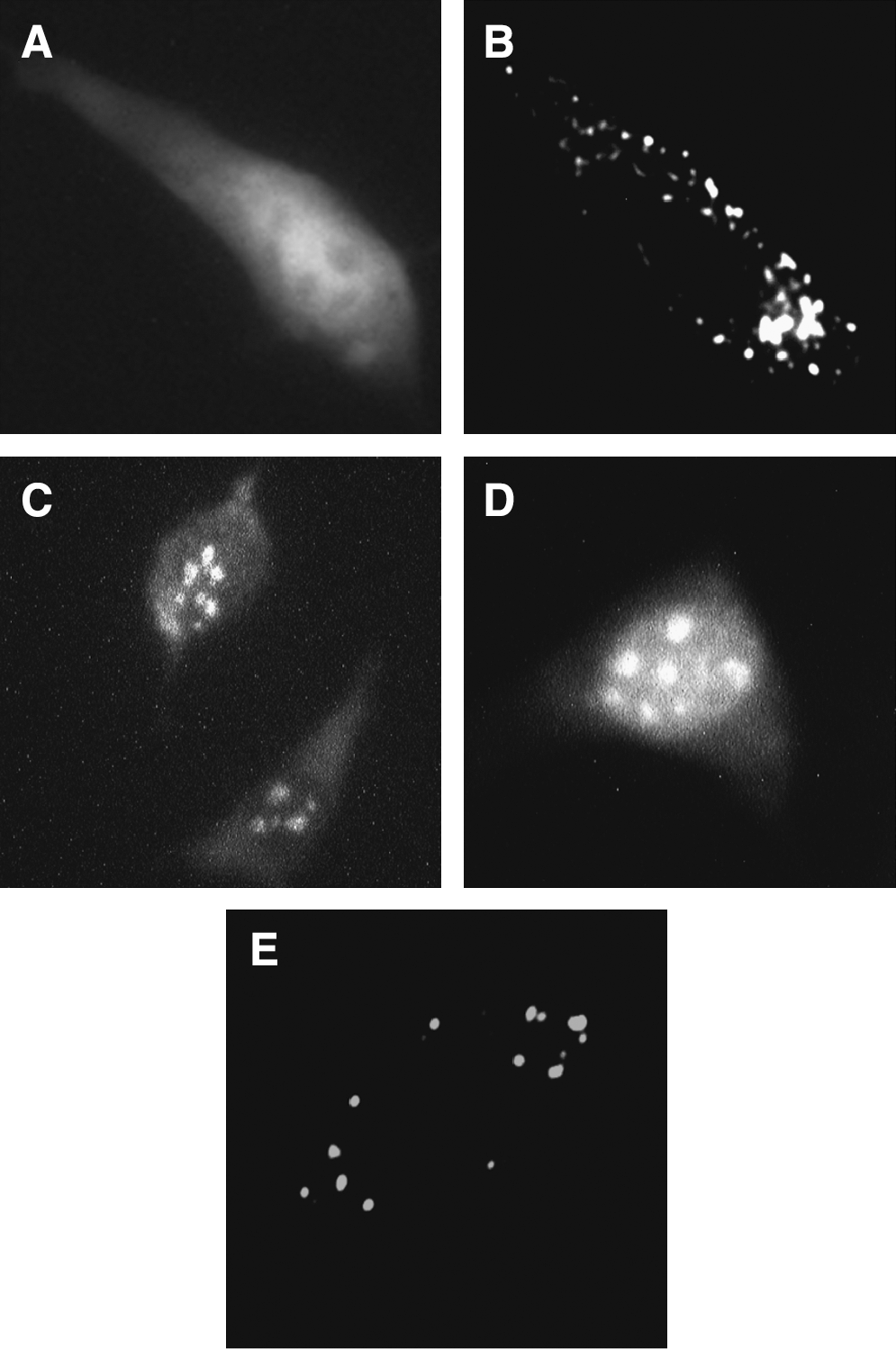
Intracellular localization of GFP-fused bovine Mx1. (
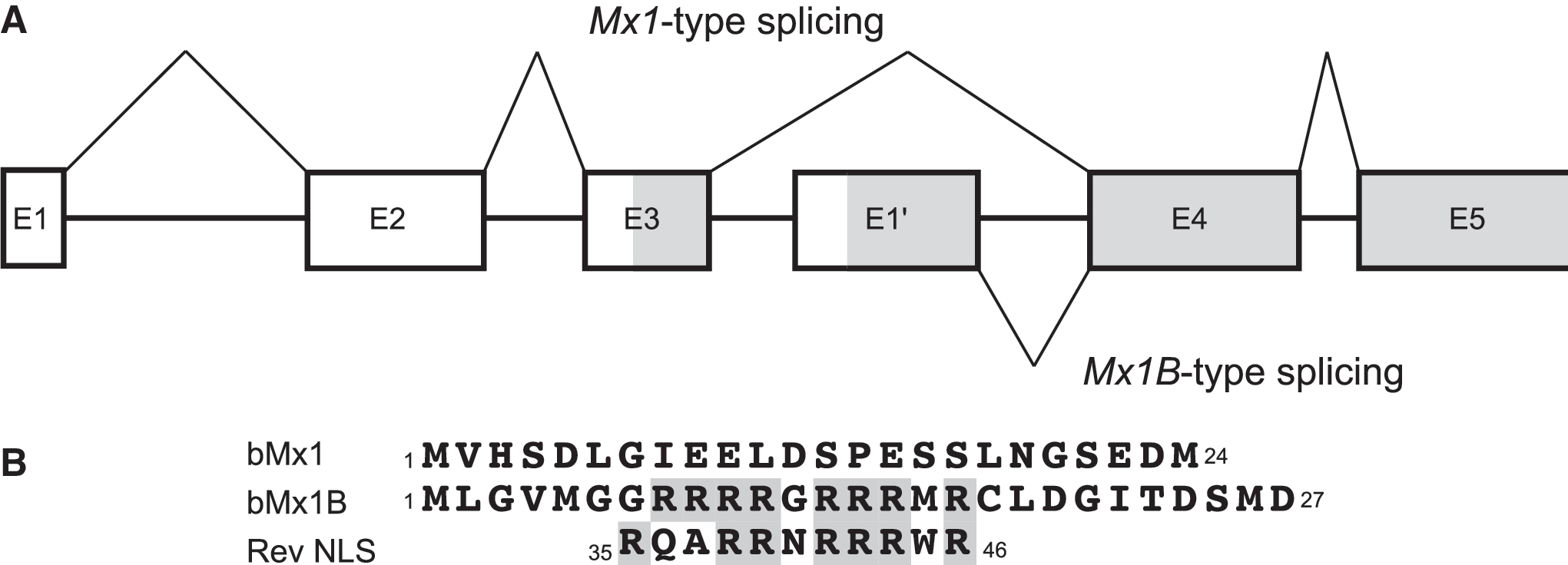
Schematic representation of bovine Mx1 genomic structure. (
Antiviral activity of bovine Mx1BΔ25
Mx1BΔ25 cDNA construct with a pCI-neo vector was also transfected into 3T3 cells, and stable mRNA-expressing cell lines were cloned. When an infection experiment was performed in the transfected cells using VSVΔG*-G, the mRNA-expressing cells exhibited significantly positive antiviral activities (p < 0.05) as shown in Fig. 3B.
Discussion
The results of this study and those of a previous study by Nakatsu and others (17) indicate that the amino acid sequence of bovine Mx1 protein is highly conserved, since only one amino acid substitution between Ile and Met at position 120 was detected in a total of 18 cows of 10 breeds. Furthermore, the Mx1 allele coding the Ile variant was assumed to be distributed mainly in the bovine population, based on the results of a PCR-RFLP test (Table 1). However, both Ile and Met types of bovine Mx1 mRNA-expressing cell lines, as well as water buffalo Mx1 mRNA-expressing cell lines, equally exhibited positive antiviral activities against VSVΔG*-G (Fig. 3A).
A water buffalo Mx1 cDNA was cloned for the first time in this study and the nucleotide sequence was determined. When Mx1 cDNA from the water buffalo was compared to that from a bovine breed, 51 nucleotide substitutions and 20 deduced amino acid exchanges were found (Fig. 2), and this level of difference was similar to that of Mx2 cDNA, in which 46 nucleotide substitutions and 12 deduced amino acid exchanges were found between them (3).
Of interest, a splicing variant, Mx1B (10), which is only different in 27 amino acids at the N-terminus from 24 amino acids of the original Mx1, was expressed constantly in leukocytes from all of the above 18 cows of 10 breeds. Furthermore, we found bovine Mx1B mRNA expression in many tissues by RT-PCR (unpublished data). In this study, no variation was detected in the 27 amino acids specific for Mx1B in any of the 29 cows and water buffalo examined. However, Mx1B mRNA-expressing cell lines did not show antiviral activity against VSVΔG*-G. We noticed that 27 amino acid residues specific for Mx1B contained an arginine-rich sequence that was similar to a nuclear