
Abstract
Upregulation of interleukin-10 (IL-10) expression has been suggested to be the mechanism by which the porcine reproductive and respiratory syndrome virus (PRRSV) suppresses the innate and adaptive immune response in infected pigs. In this study we evaluated the potential of phosphorothioate-modified IL-10 antisense oligodeoxynucleotide specific to the translation initiation region of porcine IL-10 mRNA (IL-10AS) in enhancing proinflammatory cytokine responses to PRRSV. Naïve peripheral blood mononuclear cells from eight PRRSV-seronegative pigs were transfected with IL-10AS in vitro prior to PRRSV inoculation and phorbol 12-myristate 13-acetate plus ionomycin or concanavalin A stimulation. The effects of IL-10AS on mRNA expression of IL-10, interferon-γ (IFN-γ), IFN-α, tumor necrosis factor-α (TNF-α), IL-2, and IL-4 were tested by real-time PCR. The percentages of IFN-γ-producing T-cell subsets were determined by flow cytometry. Compared to the controls, the levels of IL-10 and IL-2 mRNA were significantly reduced, while those of IFN-γ mRNA were increased, and TNF-α, IFN-α, and IL-4 mRNA were unchanged. An increase in the percentage of the IFN-γ+ population was also observed in lymphocytes and CD8β+ T cells. Our results suggest that IL-10AS has the potential to enhance proinflammatory cytokine responses to PRRSV infection.
Introduction
PRRSV infects myeloid antigen-presenting cells (APCs) (i.e., monocytes, macrophages, and dendritic cells [DCs]) (6). The virus elicits poor innate and adaptive immune responses after infection. The innate immune response to PRRSV determined by phagocytosis, microbicidal activity, expression of major histocompatability and co-stimulatory molecules, and production of proinflammatory cytokines (e.g., interleukin-1 [IL-1], type I interferon [IFN], and tumor necrosis factor-α [TNF-α]) is reduced in PRRSV-infected cells (1,4,8,10,21,32,33,38,41,54,55,57).
The adaptive immune response to PRRSV determined by the antibody and cell-mediated immune (CMI) response is delayed. PRRSV-specific antibodies appear after approximately 2 wk of infection, and most of them are specific to non-neutralizing epitopes of the virus (37,59,62). Neutralizing antibodies to PRRSV appear much later, approximately 4 wk after infection, and have relatively low titers throughout the course of infection (37,59,62). The CMI response to the virus is determined by lymphocyte blastogenesis, and enzyme-linked immunospot and flow cytometry assays for IFN-γ production are positive approximately 4–8 wk after infection (2,3,7,31,34,44,53). This is extremely late compared to the CMI responses to other swine viral pathogens, which appear within 3 d to 1 wk after infection (53).
The poor innate and adaptive immune responses to PRRSV have been suggested to be attributable at least in part to virus-induced IL-10 production (49,50). PRRSV has been reported to upregulate IL-10 production following virus infection (4,6,8,12,18 –22,25,29,38,44,49,50). PRRSV-induced IL-10 production has been reported to correlate with reduced IFN-γ production in virus-infected cells (6,44,49,50). In pigs, IL-10 has been demonstrated to suppress IFN-γ production by peripheral blood mononuclear cells (PBMCs) and T cells (6,58). In humans and mice, this cytokine has been reported to suppress APC activities in processing and presenting antigens, and in expressing IL-1, IL-12, IL-18, TNF-α, and type I IFN (13,16,36,61). The cytokine has also been reported to suppress IL-2, IL-4, and IFN-γ expression by T-helper 1 (Th-1), Th-2, and natural killer cells (14,17,26,36,39,51,52).
Suppression of PRRSV-induced IL-10 production may be a strategy to improve the immune response to PRRSV. Our previous study has demonstrated that the addition of IL-10-specific monoclonal antibodies (mAbs) into PRRSV-inoculated PBMCs could help enhance IFN-γ production in co-cultivated T cells (6). Recently, there has been a report regarding the use of phosphorothioate-modified IL-10 antisense oligodeoxynucleotides specific to porcine IL-10 mRNA's translation initiation region (IL-10AS) to suppress IL-10 transcription in porcine PBMCs (46). We hypothesize that this IL-10AS could be another potential tool for controlling IL-10 expression induced by PRRSV, and for improving the immune response to the virus. In this study, we therefore evaluate the potential of IL-10AS in suppressing IL-10 mRNA expression in PRRSV-inoculated PBMCs. We also evaluate the contribution of IL-10AS to improving the proinflammatory cytokine mRNA response and IFN-γ production in inoculated PBMCs and co-cultivated T-cell subsets (i.e., CD8β+, CD4+, and SWC6+ cells). In pigs, these T-cell subsets function as cytotoxic αβ T cells (CD8β+), T-helper, and regulatory T cells (CD4+), and serve as a major population of peripheral blood γδ T cells (SWC6+) (5,15,23,27,40).
Here we report that IL-10AS has potential for suppressing IL-10 mRNA expression in PRRSV-inoculated PBMCs, and contributes to enhancing IFN-γ mRNA expression in inoculated PBMC cultures. IL-10AS also contributes to enhancing the percentage of IFN-γ+ cells in CD8β+ T cells. These findings indicate the potential of IL-10AS for potentiating the immune response to PRRSV.
Materials and Methods
IL-10AS
The IL-10AS sequence used was 5′-TGAGCTGGGCATGGTAGA-3′ as described by Sidahmed and Wilkie (46). The scramble control oligodeoxynucleotide (ODN) sequences were 5′-TGATTGTATACTTCTTGAT-3′ (Scr1), 5′-CCAGTGACGGTGCAGGCA-3′ (Scr2), and 5′-CAAGTGCACACAAGGCTA-3′ (Scr3). All phosphorothioate-modified ODNs were synthesized by 1st Base Pte Ltd (Singapore).
