
Abstract
Confirmed reports of large domesticated cats becoming infected with highly pathogenic avian influenza (HPAI) H5N1 virus have raised questions about both the risk of infection for these animals, and their potential as vector or reservoir hosts in an influenza pandemic. With this in mind, we examined the immunogenicity of the hemagglutinin (HA) of H5N1 strain A/Vietnam/1203/04 using several different vaccination strategies. Data from ELISA assays showed that vaccination with a single dose of recombinant H5 HA protein induces a robust antibody response against both whole inactivated virus and recombinant HA antigen. Moreover, a single dose of the recombinant H5 HA protein induced hemagglutination inhibition titers ≥40, which is indicative of protective immunization. Cats receiving the IND H5N1 vaccine required two doses before similar H5 HA-specific antibody titers were observed, and despite boosting, these animals had HIA titers that were lower than or equivalent to those in the group receiving one injection of recombinant protein. In contrast, cats vaccinated with plasmid DNA encoding HA failed to develop HA-specific antibody responses above those seen in cohorts receiving an unrelated control plasmid. The results of this study indicate that recombinant H5 HA protein-based vaccines can rapidly induce high serum antibody titers, and may be more effective than either inactivated influenza virus or DNA vaccines in cats.
Introduction
Cats were not initially thought to play a role in the natural transmission of H5N1 influenza (2). However, as concerns about cats facilitating adaptation of the virus for human-to-human transmission have grown, official guidelines for controlling the spread of the H5N1 influenza virus have also begun targeting these animals (10). The proposed measures include quarantine of cats residing within a 3-km radius of an avian influenza virus outbreak (4), recommendations regarding animal husbandry (7), and intentional culling of companion animals within the vicinity of virus outbreaks (1). The unofficial responses have been especially concerning, as hundreds of cats were reportedly abandoned in both Germany and France following reports of a single infected cat (15). The deaths of naturally infected cats, evidence of cat-to-cat and the potential for cat-to-human transmission, as well as the official and unofficial reactions to outbreaks, all indicate that an effective feline vaccine against H5N1 influenza virus is greatly needed.
The two main influenza virus vaccines available in the United States are a trivalent inactivated influenza vaccine (TIV), and live attenuated influenza vaccine (LAIV). Both are made from vaccine virus strains grown in embryonated chicken eggs. In the event of an avian influenza pandemic, vaccine manufacturers will not have the resources to produce such vaccines for both humans and companion animals. Therefore alternative vaccines and vaccination strategies need to be developed to protect pets and reduce the possibility of zoonotic transmission of influenza virus. In this report we tested the immunogenicity of H5N1 hemagglutinin (HA) using several different vaccination strategies. Our results indicate that recombinant H5 HA protein vaccines are likely to be more effective than either inactivated whole influenza virus or DNA vaccines in cats.
Materials and Methods
Viruses
The virus used in microneutralization assays, A/Vietnam/1203/04 (VN1203) was kindly provided by Dr. Richard Webby (St. Jude Children's Research Hospital, Memphis, TN). The virus was cultured in embryonated chicken eggs, and the allantoic fluid was collected, clarified, aliquoted, and stored at −80°C until use. All work with the virus was done using appropriate biosafety level-3 (BSL-3) biocontainment facilities and practices, and was approved by the institutional biosafety committee and relevant regulatory agencies.
Recombinant DNA vaccines
The cDNA for the HA of VN1203 (AY818135) was commercially codon-optimized (human codon preference) and synthesized with the polybasic cleavage site removed (Genscript Corp., Piscataway, NJ), and cloned into pcDNA3.0. The cDNA of influenza NP was cloned from influenza A/Puerto Rico/8/34 (PR8) using standard methods, and then transferred into pVAX using the Gateway cloning system (Invitrogen, Carlsbad, CA). Protein expression was confirmed by transfection of Vero E6 cells followed by immunohistochemistry and Western blot (data not shown). The SARS N (control) plasmid was the kind gift of Dr. Ralph Tripp (University of Georgia, Athens, GA). The SARS N gene was obtained by PCR, and the gene was inserted into the pcDNA3.0 mammalian expression vector. Expression was confirmed by immunofluorescence after transfection of the plasmid into Vero E6 cells. All plasmids were grown using standard microbial practices, and purified using large-scale plasmid purification columns (Invitrogen).
