
Abstract
Despite a safe and effective vaccine, endemic rubella remains a problem in developing countries. Isolated cases and outbreaks can occur in areas with high vaccine coverage. Individuals, especially pregnant women who remain unimmunized or do not seroconvert, are susceptible to infection and their infants are at risk for congenital rubella syndrome (CRS). Both humoral and cellular immune responses contribute to immune protection. Classically, immunity to rubella has been assessed through the detection of rubella-specific antibody titers. In this study we examined correlates of both humoral and cellular immunity in a large population of immunized young adults in Olmsted County, MN. We were unable to find any significant correlation between cytokine production after in-vitro rubella stimulation and serum antibody titers.
Introduction
Natural infection with rubella as well as rubella vaccination induces both humoral and cellular immune responses that together confer protection from re-infection. While humoral responses have been conventionally used to measure protective immunity, cellular immune responses provide a crucial counterpart to humoral immunity (3,8,13,23), but have not been well studied in the case of rubella (1,2,10,18 –22,37,38). Investigators pursuing novel vaccines have raised the importance of cell-mediated immunity (CMI) in protection from rubella infection. The humoral and cellular arms of the adaptive immune system are closely intertwined, and cytokine production by T lymphocytes is integral to the development of effective antibody responses (5).
Several broad patterns of cytokine production can be distinguished: proinflammatory cytokines such as TNF-α or IL-6 play critical roles early in infection, Th-1 responses characterized by IL-2 or IFN-γ drive robust cytotoxic T-cell activity, and Th-2 responses defined by the production of IL-4, IL-5, IL-10, and IL-13 shape humoral immunity (27,32). To help inform further rubella vaccine development and to better understand immunologic markers of protection from rubella infection after vaccination, our objective was to identify possible associations between cytokine measures of cell-mediated immunity and rubella antibody level.
Materials and Methods
Subjects
The subjects for this study were recruited from Olmsted County, Minnesota, where vaccination rates are high and there have been no cases of natural rubella infection in the community during the lifetime of the participants (33). The study subjects were derived from two stratified, random samples of healthy children and young adults enrolled in private or public school in Olmsted County. In all, 342 healthy children between 12 and 18 y of age were enrolled between December 2001 and August 2002, while an additional 396 children (11–19 y old) were recruited between December 2006 and August 2007. The study was approved by the Mayo Clinic Institutional Review Board, and informed written consent and assent was obtained from all participants and their guardians (in the case of minors) prior to enrollment. The medical record of each participant was reviewed for documentation of receipt of two age-appropriate doses of measles, mumps, and rubella virus vaccine, live (M-M-R® II; Merck and Co., Inc., West Point, PA), and each participant was in good health at the time of enrollment. Once enrolled, a single blood sample was obtained from each of the study participants.
Rubella IgG enzyme immunoassays
For each subject, serum aliquots were stored at −80°C until use. The samples were thawed and virus-specific rubella IgG levels were determined using an automated paramagnetic particle, chemiluminescent immunoassay (Beckman Coulter, Fullerton, CA), according to the manufacturer's protocol. The assays were performed using a UniCel DxI 800 Access Immunoassay System (Beckman Coulter). WHO reference standards for anti-rubella serum were used to determine the antibody titer for each sample. For this assay the limit of detection was 0.5 IU/mL, and the coefficient of variation in our laboratory was 6%.
Preparation of peripheral blood mononuclear cells
Peripheral blood mononuclear cells (PBMC) were isolated from venous blood using Ficoll-Hypaque gradient (Sigma, St. Louis, MO) centrifugation, and washed in RPMI medium with glutamine (Gibco, Carlsbad, CA) supplemented with 5% heat-inactivated fetal calf serum (Hyclone, Logan, UT), 1 mM sodium pyruvate (Cellgro, Manassas, VA), 100 μg/mL streptomycin, and 100 U/mL penicillin (Sigma). The cells were counted and resuspended in freezing media with dimethyl sulfoxide, and 1-mL aliquots containing 106 cells were stored in liquid nitrogen until needed.
Cell culture and stimulation with rubella virus
An aliquot of PBMC for each subject was thawed and resuspended in complete RPMI 5% FCS (supplemented with pyruvate, streptomycin, and penicillin as above). Trypan blue exclusion was performed to determine cellular viability and no difference was found between samples before and after storage in liquid nitrogen. Duration of incubation and MOI of virus for each assay was optimized as previously described (28). Briefly, 2 × 105 cells for each subject in RPMI 5% FCS was added to seven wells of a 96-well plate for each of the cytokines studied. Rubella virus, Wistar RA 27/3 strain (Meruvax; Merck & Co.), in complete RPMI 5% FCS was added to three of the seven wells for each subject for each cytokine studied, to achieve an MOI of 0.05 for TNF-α assays, and an MOI of 5 for the IL-2, IL-4, IL-5, IL-6, IL-10, IL-12p40, GM-CSF, and IFN-γ assays. For each subject, PHA (Sigma) was added to one well for each cytokine studied to a concentration of 5 μg/mL. Three of the seven wells were left as unstimulated controls for each subject for each cytokine studied. The plates were then incubated at 37°C and 5% CO2 for 18 h for the GM-CSF and IL-12p40 cytokine assays, 24 h for the IL-4, IL-5, IL-6, and IL-10 cytokine assays, 2 d for the IFN-γ cytokine assays, and 8 d for the IL-2 and TNF-α assays. The supernatants were then transferred to a new 96-well plate, covered, and placed in a −80°C freezer until the time the ELISA was performed.