Antibodies
All primary and secondary mAbs were purchased from BD Pharmingen (San Diego, CA). Isotype control mAbs were from Southern Biotechnology Associates, Inc. (Birmingham, AL). Primary mAbs included fluorescein isothiocyanate (FITC)-conjugated mouse anti-porcine CD4 (clone 74-12-4, isotype IgG2b), unconjugated mouse anti-porcine CD8β (clone 295/33-25, isotype IgG2a), and unconjugated rat anti-porcine SWC6 (clone MAC320, isotype IgG2a). Secondary mAbs were FITC-conjugated rat anti-mouse IgG2a (clone R19-15), and FITC-conjugated mouse anti-rat IgG1/2a (clone G28-5). Intracellular staining mAbs were R-phycoerythrin (R-PE)-conjugated mouse anti-porcine IFN-γ (clone P2G10, isotype IgG1).
Virus and cell line
PRRSV isolate 01NP1 and MARC-145 cells were kindly provided by Dr. Roongroje Thanawongnuwech of the Veterinary Diagnostic Laboratory, Faculty of Veterinary Science, Chulalongkorn University, Bangkok, Thailand. The virus was propagated in MARC-145 cells grown in DMEM++ (DMEM, 10% heat-inactivated fetal bovine serum [FBS], penicillin [100 IU/mL], streptomycin [100 μg/mL], and amphotericin B [250 ng/mL]; all from Gibco, Grand Island, NY) to the titer of 106 tissue culture infectious dose 50%/mL (TCID50/mL). Virus titer was determined from cytopathic effects of the virus in MARC-145 cells after 3 d of PRRSV inoculation. The PRRSV used in this study was at its seventh passage. Mock antigens were prepared in the same fashion as virus antigens, except that no viruses were inoculated.
Isolation of PBMCs
Blood was collected from eight 24-wk-old PRRSV-seronegative pigs from a commercial producer using ethylenediamine tetraacetic acid (EDTA; J.T. Baker, Phillipsburg, NJ) as an anticoagulant. PBMCs were isolated from the blood sample by Ficoll-Hypaque gradient centrifugation using Histopaque®-1077 (Sigma-Aldrich, St. Louis, MO). Contaminating red blood cells in the isolated PBMCs were lysed with cold red blood cell lysis buffer (0.156 M ammonium chloride, 10 mM sodium bicarbonate, and 1 mM EDTA). The PBMCs were resuspended in reduced serum medium (Optimem® I; Gibco) to the concentration of 106 cells/mL.
Transfection of PBMCs with IL-10AS
Optimization of IL-10AS concentration
IL-10AS mixtures containing 0.5, 1, or 2 μM IL-10AS in transfection media (Lipofectamine™ RNAiMAX; Invitrogen, Carlsbad, CA, in Optimem I at 1.5% v/v) were prepared and incubated at room temperature for 30 min. Then 20 μL of the IL-10AS mixture were added to each well of a 96-well flat-bottom plate, followed by 100 μL of PBMCs (106 cells/mL). The cultures were mixed gently by rocking the plates back and forth for 5 min, then incubated at 37°C in a humidified 5% CO2 atmosphere for 4 h. The media were removed and replaced with 200 μL of RPMI++ (RPMI-1640 with L-glutamine, 10% heat-inactivated FBS, penicillin [100 IU/mL], streptomycin [100 μg/mL], and amphotericin B [250 ng/mL]; all from Gibco), and 50 μL of phorbol 12-myristate 13-acetate plus ionomycin (PMA/I; 7 and 430 ng/mL final concentrations, respectively; Calbiochem, Darmstadt, Germany) or concanavalin A (ConA; 5 μg/mL final concentration; Sigma-Aldrich). The plates were incubated at 37°C in a CO2 incubator for 12 and 16 h after PMA/I and ConA stimulation, respectively. The cells were then harvested and evaluated for IL-10 mRNA expression. PBMCs cultured without IL-10AS transfection in the presence or absence of cytokine inducers were used as positive and negative controls, respectively. PBMCs treated with scramble ODN or transfection media prior to cytokine stimulation served as scramble and transfection media controls, respectively.
Evaluation of IL-10AS specificity
PBMCs were transfected with IL-10AS (2 μM), and stimulated with either PMA/I or ConA as described above. PMA/I-stimulated cells were assessed for IL-10, IFN-γ, IFN-α, and TNF-α mRNA expression (6), and ConA-stimulated cells were assessed for IL-10, IL-2, and IL-4 mRNA expression (56).
Evaluation of the effects of IL-10AS on cytokine mRNA expression in PRRSV-inoculated PBMCs
PBMCs were transfected with IL-10AS as described above. After transfection, the media were removed and replaced with 100 μL RPMI++ and 100 μL PRRSV (106 TCID50/mL). The plates were incubated at 37°C in a 5% CO2 incubator for 48 h. For evaluation of IL-10, IFN-γ, IFN-α, and TNF-α mRNA expression, 50 μL of PMA/I were added and the plates were incubated for an additional 12 h (6). For determination of IL-10, IL-2, and IL-4 mRNA expression, 50 μL of ConA were added to the culture and the plates were incubated for an additional 16 h (56). For evaluation of intracellular IFN-γ production, 50 μL of PMA/I/G (PMA/I plus GolgiStop® solution; BD Pharmingen, at the manufacturer-recommended dilution) were added, and the plates were incubated for an additional 12 h (42). PBMCs treated with transfection media alone (without IL-10AS) served as PRRSV-inoculated transfection media controls, and those left untransfected served as PRRSV-inoculated controls. Other controls included PBMCs receiving culture media alone (negative control); culture media plus PMA/I, ConA, or PMA/I/G (positive control); and mock antigens plus PMA/I, ConA, or PMA/I/G (mock control). All PBMCs were determined for cell viability at the end of the transfection period, PRRSV inoculation, and PMA/I, ConA, or PMA/I/G stimulation using trypan blue (Gibco).
