
Abstract
The roles of regulatory T cells (Tregs) and PD-1 in hepatitis B have not been clearly described. Also, the role of B and T lymphocyte attenuator (BTLA), which serves as a negative regulator of T-cell activation, is still unknown in hepatitis B. In this study, we analyzed the frequency of circulating CD4+CD25high Tregs in patients with chronic hepatitis B (CHB), and subsequently investigated expression of PD-1 and BTLA on CD4+ T cells, as well as their relationships with the clinical index of CHB patients. A total of 39 CHB patients and 19 healthy persons as controls were enrolled in the study. We found that the frequency of CD4+CD25high Tregs and PD-1 expression on CD4+ T cells was significantly increased in CHB patients compared with normal controls. However, BTLA expression on CD4+ T cells showed no significant difference between the two groups. The frequency of Tregs was significantly higher in patients with HBV DNA titers ≥108 than in those with HBV DNA titers <108. Circulating CD4+CD25high Treg frequency and PD-1 expression on CD4+ T cells correlated positively with serum HBV DNA load in CHB patients. Our findings suggest that the increased frequency of CD4+CD25high Tregs and PD-1 expression on CD4+ T lymphocytes may inhibit the cellular immune response against HBV and affect viral clearance, leading to the persistence of chronic HBV infection.
Introduction
In recent years, regulatory T cells (Tregs) and related negative regulation molecules have been a focus of research in the immune mechanism of disease. Tregs represent approximately 5–10% of circulating CD4+ T cells in mice and humans (28), and play an important role in the maintenance of immunological tolerance to both self and foreign antigens by suppressing aggressive T-cell responses (16,24,27). Evidence indicates that removal or reduction of Tregs can enhance immune responses against infectious microbes (1,2,14,15,21,25,31,40). Expression of programmed cell death 1 (PD-1, also called CD279), a negative regulatory molecule in the CD28/B7 family, is induced on CD4+ T cells, CD8+ T cells, natural killer T cells, B cells, and activated monocytes during the course of many diseases. The intracellular domain of PD-1 contains two tyrosine signaling motifs (ITIM and ITSM). Data indicate that PD-1 inhibits T-cell function by recruiting the intracellular phosphatase SHP2 that dephosphorylates and deactivates downstream signal transducers (8,13,22). B and T lymphocyte attenuator (BTLA) is a newly discovered member of the CD28 family of co-stimulatory molecules that may have an inhibitory function similar to that of PD-1 and CTLA-4. BTLA may have a role in controlling late phases of immune responses and may play a role in autoimmunity (7,37). Tregs, PD-1, and BTLA, which play an important role in chronic viral infection, not only serve as negative immune regulatory components, but also have some mutual synergies. Blocking the PD-1/PD-L1 pathway by anti-PD-L1 monoclonal antibody can inhibit the function of Tregs.
It has been demonstrated that HBV-specific CD8+ cytotoxic T lymphocyte (CTL) and CD4+ T helper cell-induced immune responses play a crucial role in the control of HBV infection during the early stage (38). Quantity and/or functional abnormalities of CD4+CD25high Tregs can attenuate HBV-specific CTL responses, resulting in an inability to completely clear HBV from the body. The latest study indicates that the PD-1 signaling pathway is involved in chronic viral infection and may become a new therapeutic target. However, the role of BTLA in chronic HBV infection has not been investigated. In this study, we analyzed the frequency of circulating CD4+CD25high Tregs in patients with chronic hepatitis B (CHB) infection, as well as the expression of PD-1 and BTLA on CD4+ T cells, and their relationships with HBV-related clinical markers.
Materials and Methods
Study subjects
Thirty-nine chronic hepatitis B patients, including 37 hepatitis B e antigen (HBeAg)-positive and two HBeAg-negative individuals, were enrolled in the study. CHB diagnoses were made according to the diagnostic standard of the Chinese National Program for Prevention and Treatment of Viral Hepatitis. All patients were hospitalized or followed-up in our unit. Patients co-infected with human immunodeficiency virus (HIV), hepatitis C virus (HCV), or hepatitis D virus (HDV) were excluded from this study. In addition, patients who were pregnant or who had received antiviral, immunosuppressive, or immunomodulatory treatment during the last 12 months were also excluded from this study. The blood samples from 19 healthy donors were also selected as controls. Our study was approved by the ethics committee of the Fourth Military Medical University, and all participants gave informed consent before sampling.