localization signal in Rev protein of HIV (23) (Fig. 5). VSV is a virus that multiplies in the cytoplasm, and we considered this to be a reason why Mx1B mRNA-expressing cells did not show antiviral activity against VSVΔG*-G with different intracellular localization for the host-and-pathogen relationship. The results of this study showed not only that bovine Mx1B is localized mainly in the nucleus (Fig. 4C), but also that the Mx1B-specific sequence is a nuclear localization signal (Fig. 4D). Furthermore, Mx1BΔ25, a mutant with deletion of the first 25 amino acids specific for Mx1B, shifted localization to the cytoplasm, and the mRNA-expressing cells clearly acquired positive antiviral activity against VSVΔG*-G (Fig. 3B). We now have a plan to determine whether bovine Mx1B has antiviral activity against viruses replicating in the nucleus.
Mx1 and Mx2 proteins have been found in the mouse, and it has been shown that Mx1 localizes in the nucleus and inhibits influenza virus replication in the nucleus (2), whereas Mx2 localizes in the cytoplasm and blocks VSV (7) and hantavirus (8) multiplication in the cytoplasm. Intracellular localization of Mx proteins and the place where viruses multiply must be correlated. Rodent Mx1 as well as bovine Mx1B localize in the nucleus with a granular-like pattern. However, a nuclear localization signal of bovine Mx1B is found in the N-terminus, and that of mouse Mx1 exists in the C-terminus. Functional Mx proteins have been found to localize in the cytoplasm of mammals, including humans (20), mice (7), dogs (16), and pigs (15,19). Furthermore, high-level expression of Mx1 protein correlated with IFN-τ has been observed in the endometrium of pregnant sheep (4,18), and sheep and bovine are closed species. In this respect, the intracellular localization and immune defense mechanism of bovine Mx1 and Mx1B might be more specialized than those of other mammalian Mx proteins.
Footnotes
Acknowledgments
Blood samples from various bovine breeds were obtained from the National Livestock Breeding Center, Tokachi Station, Otohuke, Japan, and the blood sample from the water buffalo was obtained from Japan Heidi Farm, Naganuma, Hokkaido, Japan. We would like to thank their staff members for giving us the samples.
Author Disclosure Statement
No conflicting financial interests exist.