Reverse transcriptase PCR
RNA extraction and reverse transcription
Total RNA was extracted from PBMCs using the RNeasy® kit (Qiagen, Valencia, CA) according to the manufacturer's instructions. Contaminating DNA was eliminated by DNAse I (Fermentas, Glen Burnie, MD). Total RNA was eluted in 40 μL of RNase-free water, 10 μL of which were assigned to reverse transcription using the Omniscript RT kit (Qiagen), 0.5 μg random hexamers (Qiagen), and 40 U ribonuclease inhibitor (RiboLock™; Fermentas).
Real-time PCR
Complementary DNA (cDNA) was used as a template for real-time PCR of porcine IL-10, IFN-γ, IFN-α, TNF-α, IL-2, IL-4, and ribosomal protein L32 (RPL32). The PCR was performed on the MJ Research PTC-200 thermal cycler in a total reaction volume of 50 μL, consisting of 2 μL cDNA template, 0.3 μM each of forward and reverse primers, and 25 μL PCR buffer (QuantiTect SYBR Green PCR Master Mix; Qiagen). All primer sequences were from Royaee et al. (44). The PCR conditions were initial activation at 95°C for 15 min; and 40 cycles of denaturation at 94°C for 15 sec, annealing at 55° for 30 sec, and extension at 72°C for 30 sec. The threshold cycles (CT) of all genes were collected and used for calculation of cytokine mRNA expression by the ΔΔCT method. A melting curve analysis was performed after the completion of the PCR cycles. PCR products were determined for size correction by agarose gel electrophoresis, and visualized under ultraviolet light with Quantity One software (version 4.5.0; Bio-Rad, Hercules, CA).
Immunofluorescent staining
At the end of the PMA/I/G stimulation period, PBMC culture plates were chilled on ice for 15 min. The cells were harvested and washed twice with PBS++ (PBS, 0.5% heat-inactivated FBS, and 0.1% sodium azide; Fisher Scientific, Pittsburgh, PA). The washed cells were then incubated with 50 μL of primary mAbs (anti-CD4, anti-CD8β, or anti-SWC6) at an optimum dilution in the dark at 4°C for 30 min, followed by three washes with PBS++. For anti-CD8β and anti-SWC6 mAb staining, the cells were subsequently incubated with 50 μL of appropriate secondary antibody at 4°C in the dark, followed by three washes with PBS++. All cells were then fixed with 100 μL of BD Cytofix/Cytoperm solution (BD Pharmingen) for 30 min at 4°C in the dark, washed twice with 500 μL of BD Perm/Wash (BD Pharmingen), and 50 μL of R-PE-conjugated anti-IFN-γ mAbs were added. The cells were incubated in the dark for 30 min at 4°C, washed twice, and fixed with 300 μL of 1% formaldehyde (Fisher Scientific) in PBS. Membrane fluorescence of the stained cells was analyzed by a FACScan cytometer (BD Pharmingen). Flow cytometry data were analyzed using CellQuest analysis software (version 3.3; BD Biosciences, San Jose, CA). A dot plot of linear side scatter versus linear forward scatter was generated to gate lymphocyte populations from total PBMCs. A histogram based on FITC-conjugated CD4, CD8β, or SWC6 was applied to lymphocyte populations to differentiate T-cell subsets. A histogram based on R-PE-conjugated IFN-γ was applied to each T-cell subset to obtain the percentage of IFN-γ+ cells.
Statistical analysis
All statistical analyses were performed using JMP6 software (SAS Institute Inc., Cary, NC). Mean differences of IL-10, IFN-γ, IFN-α, TNF-α, IL-2, and IL-4 mRNA expressions were tested by one-way analysis of variance, followed by Dunnett's test, using the mean of the positive control group as a control, and where indicated, using the mean of the PRRSV-inoculated group or transfection media control group as a control. The same statistical methods were applied for the analysis of mean differences of the percentages of viable cells and the percentages of IFN-γ+ cells in lymphocytes and T-cell subsets. Statistical significance was set at p < 0.05 throughout this study.
Results
Determination of IL-10AS optimal concentration and specificity
IL-10AS has been demonstrated to downregulate IL-10 mRNA expression in ConA-stimulated porcine PBMCs (46). In this study, we further investigated the potential of using IL-10AS to downregulate IL-10 mRNA expression in porcine PBMCs inoculated with PRRSV in the presence of a cytokine inducer (i.e., PMA/I or ConA). To determine the optimal concentration of IL-10AS, PBMCs were transfected with IL-10AS at various concentrations and subsequently stimulated with PMA/I or ConA, and the IL-10 mRNA expression was analyzed by real-time PCR. The IL-10AS at 2 μM showed significant suppression of IL-10 mRNA expression in both PMA/I- and ConA-stimulated PBMCs (Fig. 1); thus this concentration was selected for further investigation. The IL-10AS used had no effect on IFN-γ, IFN-α, TNF-α, IL-2, and IL-4 mRNA expression in the stimulated PBMCs, which indicated its specificity to IL-10 mRNA (data not shown). The scramble ODN control PBMCs did not show any reduction in IL-10 mRNA expression (Fig. 1).