Animals
Fifteen specific pathogen-free 1-year old neutered male domestic short-haired cats obtained from Liberty Research, Inc. (Liberty, NY) were used in this study. All animals were seronegative prior to vaccination as measured by ELISA assay using inactivated H5N1 influenza virus (VN1203) as antigen. All animals were given food and water ad libitum, and handled in compliance with the University of Georgia Institutional Animal Care and Use Committee requirements. The University of Georgia is fully accredited by the Association for Assessment and Accreditation of Laboratory Animal Care, International (AAALAC).
Study design
The animals were randomly divided into five experimental groups composed of three cats each. In three of the experimental groups, one group received a plasmid with cDNA encoding the HA from an H5N1 influenza virus (H5 VN1203; 200 μg pcDNA 3.1 vector [Y1933], codon-optimized); another group was administered a plasmid encoding influenza A NP cDNA (NP PR8; 200 μg, pVAX vector, native sequence); and the third group received influenza H5 HA (VN1203) baculovirus-expressed recombinant protein (BEI Resources, Manassas, VA) in montanide (SEPPIC, Paris, France). The negative control group was vaccinated with a plasmid containing non-influenza cDNA (SARS virus N protein; 200 μg pcDNA 3.1 vector). The positive control group was administered the IND H5N1 inactivated subvirion vaccine (10 μg; BEI Resources). The plasmid vaccines (200 μg) were intramuscularly (IM) administered at two different sites (100 μg/site) in each hind leg. The recombinant H5 HA protein (10 μg) was mixed with an equal volume of montanide (0.6 mL total), and the inactivated H5N1 subvirion vaccine was diluted in PBS (10 μg in 0.33 mL). Both were administered IM. All animals were boosted using the same protocol 3 wk after the initial vaccination. Injection sites were monitored and no adverse reactions were noted. Blood samples (3 mL) were taken under mild sedation with acepromazine (0.05–0.1 mg/kg SC; Butler Animal Health Supply, Dublin, OH) prior to vaccination, and at 21 and 44 d after the initial vaccination. ELISAs were run using both inactivated whole H5N1 influenza virus (VN1203), and recombinant H5 HA using serum samples as previously described (8).
Hemagglutination inhibition assay (HIA)
After RDE treatment, the cat sera were subjected to twofold dilutions on 96-well V-bottom plates. Inactivated whole H5N1 influenza virus (VN1203/04), corresponding to 4 HA units, was then added to each well. After incubation at room temperature for 30 min, 0.8% chicken RBCs were added to each well. After gentle mixing, the plates were allowed to sit for an additional 40 min at room temperature. The HIA titer was interpreted as the highest dilution of sera that resulted in complete inhibition of hemagglutination using 4 HA units.
Microneutralization assay
One day prior to assay, Madin-Darby canine kidney (MDCK) cells were added to 96-well flat-bottom tissue culture plates at a concentration of 3 × 104 cells/well and incubated overnight at 37°C and 5% CO2. Cat sera were heat inactivated at 56°C for 30 min and allowed to cool to room temperature. In duplicate, samples were twofold serially diluted in MDCK infection media (MEM + l-glutamine + 1 μg/mL TPCK trypsin) in a 96-well round-bottom plate with a final volume of 100 μL/well. An equal volume of influenza virus (VN1203) at a concentration of 1 × 103 TCID50/mL diluted in infection media was added to the samples and incubated at 37°C and 5% CO2 for 1 h. After incubating, the samples were added to the plates containing MDCK cells that had been rinsed previously with sterile PBS. The plates were then incubated for 48 h at 37°C and 5% CO2. After incubating, 50 μL of culture supernatant was mixed with 50 μL of 0.5% chicken red blood cells and incubated for 30 min at room temperature. Samples were scored based on the presence or absence of hemagglutination. Data are presented as the reciprocal of the serum titer that prevented virus infection as measured by hemagglutination.
Results and Discussion
The discovery that both domestic cats and large felids are naturally susceptible to infection with highly pathogenic avian H5N1 influenza virus has raised concern over the role of cats in the potential transmission of virus to people, and the responses of both officials and pet owners to even small well-contained H5N1outbreaks are concerning (1,2,4,8,9,17). In addition, recent studies have found that cats can be subclinically infected with H5N1 influenza virus, and that commercial influenza antigen tests designed for human use, which are being sold to veterinarians for use in cats, have a very low sensitivity and did not detect virus in experimentally inoculated animals (13). All of these factors suggest that a feline vaccine against H5N1 influenza virus is needed.