Virological assessment
Hepatitis B surface antigen, anti-HB antibody, HBeAg, anti-HBe antibody, anti-HBc antibody, anti-HCV antibody, anti-HDV antibody, and anti-HIV antibody titers in the sera of the patients were determined using commercial enzyme immunoassay kits (Kehua Biotech, Shanghai, China). Serum HBV DNA was quantified by a sensitive real-time PCR kit (PG Biotech, Shenzhen, China), with a detection limit of 300 copies/mL.
Isolation of peripheral blood mononuclear cells (PBMCs)
PBMCs were isolated from 20 mL of fresh heparinized blood by Ficoll-Hypaque (Sigma-Aldrich, St. Louis, MO) density gradient centrifugation. After isolation, the PBMCs were immediately cryopreserved in a medium containing 85% fetal bovine serum (FBS) and 15% dimethylsulfoxide (DMSO). The PBMCs were thawed with a step-by-step, gradual dilution method (26). Cell viability was over 90% as assessed by trypan blue exclusion.
Flow cytometry analysis
Flow cytometric analysis was performed on the stored PBMCs samples using fluorochrome-conjugated antibodies specific for the surface markers CD4, CD25, PD-1, and BTLA. Approximately 1 × 106 PBMCs diluted with 2 mL of phosphate-buffered saline (PBS) with 1% FBS were transferred to 5-mL sterile tubes. The cells were harvested at 1500 rpm for 10 min at 4°C. The following antibodies were used for staining: anti-CD4-FITC (eBioscience, San Diego, CA), anti-CD25-PE-Cy5 (eBioscience), anti-PD-1-APC (eBioscience), and anti-BTLA-PE (eBioscience). The cells were incubated and stained at 4°C in the dark for 30 min, and then were analyzed with a four-color FACSCalibur analyzer (BD Biosciences Immunocytometry Systems, San Jose, CA, USA). Acquisitions were performed with CellQuest Pro software (BD Biosciences Immunocytometry Systems, San Jose, CA, USA), and analyses were performed with FlowJo version 5.7.2 for Windows (Tree Star Inc., Ashland, OR). Isotype control antibodies were used to separate positive and negative cells in the FITC, PE-Cy5, PE, and APC fluorescence channels.
Semi-quantitation of BTLA mRNA by RT-PCR
Total RNA was extracted from PBMCs (3 × 106 cells) obtained from CHB patients and normal controls. The PBMCs were resuspended in 1 mL of Trireagent (Ambion, TX, USA), and RNA was extracted according to the manufacturer's instructions. RNA was quantified using the RiboGreen Kit (Molecular Probes, Eugene, OR). The primers used were commercially available: BTLA-U: 5′-AGA AGC AAG CAC CAG GCA AAA-3′, BTLA-L: 5′-GAC CCA AGC ACT AAC ATG AAC A-3′. As an internal control, reverse transcription PCR (RT-PCR) for GAPDH was also performed in our experiments. The extracted RNA was reverse transcribed with the TIANScript reverse transcription kit (Invitrogen Life Technologies, Carlsbad, CA) according to the manufacturer's instructions. The RT-PCR reaction was performed in a MyCycler thermal cycler (Bio-Rad, Hercules, CA). The resulting RT-PCR products were separated by gel electrophoresis on 2% agarose gel with 0.5 μg/mL ethidium bromide (EB) in Tris borate ethylenediaminetetraacetic acid (TBE) buffer, visualized using a Vilber Lourmat transilluminator, and photographed with the use of Biocapt MW version 10.02 software for Windows (Vilber Lourmat, Cedex, France).