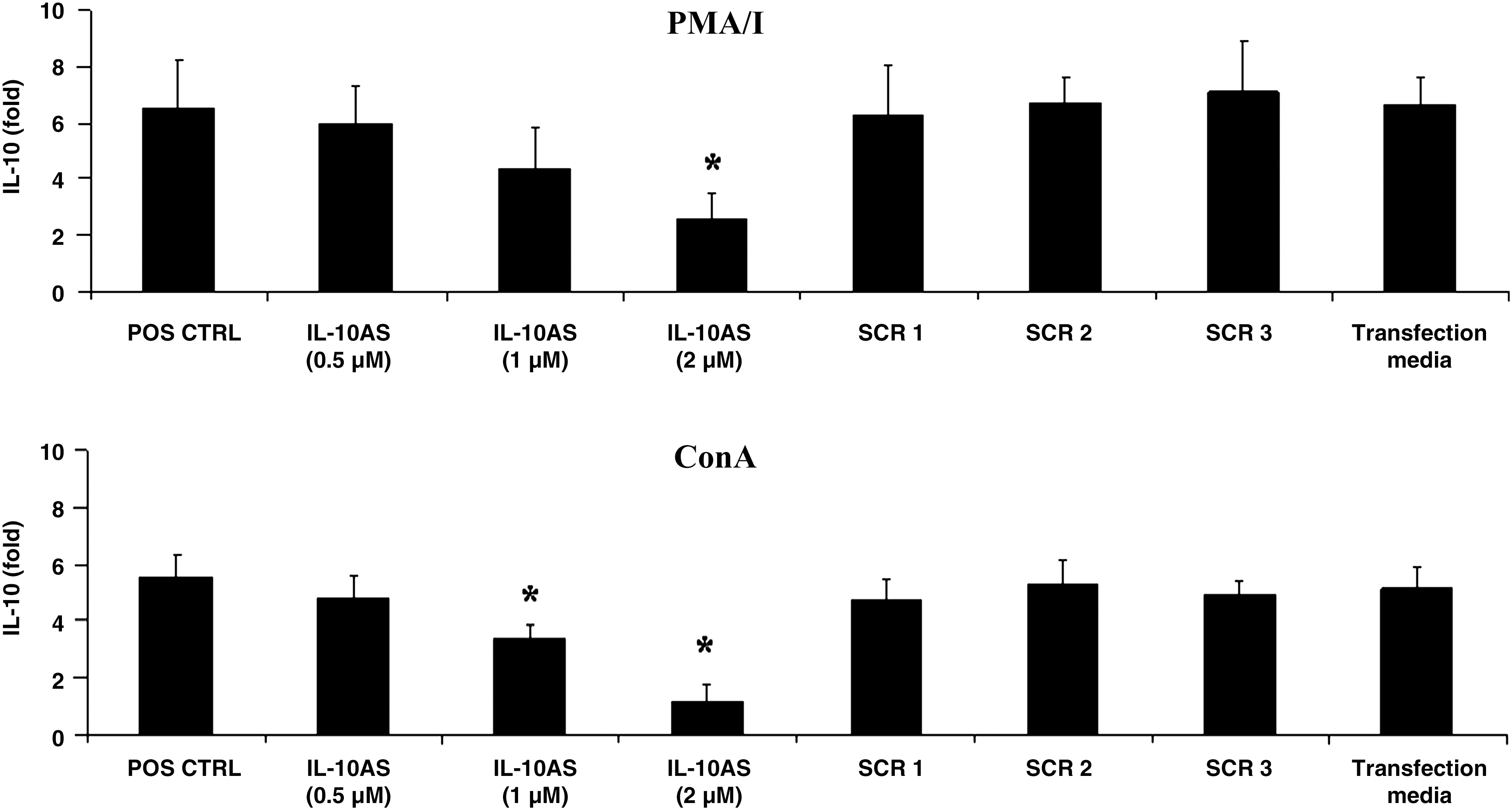
IL-10 mRNA expression in PBMCs transfected with various concentrations of IL-10AS and stimulated with PMA/I or ConA. PBMCs were transfected with the indicated concentrations of IL-10AS for 4 h and subsequently stimulated with PMA/I or ConA for 12 or 16 h, respectively, prior to real-time PCR analysis. PBMCs treated with culture media alone prior to stimulation with either cytokine inducer served as positive controls (POS CTRL). PBMCs transfected with scramble ODN (SCR1, SCR2, and SCR3) at 2 μM, or treated with transfection media (Transfection media) prior to stimulation with PMA/I or ConA, served as scramble controls and transfection media control, respectively. Data were normalized with RPL32 mRNA expression of the same animal. Error bars indicate the standard error of the mean (SEM). Asterisks represent significant mean difference of IL-10 mRNA expression between the treatment and positive control groups (p < 0.05).
IL-10AS suppressed IL-10 mRNA expression in PRRSV-inoculated PBMCs
PRRSV-inoculated PBMCs demonstrated significantly increased IL-10 mRNA expression in response to PMA/I or ConA stimulation, compared to the absence of PRRSV (positive control) (Fig. 2). Transfection of PBMCs with IL-10AS prior to PRRSV inoculation and cytokine stimulation significantly reduced IL-10 mRNA expression (Fig. 2). This observation was not due to different percentages of viable PBMCs after the treatments. The percentages of viability of PBMCs following IL-10AS transfection and PMA/I or ConA stimulation were 50.2 ± 3.8% and 47.1 ± 3.4%, respectively. These values were not significantly different from those of the positive control (49.3 ± 4.1% and 47.4 ± 3.4%) and the PRRSV-inoculated group (48.7 ± 3.1% and 46.3 ± 3.7%).