In the current study, recombinant H5 protein (rH5) vaccine produced the highest antibody titers against both recombinant H5 protein and whole inactivated VN1203 virus as measured by ELISA (Fig. 1), HIA assay (Fig. 2A and B), and virus neutralization assay (VN; Fig. 2C). Specifically, a single immunization with the rH5 vaccine induced a >10-fold increase in HIA titer in 2/3 cats, and a >100-fold increase in the remaining animal (Fig. 2A). In humans, a HIA titer of 40 or a four-fold increase in HIA titer after vaccination is indicative of a protective response after vaccination with the trivalent inactivated influenza vaccine (5). Moreover, Vahlenkamp et al. demonstrated that cats vaccinated twice with an adjuvanted heterologous H5N6 whole virion inactivated influenza vaccine having HIA titers of 9–17 against the H5N1 challenge virus, were protected from lethal challenge (19). Thus the HIA responses induced by a single immunization with the rHA are likely protective against H5N1 infection. A second immunization with rHA resulted in a >100-fold increase in HIA titer, and >1000-fold increase in VN titer in all animals compared to pre-vaccination data (Fig. 2B and C, respectively). Although the concentration of H5HA was the same (10 μg) as that in the recombinant protein vaccine, a single dose of subvirion H5N1 IND vaccine only induced a very modest increase in antibody titers as measured by ELISA (Fig. 1B and 1E), and there was no detectable increase in HIA titer (Fig. 2A). A second administration of subvirion vaccine induced substantial antibody responses against recombinant H5 HA in ELISA (Fig. 1C), but surprisingly induced little response against whole inactivated virus (Fig. 1F). The second vaccination with H5N1 IND vaccine also increased the HIA titer in sera from all animals in the group, and induced high VN titers (GMT of 323 for 3 animals); however, none of the animals in this group possessed serum HIA titers greater than or equal to the levels observed in cats receiving the adjuvanted rHA vaccine (Fig. 2B). The more sensitive VN assay demonstrated higher titers in the H5N1 IND vaccinated animals; however, the adjuvanted rHA vaccine titers still met or exceeded the highest serum antibody titers measured from the H5N1 IND vaccinated cats (Fig. 2C).
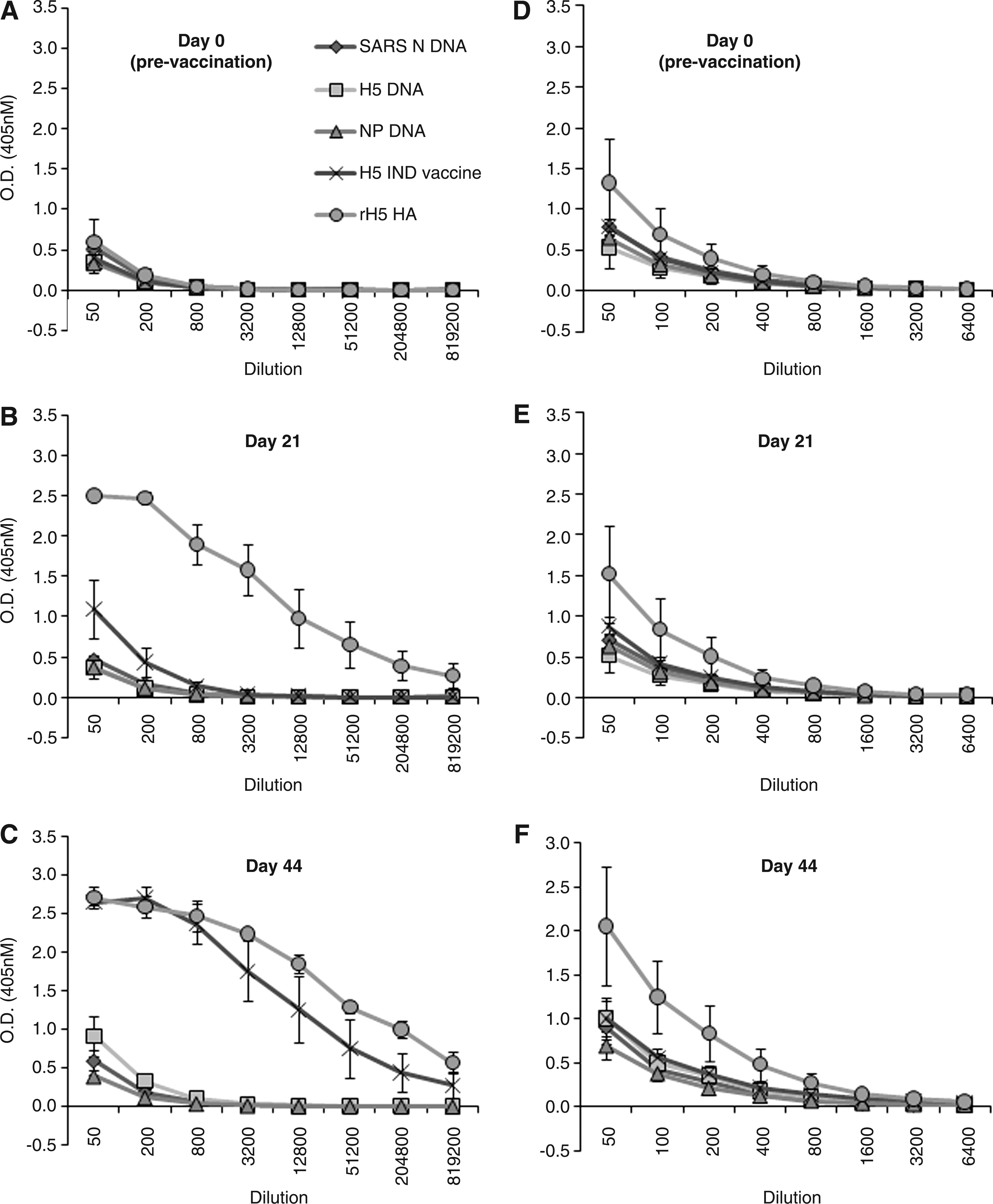
Vaccination of cats with different influenza virus vaccines induces hemagglutinin-specific serum antibody responses. Groups of cats were vaccinated with plasmid DNAs encoding the SARS-CoV nucleoprotein (SARS N DNA), H5 influenza virus hemagglutinin (H5 DNA) protein, influenza virus nucleoprotein (NP DNA), H5 influenza virus IND vaccine (H5 IND vaccine), or baculovirus-produced recombinant H5 HA protein (rHS HA). ELISA assays were performed on individual serum samples isolated from each animal on day 0, day 21, and day 44, using recombinant H5 HA protein (