Statistical analysis
All data were analyzed using SPSS version 13.0 for Windows software (SPSS Inc., Chicago, IL). Mann-Whitney U tests were used for the comparison between groups. Pearson's correlation tests were performed for correlation analysis. All tests were two-tailed, and p values <0.05 were considered significant.
Results
Increased frequency of circulating CD4+CD25high Tregs in CHB patients
We analyzed peripheral blood from 39 CHB patients and 19 normal controls (the clinical parameters are listed in Table 1), to determine the percentage of CD25+ T cells in the total CD4+ T-cell population. In the CHB patients, the CD4+CD25high population represented 4.48–13.89% of CD4+ T cells, but the CD4+ CD25+ T cell population represented 13.00–32.03% of CD4+ T cells (Fig. 1A and B). These data indicate that the frequency of CD4+CD25high Tregs in CHB patients (mean ± SD: 7.38 ± 2.02%) was significantly higher than in normal controls (mean ± SD: 4.99 ± 1.15%) (p < 0.0001; Fig. 1C). However, there was no statistically significant difference in the frequency of the CD4+CD25+ T-cell populations between the two groups (p = 0.164; Fig. 1D).
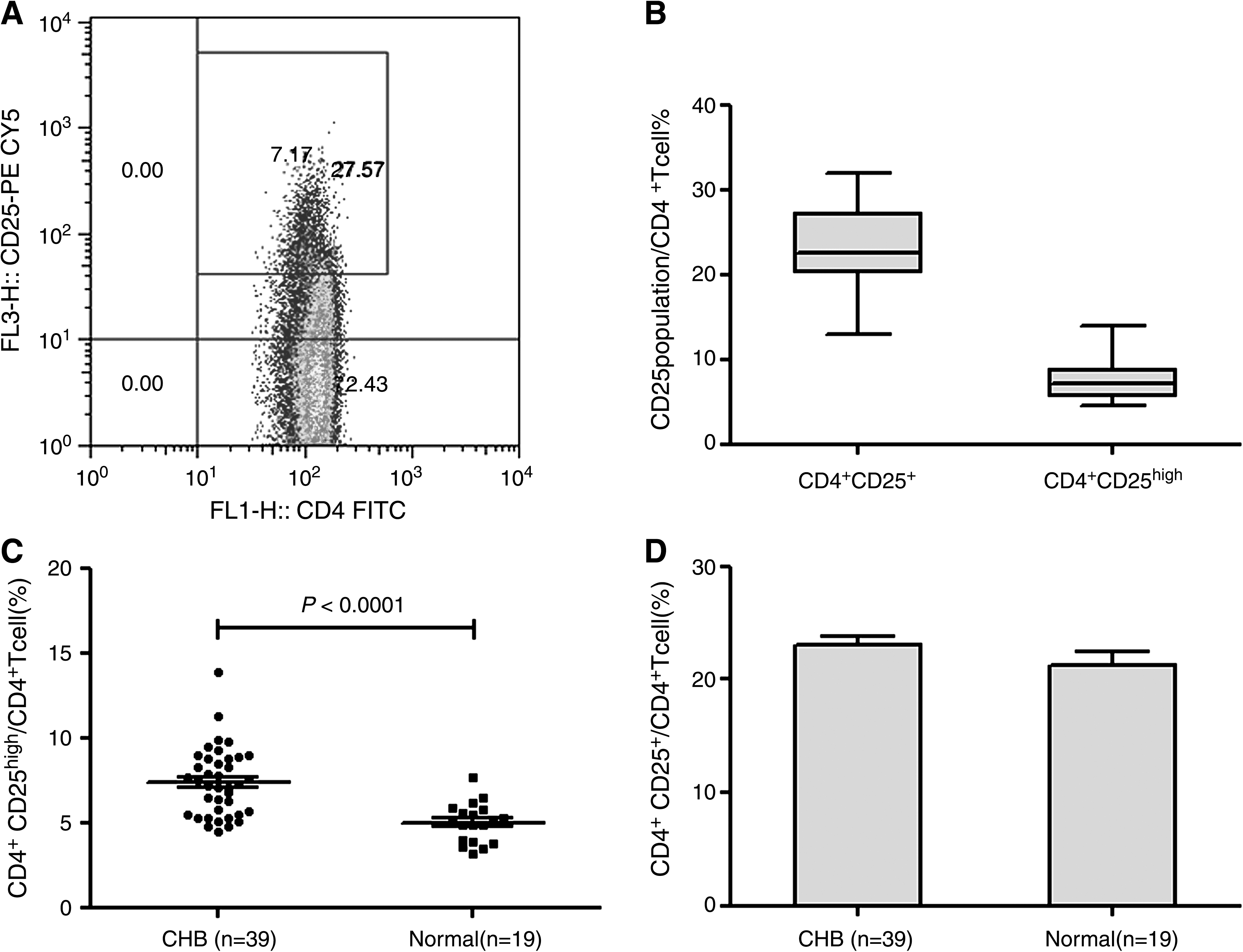
Frequency of circulating CD4+CD25+ T regulatory T cells (Tregs) in CHB patients and normal controls. (
Abbreviations: ALT, alanine aminotransferase; HBV, hepatitis B virus; HBeAg, hepatitis B e antigen.