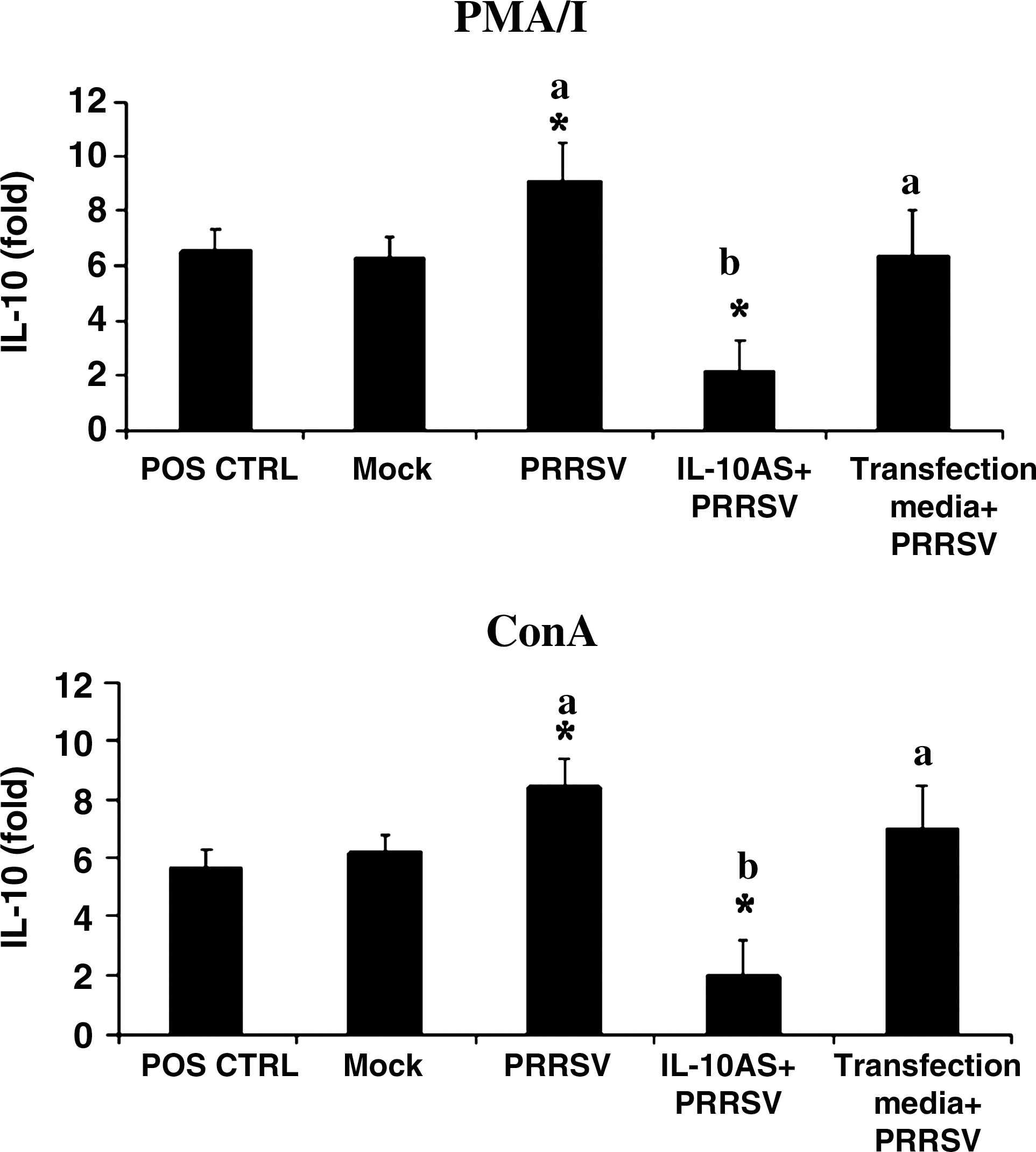
IL-10AS suppresses IL-10 mRNA expression induced by PRRSV. PBMCs were transfected with IL-10AS at 2 μM for 4 h, then cultivated with PRRSV for 48 h, and stimulated with PMA/I or ConA for 12 or 16 h, respectively (IL-10AS+PRRSV). The IL-10 mRNA was determined by real-time PCR analysis. PBMCs stimulated with PMA/I or ConA, in the absence of IL-10AS and PRRSV, served as positive controls (POS CTRL). PBMCs treated with culture media plus mock antigens or PRRSV and stimulated with either cytokine inducer served as the mock control (Mock) and PRRSV-inoculated control (PRRSV), respectively. PBMCs treated with transfection media (no IL-10AS) plus PRRSV (Transfection media+PRRSV), and stimulated with either cytokine inducer served as PRRSV-inoculated transfection media control. Data were normalized with RPL32 mRNA expression from the same animal. Error bars indicate the standard error of the mean. Asterisks represent a significant mean difference in IL-10 mRNA expression between treatment and positive control groups (p < 0.05). Different letters (a and b) indicate significant mean differences of IL-10 mRNA expression between the transfected and PRRSV-inoculated control groups (p < 0.05).
IL-10AS effects on proinflammatory cytokine mRNA expression in PRRSV-inoculated PBMCs
Compared to the absence of PRRSV, PRRSV-inoculated PBMCs demonstrated significantly reduced IFN-γ and IFN-α mRNA expression in response to PMA/I stimulation (Fig. 3). The virus-inoculated PBMCs also demonstrated significantly increased IL-2 mRNA expression in response to ConA stimulation (Fig. 3). No significant modulation in TNF-α or IL-4 mRNA expression was observed in inoculated PBMCs in response to PMA/I and ConA stimulation (Fig. 3). No significant modulation in any cytokine mRNA expression was detected in the mock controls (Fig. 3).

Effect of IL-10AS on IFN-γ, IFN-α, TNF-α, IL-2, and IL-4 mRNA expression. PBMCs were transfected with IL-10AS at 2 μM for 4 h, then cultivated with PRRSV for 48 h, and stimulated with PMA/I (for IFN-γ, IFN-α, and TNF-α), or ConA (for IL-2 and IL-4) for 12 or 16 h, respectively, prior to real-time PCR analysis (IL-10AS+PRRSV). PBMCs treated with culture media alone and stimulated with cytokine inducer served as the positive control (POS CTRL). PBMCs treated with culture media plus mock antigens or PRRSV and stimulated with cytokine inducer served as the mock control (Mock), and PRRSV-inoculated control (PRRSV), respectively. PBMCs treated with transfection media (no IL-10AS) plus PRRSV and stimulated with cytokine inducer served as PRRSV-inoculated transfection media control (Transfection media+PRRSV). Data were normalized with RPL32 mRNA expression from the same animal. Error bars indicates the standard error of the mean. Asterisks represent significant mean difference of cytokine mRNA expression between treatment and positive control groups (p < 0.05). Different letters (a and b) indicate significant mean differences in cytokine mRNA expression between the transfected and PRRSV-inoculated control groups (p < 0.05).