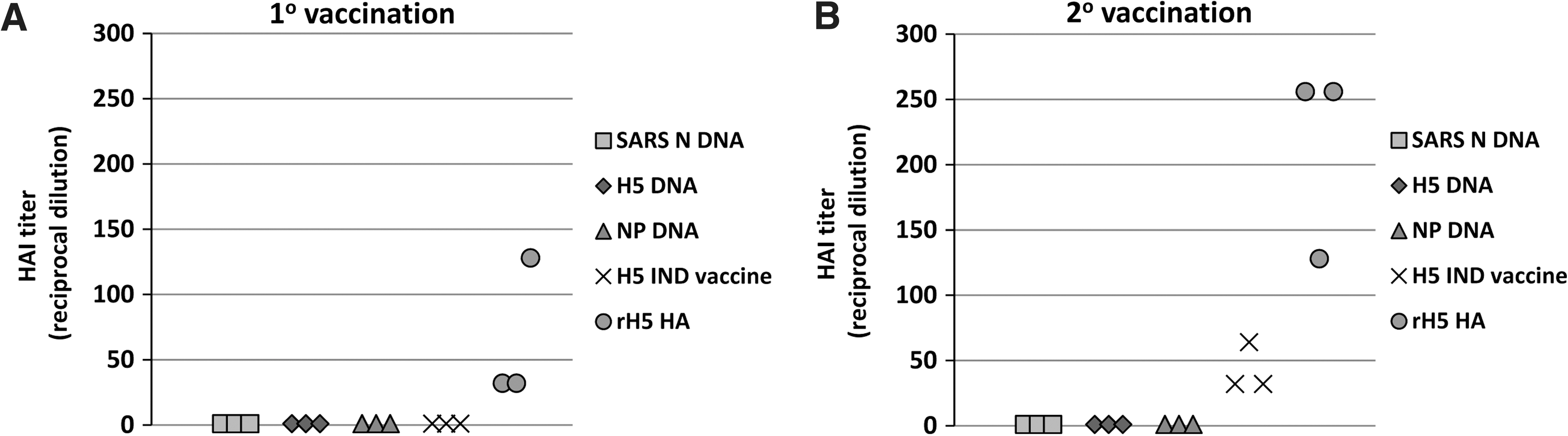
Cats vaccinated with inactivated virus or recombinant H5 HA protein developed serum antibody responses as measured by HIA and virus neutralization (VN) assays with VN1203 H5N1 influenza virus. Sera from individual animals taken at day 21 (
As expected, vaccination with the control plasmid encoding the SARS-CoV N protein did not induce antibody responses against rH5 protein or whole inactivated virus (Figs. 1 and 2). However, specific antibodies against whole virus and recombinant H5 HA were not detected in any of the cats that received plasmids encoding the H5 HA protein or the influenza virus NP (Fig. 1). In correlation with ELISA data, HIA titers were also below the limit of detection in all groups receiving plasmid DNA-based vaccines (Fig. 2). Moreover, sera from cats receiving the NP-expressing plasmid were tested in a commercially available species-independent NP-specific competition ELISA and showed no NP-specific serum antibody responses compared to control sera (data not shown). Given that protein expression by the plasmid-based vaccines was confirmed in vitro prior to administration, the failure of the influenza plasmid vaccines to induce measurable immune responses, even when measured by ELISA using whole virus as antigen (Fig. 1D, E, and F), was unexpected. These data suggest that multiple booster vaccinations or alternate routes of administration may be required when using plasmid-based vaccines in felines. Others have reported improved immunogenicity by intradermal immunization with a plasmid vaccine against rabies virus (18), while a DNA vaccine encoding calicivirus capsid failed to induce serum antibody responses, but provided partial protection against feline calicivirus challenge (16). Whether immunization with influenza H5 HA- or NP-expressing plasmid constructs can provide protection against influenza infection, despite the absence of detectable serum antibody responses, will require additional investigation.
Previous studies demonstrated that H5 HA is immunogenic in cats if delivered in the context of a viral-vectored vaccine. Detectable HIA serum antibody responses against both homologous and heterologous H5 influenza HA were shown in cats vaccinated with a fowlpox virus encoding the H5 HA gene (7). A single administration of the viral-vectored vaccine induced measurable antibody responses to homologous antigen, but HIA titers against a heterologous antigen were only detected after booster vaccinations (7). Interestingly, a single dose of the rH5 vaccine induced the generation of antibodies against H5 antigen, as measured by ELISA and HIA assays, equivalent to or stronger than the live poxvirus-based vaccine (7). When these data are compared, they suggest that robust antibody responses against H5 HA protein, as measured by both ELISA and HIA assay, can be achieved by recombinant protein without the use of a live virus vector; however, reactivity against other H5 HA antigens was not determined.