The symbols + and − in parentheses represent the positive and negative results determined on serum HBV antigen, or refer to a detectable or undetectable level.
Expression of PD-1 and BTLA on CD4+ T cells in CHB patients
PD-1 and BTLA are important negative immune regulatory molecules. The percentage of CD4+ PD-1+ T cells in CHB patients (mean ± SD: 58.73 ± 11.11%) was significantly higher than that in normal controls (mean ± SD: 47.75 ± 12.69%) (p = 0.0013; Fig. 2C). However, the percentage of CD4+ BTLA+ T cells in CHB patients (mean ± SD: 2.79 ± 0.29%) showed no statistically significant difference with that of normal controls (mean ± SD: 2.386 ± 0.4810%) (p = 0.085; Fig. 2D). Furthermore, we also tested BTLA expression at the mRNA level. The relative BTLA mRNA levels in PBMCs from normal controls (n = 10) and CHB patients (n = 16) were 1.010 ± 0.1449 and 1.097 ± 0.0634, respectively (Fig. 2E); there was no significant difference between the groups (p = 0.916). The percentage of CD4+CD25high PD-1+ T cells in CHB patients (mean ± SD: 97.78 ± 5.657%) was not significantly different than that of normal controls (mean ± SD: 99.11 ± 1.133%) (p = 0.37; Fig. 3A and C). The percentage of CD4+CD25high BTLA+ T cells in CHB patients (mean ± SD: 9.811 ± 6.013%) showed no statistically significant difference with normal controls (mean ± SD: 13.97 ± 10.84%) (p = 0.11; Fig. 3A and B).

PD-1 and BTLA expression in circulating CD4+ T cells in CHB patents and normal controls. (

PD-1 and BTLA expression in circulating CD4+CD25high T cells in CHB patents and normal controls. (
Relationships between CD4+CD25high Tregs, CD4+PD-1+ T cells, and serum HBV DNA quantities
CHB patients were divided into two groups based on their serum HBV DNA load. A total of 108 copies/mL was used as the cut-off value of HBV DNA. Patients with a serum HBV DNA load ≥108 copies/mL had a higher mean percentage of circulating CD4+CD25high Tregs than patients with serum HBV DNA loads <108 copies/mL (7.75 ± 2.18% versus 6.55 ± 2.34%, respectively; p = 0.0043). There were also significant differences in CD4+CD25high Treg populations between the two groups and normal controls (Fig. 4A). Patients with serum HBV DNA loads ≥108 copies/mL showed no statistically significant difference in the number of cells expressing PD-1 on CD4+ T cells compared with patients with serum HBV DNA loads <108 copies/mL (61.18 ± 12.02% versus 54.36 ± 7.88%, respectively; p = 0.0552), but the level of PD-1 expression on CD4+ T cells in patients with serum HBV DNA loads ≥108 copies/mL was higher than that in patients with serum HBV DNA loads <108 copies/mL. There were significant differences between CD4+ PD-1+ T cells in patients with serum HBV DNA loads ≥108 copies/mL and patients with serum HBV DNA loads <108 copies/mL compared with normal controls (Fig. 4B).