Compared to the PRRSV-inoculated PBMCs, PBMCs transfected with IL-10AS prior to PRRSV inoculation and cytokine stimulation demonstrated increased IFN-γ mRNA expression (p = 0.12), and significantly reduced IL-2 mRNA expression (Fig. 3). The PBMCs, however, demonstrated unchanged IFN-α, TNF-α, and IL-4 mRNA expression (Fig. 3). No significant modulation in any cytokine mRNA expression was observed in the PRRSV-inoculated transfection media controls (Fig. 3).
IL-10AS effects on the percentage of IFN-γ+ cells in lymphocytes and T-cell subsets
IL-10AS was investigated further for its contribution to the percentage of IFN-γ+ cells in lymphocytes and T-cell subsets (i.e., CD8β+, CD4+, and SWC6+ cells). In pigs, CD8β+ T cells serve as cytotoxic αβ T cells, while CD4+ T cells serve as T-helper cells, and a small percentage of them function as regulatory T cells (5,23,27,40). SWC6+ T cells, on the other hand, serve as a major subset of peripheral blood γδ T cells (15,23). Their IFN-γ response to PRRSV has never been determined.
The lymphocyte population from PRRSV-inoculated PBMCs had a significantly reduced percentage of IFN-γ+ cells in response to PMA/I/G stimulation in comparison to the absence of PRRSV (Fig. 4). Lymphocytes from mock control PBMCs did not show such a significant reduction (Fig. 4). The significant reduction of the percentage of IFN-γ+ cells was restricted to CD8β+, and was not observed for the CD4+ and SWC6+ T-cell subsets (Fig. 5). The significant reductions of the percentage of IFN-γ+ cells was not due to virus-induced cell death. The percentage of viable PBMCs after PRRSV inoculation and subsequent PMA/I/G stimulation was 45.7 ± 3.1%, which was not significantly different from that in the absence of virus (48.1 ± 5.4).

Effect of IL-10AS on the percentage of IFN-γ+ cells in the lymphocyte population. PBMCs were transfected with IL-10AS at 2 μM for 4 h, then cultivated with PRRSV for 48 h, and stimulated with PMA/I/G for another 12 h (IL-10AS+PRRSV). Lymphocytes from these PBMC cultures were harvested, gated, and determined for the percentage of IFN-γ+ cells by flow cytometry. Controls included lymphocytes from the following PBMC cultures: PBMCs treated with culture media alone and stimulated with PMA/I/G (positive control; POS CTRL); PBMCs treated with culture media plus mock antigens and stimulated with PMA/I/G (mock control; MOCK); PBMCs treated with culture media plus PRRSV and stimulated with PMA/I/G (PRRSV-inoculated control; PRRSV); and PBMCs treated with transfection media (no IL-10AS) plus PRRSV, and stimulated with PMA/I/G (PRRSV-inoculated transfection media control; Transfection media+PRRSV). Asterisks represent a significant mean difference in the percentage of IFN-γ+ cells between treatment and positive control groups (p < 0.05). The same letter (a) indicates no significant mean difference in the percentage of IFN-γ+ cells between transfected and PRRSV-inoculated control groups (p > 0.05).
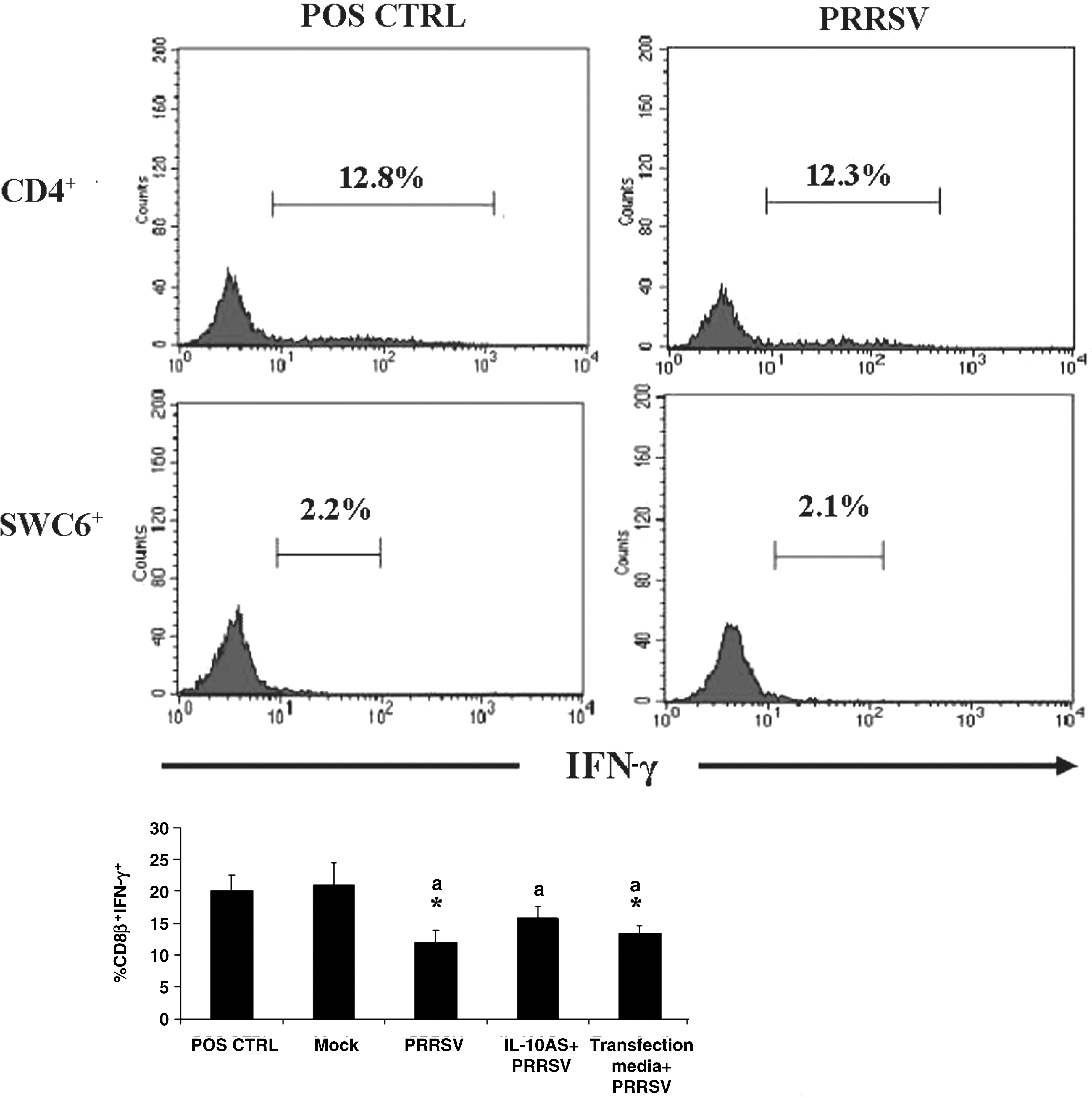
Effect of IL-10AS on the percentage of IFN-γ+ cells in T-cell subsets (CD4+ and SWC6+). PBMCs were transfected with IL-10AS at 2 μM for 4 h, then cultivated with PRRSV for 48 h, and stimulated with PMA/I/G for another 12 h (IL-10AS+PRRSV). T-cell subsets from these PBMC cultures were gated and assessed for the percentage of IFN-γ+ cells by flow cytometry (CD8β+IFN-γ+). Controls included T-cell subsets from the following PBMC cultures: PBMCs treated with culture media alone and stimulated with PMA/I/G (positive control; POS CTRL); PBMCs treated with culture media plus mock antigens and stimulated with PMA/I/G (mock