Recently it was shown that cats can be protected against a lethal high-dose challenge infection with highly pathogenic H5N1 avian influenza virus by an inactivated, whole virion H5N6 avian influenza virus vaccine (19). These results emphasize the importance of the immune response to the HA, since the challenge virus was heterologous to the vaccine virus for neuraminidase (19). The H5 recombinant protein vaccine used in this study induced higher HIA titers than the inactivated H5N1 IND subvirion vaccine; therefore it would be expected to be at least, if not more, protective against a similar heterologous challenge.
The results of this study indicate that a recombinant H5 protein vaccine induces a robust humoral immune response in cats after a single dose. This approach has several advantages over traditional inactivated and live viral-vectored influenza virus vaccines. First, recombinant protein-based vaccines are generally considered to be safe for use in animals and humans since they are not infectious. Given the possibility that cats may be infected with feline immunodeficiency virus when vaccinated, the administration of a recombinant protein would be less of a concern. Second, recent advances and options for the production of recombinant proteins in prokaryotic, baculovirus, and mammalian cell culture make this approach less costly and more attractive as a potential vaccine approach. Third, the production of recombinant protein does not require extensive biocontainment procedures or large scale propagation of virus in chicken eggs. Thus an efficacious, single-dose, recombinant protein-based vaccine could be made without using resources needed to produce influenza vaccines for human use.
The recent report of a companion animal cat being infected by the pandemic H1N1 human influenza virus (American Veterinary Medical Association Press Release, November 4, 2009;
Footnotes
Acknowledgments
We thank the Winn Feline Foundation for providing funding for this study. We also acknowledge the expert assistance provided by Tanya Cooper RVT, and the University of Georgia Research Animal Resources animal care personnel. In addition, we would like to thank Dr. Egbert Mundt for guidance in performing HIA assays. The following reagents were obtained through the National Institutes of Health (NIH) Biodefense and Emerging Infections Research Resources Repository, National Institute of Allergy and Infectious Diseases, NIH: Monovalent Influenza Subvirion Vaccine, rgA/Vietnam/1203/2004 (H5N1), IND batch #04-065, NR-4143; Recombinant H5 HA protein, lot #4146578, NR-660.
Author Disclosure Statement
No competing financial interests exist.