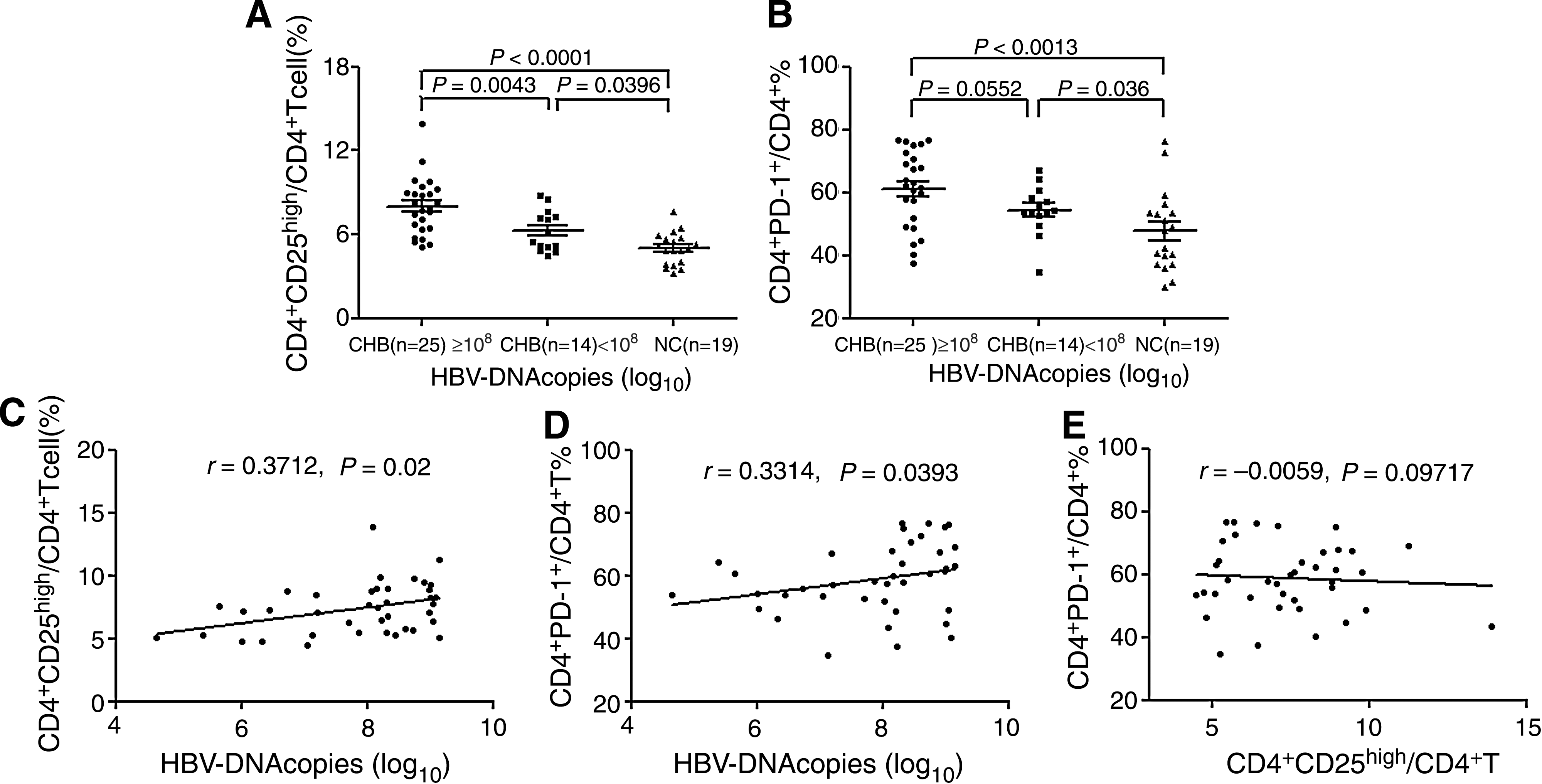
Association between serum HBV DNA load and circulating CD4+CD25high Treg frequency or CD4+ PD-1+ T cells in CHB patients. Patients were divided into two groups based on serum HBV DNA load (
Factors related to PD-1 expression and Tregs in CHB patients
To investigate whether the increase in circulating CD4+CD25high Tregs and CD4+ PD-1+ T cells correlated with HBV replication level, we measured the viral titers of serum from all tested CHB patients. Spearman analysis showed that the frequency of CD4+CD25high Tregs (r = 0.3314, p = 0.0393) and CD4+ PD-1+ T cells (r = 0.3712, p = 0.02) was positively correlated with serum HBV DNA load (Fig. 4C and D). However, there was no correlation between Treg frequency, PD-1 expression, and serum alanine aminotransferase (ALT) level (data not shown) in the CHB patients. These results suggest that an increased frequency of CD4+CD25high Tregs and CD4+ PD-1+ T cells may be associated with a weaker immune response, leading to poor viral clearance in CHB patients. In addition, there was no correlation between CD4+CD25high T cells and PD-1 expression on CD4+ T cells in CHB patients (p = 0.9717; Fig. 4E).
Discussion
CD4+CD25+ Tregs represent a unique lineage of T cells with a critical role in maintaining immunological homeostasis, and are very potent suppressors of the immune response. Tregs have also been demonstrated to play a crucial role in mediating tolerance and suppressing the activation and proliferation of innate and adaptive lymphocytes (29,30,34). The generation of Tregs may be a normal process that occurs to prevent immunopathological damage, and thereby contributes to viral persistence. The mechanisms responsible for T-cell tolerance in chronic HBV infection are not completely understood. Some studies have shown that patients with HCV or HIV infection have increased CD4+CD25+ Treg frequency in peripheral blood (3,35). Removal of Tregs in HIV-infected PBMCs can significantly enhance the ability of anti-HIV-specific CD4+ and CD8+ T cells to secrete cytokines (18). In our study, we found that the frequency of circulating CD4+CD25high Tregs in CHB patients was significantly higher than that of normal controls. These results agree with the results published by Stoop et al. (32), which