control; Mock); PBMCs treated with culture media plus PRRSV and stimulated with PMA/I/G (PRRSV-inoculated control; PRRSV); and PBMCs treated with transfection media (no IL-10AS) plus PRRSV and stimulated with PMA/I/G (PRRSV-inoculated transfection media control; Transfection media+PRRSV). Error bars indicate the standard error of the mean. Asterisks represent a significant mean difference in the percentage of IFN-γ+ cells between treatment and positive control groups (p < 0.05). The same letter (a) indicates no significant mean difference in the percentage of IFN-γ+ cells between the transfected and PRRSV-inoculated control groups (p > 0.05).
Compared to the lymphocyte population and CD8β+ T cells from PRRSV-inoculated PBMCs, transfection of PBMCs with IL-10AS prior to PRRSV inoculation and PMA/I/G stimulation resulted in an increased percentage of IFN-γ+ cells in lymphocytes (p = 0.11), and CD8β+ T cells (p = 0.068; Figs. 4 and 5). No increase in the percentage of IFN-γ+ cells was detected in lymphocytes and T-cell subsets of the PRRSV-inoculated transfection media controls (Fig. 4 and 5).
Discussion
The present study investigated the potential of IL-10AS in downregulating IL-10 mRNA expression in PBMCs inoculated with PRRSV and stimulated with a cytokine inducer (PMA/I or ConA). The study also evaluated the effects of IL-10AS on proinflammatory cytokine mRNA expression and percentages of IFN-γ+ cells in inoculated PBMC and T-cell subsets.
Increased IL-10 production has been proposed to be a strategy of PRRSV to suppress innate and adaptive immune responses of infected pigs. PRRSV-enhanced IL-10 production has been reported to correlate with suppressed IFN-γ production in virus-infected cells (6,44,49,50). In humans and mice, IL-10 has been reported to suppress proinflammatory cytokine expression and CMI response (13,14,16,17,26,36,39,51,52,61). In pigs, the cytokine has been reported to suppress IFN-γ production in PBMCs and T cells (6,58).
The IL-10AS evaluated in the present study has a phosphorothioate backbone. Theoretically, phosphorothioate-modified antisense ODN controls target mRNA expression by binding specifically to target mRNA, and forming mRNA/DNA duplexes that trigger RNaseH to cleave hybridized target mRNA (45,48). This results in reduced amounts or a complete absence of intact mRNA templates for translation, and thereby reduced target protein production (45,48).
The potential of IL-10AS in downregulating IL-10 mRNA expression induced by PRRSV is shown in Fig. 2. This finding suggests that IL-10AS may be useful as a tool for controlling IL-10 expression due to PRRSV infection.
The contribution of IL-10AS to enhancing proinflammatory cytokine response to PRRSV has been observed only for IFN-γ expression. Transfection with IL-10AS contributed to increasing IFN-γ mRNA expression in PRRSV-inoculated PBMCs (Fig. 3). The increased IFN-γ mRNA expression seen in response to IL-10AS was in accordance with several reports in humans and mice, that IL-10 reductions led to increased IFN-γ expression (9,24,28,30,47). This observation indicates the potential of IL-10AS in enhancing the IFN-γ response to PRRSV.
In contrast to IFN-γ mRNA expression, transfection with IL-10AS did not contribute to increasing IFN-α mRNA expression in PRRSV-inoculated PBMCs (Fig. 3). To date, the mechanism behind PRRSV's IFN-α suppression is not known. The finding that IL-10AS transfection makes no contribution to enhancing IFN-α expression suggests that PRRSV might not utilize IL-10, at least solely, for this cytokine suppression. Future study is needed to elucidate exactly how PRRSV suppresses IFN-α.
IL-10AS transfection unexpectedly suppressed IL-2 mRNA expression in PRRSV-inoculated PBMCs (Fig. 3). Previous studies regarding the relationship of IL-10 and IL-2 reported that these two cytokines usually have an inverted relationship (26,39,51,52). The finding that IL-10AS downregulated IL-2 expression was thus not expected. One possible explanation may be that IL-10AS interfered with PRRSV's mechanism of IL-2 induction. This was probably due to IL-10AS binding to and facilitating degradation of PRRSV-induced mRNA molecules that signal for IL-2 induction. Such mRNA molecules should not be upregulated by ConA alone, since no IL-2 reduction was detected in ConA-stimulated IL-10AS-transfected PBMCs. At present, the mechanism behind PRRSV's IL-2 induction in ConA-stimulated naïve PBMCs is not known. Further investigations are needed to determine this mechanism, as well what causes IL-10AS-mediated IL-2 suppression.
In addition to evaluation at the mRNA level, the contribution of IL-10AS to enhancing the proinflammatory cytokine response to PRRSV was also evaluated at the protein level. In this study, we determined the cytokine production by using a flow cytometry-based assay. The reason for using this type of assay was because it allowed us to obtain more information on lymphocytes, and each T-cell subset's response to PRRSV and IL-10AS than other types of assays (e.g., ELISA and ELISPOT). This assay, however, has a limitation on the availability of mAbs that are applicable to it that are specific for porcine cytokines. For this study, the mAb specific for IFN-γ seems to be the only mAb proven effective for this application (6,7,42). Thus we evaluated only the contribution of IL-10AS to IFN-γ production.
Like IFN-γ mRNA expression, transfection of PRRSV-inoculated PBMCs with IL-10AS contributed to increasing percentages of IFN-γ+ cells in co-cultivated lymphocytes and CD8β+ T cells (Figs. 4 and 5). This finding was in accordance with a previous report that neutralization of PRRSV-induced IL-10 resulted in enhanced percentages of IFN-γ+ cells in T cells cultivated with the virus (6). Based on the reported function of CD8β+ T cells as cytotoxic T cells (5,23,40), this finding suggests that IL-10AS may have potential in enhancing the cytotoxic T-cell response to PRRSV.
Unlike CD8β+ T cells, CD4+ and SWC6+ T cells did not demonstrate reduced percentages of IFN-γ+ cells when cultivated with the virus (Fig. 5). This finding suggests that these two T-cell subsets are less susceptible to PRRSV suppression than CD8β+ T cells. The rationale behind this is not known. This assumption was supported by a previous report that CD4+ is the predominant T-cell subset responsible for IFN-γ production following PRRSV infection (34).
This study reports new evidence about the IFN-γ response of SWC6+ T cells to PRRSV. The IFN-γ response of this γδ T-cell subset to PRRSV has never before been studied. As shown in Fig. 5, SWC6+ T cells produced some IFN-γ following PRRSV inoculation. The percentage of IFN-γ+ cells detected was low, but it corresponded to that previously reported in peripheral blood γδ T cells (6). As the role of SWC6+ T cells in PRRSV protection is currently unclear, this finding suggests that these cells may make some contribution to protection against PRRSV.
It should be noted that the effects of IL-10AS seen in the present study were determined from the transfection of the entire PBMC population. Thus the precise cell subpopulations and the percentages that were efficiently transfected with IL-10AS are not known. This information is essential, since it may affect the magnitude of the proinflammatory cytokine response to PRRSV. Efficient transfection of IL-10-producing cells (e.g., PRRSV-infected monocytes and co-cultivated T cells) would reduce overall IL-10 expression in the culture, probably to levels at which proinflammatory cytokine expression can be upregulated. On the other hand, inefficient transfection of these cells would probably reduce only some IL-10 expression, but probably not to a level at which an increased proinflammatory cytokine response can be triggered. Since this study lacks data about the exact transfected PBMC subpopulations and their transfection percentages, we do not know whether or not the results observed were affected by differential IL-10AS transfection of PBMC subpopulations. Future study is needed to determine the transfected cell types, and their levels of efficiency of IL-10AS transfection of PRRSV-inoculated PBMCs.
It is worth noting that the present study was conducted in vitro. Thus many potential cell interactions that may affect the effectiveness of IL-10AS in vivo may not be present. These cell interactions are, for example, PRRSV-infected DC and PRRSV-infected macrophage interactions with cytotoxic T cells. In vivo, PRRSV-infected DCs and macrophages are professional APCs that prime and activate PRRSV-specific cytotoxic T cells. These APCs are reportedly capable of expressing IL-10 following PRRSV infection, and thus they are capable of downmodulating cytotoxic T-cell differentiation (4,6,8,20 –22,25,29,38). These cells may need to be transfected with IL-10AS in vivo in order to control their IL-10 expression. In addition to PRRSV-infected DCs and macrophages, IL-10AS may also need to transfect regulatory T cells, since these cells may also contribute to increased IL-10 production following PRRSV infection (60).
When applied in vivo, IL-10AS may be utilized as a vaccine adjuvant for a PRRSV-modified live-virus vaccine, which has been reported to elicit a delayed CMI response (63). For this, the IL-10AS would be expected to help reduce APC production of IL-10, and contribute to accelerating and magnifying the proinflammatory cytokine response of cytotoxic T cells, as well as of the APCs themselves.
In conclusion, in the present study we report about the potential of IL-10AS for downregulating IL-10 mRNA expression, and in upregulating IFN-γ mRNA expression, as well as in enhancing the percentage of IFN-γ+ cells in lymphocytes and CD8β+ T cells. These findings suggest the efficacy of IL-10AS for potentiating the overall immune response, and particularly the cell-mediated immune response, to PRRSV.
Footnotes
Acknowledgments
The authors thank Drs. James A. Roth and Ratree Platt at Iowa State University and Dr. Dale E. Taneyhill at Chiang Mai University for critical review of the manuscript. The authors thank Ms. Reunkaew Praphrute for excellent laboratory assistance. This work is supported by the Thailand Research Fund (grant no. MRG5080167), the Commission on Higher Education, and the National Science and Technology Development Agency (the NSTDA Research Chair Grant).
Author Disclosure Statement
No competing financial interests exist.