showed that the frequency of CD4+CD25+ CD45RO+CTLA4+ Tregs in CHB patients was significantly higher than in healthy controls. However, Franzese et al. (12) found no difference in levels of CD4+CD25high Tregs between CHB patients and healthy controls. These contradictory results might stem from a lack of specific and stable markers, and a nomenclature that poorly reflects the complexity of Treg populations. Despite these varied results, growing evidence still suggests that Tregs are involved in the suppression of antiviral T-cell responses in chronic CHB infection.
High viral and/or antigen load may be an important cause of the T-cell hyporesponsiveness to HBV infection that is often seen in patients with CHB. Recovery of T-cell reactivity is likely to be essential for resolution of chronic viral infections. Boni et al. (4,6) found that lamivudine treatment, accompanied by a decline in patient viral load, can restore the host immune response, including T-helper cells and cytotoxic T-cell activity. These results suggest that high viral loads contribute to persistent T-cell hyporeactivity in CHB. We observed that patients with serum HBV DNA loads ≥108 copies/mL had a higher mean percentage of circulating CD4+CD25high Tregs than patients with serum HBV DNA loads <108 copies/mL and normal controls. Further analysis also showed that there was a positive correlation between circulating CD4+CD25high Treg frequency and serum HBV DNA load. The results are identical to the data of Xu et al. (39) and Yang et al. (41). However, two earlier studies did not find a significant association between circulating Treg frequency and HBV DNA load in CHB patients (12,32). Our results showed that an increase in Treg populations is associated with an increase in HBV replication. These data indicate that the immunosuppressive effect of Tregs may lead to virus replication, suggesting that Tregs may be a potential cause of the immune tolerance to chronic hepatitis B, and may contribute to the maintenance of active viral replication, thus subverting the host immune response. In addition, a positive correlation between HBeAg level, HBV DNA level, and the frequency of Treg cells in the blood of chronically infected patients further supports a role of Treg cells in HBV infection (33,39,41).
PD-1 belongs to a family of negative co-stimulatory molecules that appears to play an important role in the regulation of peripheral tolerance and antiviral immunity. The interaction between PD-1 and its ligand programmed death-1 ligand-1 (PD-L-1) has been shown to contribute directly to T-cell dysfunction and poor control of viral replication in established chronic viral infection models in mice (19). Several groups have demonstrated that PD-1 expression is significantly higher on HBV-specific CD8+ T cells from CHB patients, and that blocking PD-1–PD-L-1 interactions in vitro reverses the exhaustion of viral-specific CD8+ T cells and restores cytokine production and proliferation (5,10,23). These studies indicate that PD-1 expression on virus-specific T cells may limit antiviral effectiveness by antagonizing function and proliferation, and may be a potentially valuable therapeutic target in chronic viral infections. Our data indicate that PD-1 expression on CD4+ T lymphocytes in chronic HBV infection was significantly increased compared with normal controls, but there was no difference in expression of PD-1 on CD4+CD25high T cells in peripheral blood between chronic hepatitis B patients and normal controls. Also, there was no difference of PD-1 expression on CD4+ T lymphocytes between that of patients with HBV DNA loads ≥108 copies/mL and that of patients with HBV DNA loads <108 copies/mL. Our data also showed that the level of PD-1 expression on CD4+ T cells was positively associated with plasma HBV DNA load. The aforementioned results suggest that high viral load may stimulate PD-1 expression on CD4+ T cells and promote functional exhaustion. Collectively, these results confirm that high expression of PD-1 on CD4+ T cells may be involved in persistence of HBV viremia in CHB patients. In our study, we found no correlation between PD-1 expression on CD4+ T cells and the frequency of circulating CD4+CD25high Tregs. Both CD4+ PD-1+ T cells and CD4+CD25high T cells may be subpopulations of CD4+ suppressor cells in CHB. A recent study found that PD-L-1 negatively regulates Tregs by limiting STAT-5 phosphorylation in patients chronically infected with HCV (11). Future studies are needed to clarify the relationship between PD-1 expression and high expression of CD25 on CD4+ T cells.
BTLA is a newly discovered inhibitory receptor and antagonist of T-cell activation. BTLA-deficient mice showed evidence of an inhibitory role of BTLA in lymphocytes, based on increased antibody responses and heightened sensitivity to antigen-induced experimental allergic encephalomyelitis (37). It was found that BTLA expression was increased on infiltrating T cells of patients with lung cancer, but compared with healthy donors, there were no obvious differences between levels of BTLA expression on peripheral T lymphocytes of these patients (36). We also found no difference in expression of BTLA on CD4+ T cells in peripheral blood between chronic hepatitis B patients and normal controls, at both the mRNA and protein levels, Moreover, the expression of BTLA on CD4+CD25 high T cells in peripheral blood showed no significant difference between chronic hepatitis B patients and normal controls. We hypothesized that the inhibitory function of BTLA may be enhanced to some extent, but there was no obvious change in BTLA expression levels. Further detailed investigation of the expression and function of intrahepatic BTLA in chronic hepatitis B infection needs to be carried out before the role of BTLA becomes clear.
Conclusion
Taken together, our findings demonstrate that the frequency of circulating CD4+CD25high Tregs and PD-1 expression levels on CD4+ T cells were significantly increased in chronic HBV infection, and that these variables were positively correlated with serum HBV DNA load level in CHB patients. Tregs and PD-1, as negative regulators of immunity, may not only contribute to the persistence of chronic HBV infection, but may also be potential therapeutic targets for the treatment of CHB.
Footnotes
Acknowledgments
We thank the volunteers who generously participated in this study. This work was supported by a grant from National Key Technologies Research and Development Program of China during the 11th Five-Year Plan Period (no. 2008ZX10002-008).
Author Disclosure Statement
No conflicting financial interests exist.