Cytokine response by enzyme-linked immunosorbent assay
The 96-well plates were thawed and commercially available ELISA kits (BD Biosciences, San Jose, CA) were used according to the manufacturer's instructions and with the manufacturer's standards to assay the supernatants for concentrations of IL-2, IL-4, IL-5, IL-6, IL-10, IL-12p40, GM-CSF, IFN-γ, and TNF-α.
Cytokine response by ELISPOT
Commercial ELISPOT kits for IL-10 (BD Biosciences) and IFN-γ (R&D Systems, Minneapolis, MN) were used as previously described (29). For both cytokine assays, ELISPOT plates were coated overnight at 4°C with capture antibody, then washed and blocked for 2 h. PBMC were thawed and counted. Duration of incubation and MOI of virus for each assay was optimized as previously described (28). After resuspension in RPMI 5% FCS, 2 × 105 cells were added to seven wells for each subject for each assay. Rubella virus in RPMI 5% FCS was added to three wells for each subject for each assay (MOI of 2.5). PHA in RPMI 5% FCS was added as a positive control to one well for each subject for each assay (final concentration of 5 μg/mL). For each subject, three wells remained as unstimulated controls. The plates were then placed in a 37°C 5% CO2 incubator for 24 h.
For the IL-10 ELISPOT assay, the cells were removed and detection antibody was added to each well at room temperature for 2 h. The detection antibody was discarded and streptavidin-HRP was added to each well for 1 h at room temperature and then removed. TMB-H substrate (Moss Inc., Pasadena, MD) was added to each well for 30 min at room temperature in the dark. The plates were then washed extensively and dried prior to counting. For the IFN-γ ELISPOT assay, following the 24-h 37°C incubation, the cells were removed and detection antibody was added to each well and incubated at 4°C overnight. Detection antibody was then discarded and streptavidin-AP was added for 2 h at room temperature, then removed. BCIP/NBT chromogen was then added to each well for 1 h at room temperature in the dark. The plates were then washed extensively and dried prior to counting.
An ImmunoSpot S4 Pro Analyzer from C.T.L. (Cleveland, OH) was used to count the ELISPOT plates. Counting parameters were created separately for IL-10 and IFN-γ. The parameters included sensitivity, spot size threshold, background balance, and spot separation. A random selection of subjects was used to create the initial counting parameters for IL-10 and IFN-γ, and then held constant for all ELISPOT plates for both cytokines. Quality control was performed by a single operator for each plate, removing debris and manually excluding overdeveloped areas.
Statistical analysis
The purpose of the efforts reported here was to assess associations between rubella humoral and cellular immune response measures. The following outcomes were examined: levels of rubella-specific IgG antibody titers (measured in IU/mL), nine measures of rubella virus-specific in-vitro cytokine secretion (IL-2, IL-4, IL-5, IL-6, IL-10, IL-12p40, IFN-γ, TNF-α, and GM-CSF, each reported in pg/mL), and two measures of CMI via rubella vaccine-induced memory T-cell frequencies (IFN-γ and IL-10, evaluated as count variables). Assessments of cytokine secretion and CMI resulted in six recorded values for each of the outcomes of interest per individual: three prior to stimulation with rubella virus and three post-stimulation. For these outcomes, a single response measurement per individual was obtained for each outcome by subtracting the median of the three unstimulated values from the median of the three stimulated values. Assessments of antibody levels resulted in only one recorded value per individual. Data were descriptively summarized across individuals using frequencies and percentages for categorical variables, and medians, ranges, and inter-quartile ranges for continuous variables.
Linear associations between antibody levels and measures of cellular immune response were formally assessed using correlation coefficients and corresponding 95% confidence intervals. To correct for the inherent data skewness found in each of the immune response measures, Spearman (rank-based) correlations were used throughout. Primary analyses were based on simple, unadjusted correlation coefficients. Secondarily, we calculated partial correlation coefficients to evaluate associations after adjustment for the potential confounding effects of age at enrollment, gender, race, age at first rubella vaccination, age at second rubella vaccination, and cohort status (cohort 1 versus cohort 2). Finally, we re-assessed associations with cellular immunity by examining the extremes of antibody response. We compared distributions of cytokine secretion and ELISPOT measures in subjects with low antibodies (<25 IU/mL) and high antibodies (≥75 IU/mL) using Wilcoxon rank sum tests. Analyses were carried out using SAS version 9 (SAS Institute, Inc., Cary, NC) and S-Plus version 8 (Insightful Corporation, Seattle, WA) software systems.
Results
Our study population of healthy school-age children and young adults were between 11 and 19 y of age. The gender breakdown was 54% male and 46% female, and most of the subjects were Caucasian (91%). The median age at first and second rubella immunization was 11 mo and 15 y, respectively. We tested serum rubella-specific IgG titers as well as cytokine secretion (IL-2, IL-4, IL-5, IL-6, IL-10, IL-12p40, GM-CSF, TNF-α, and IFN-γ) following in-vitro rubella stimulation.
Measures of rubella-specific immunity
Rubella-specific antibody levels ranged between 2.5 and 329.8 IU/mL. The median antibody level was 34.40 IU/mL (IQR 19.2, 63.7), with 87% of subjects having an antibody level ≥15 IU/mL (Fig. 1). Secreted levels of the nine cytokines tested are described in Table 1. There was a robust IL-6 response (median 3680.99 pg/mL), moderate IL-2, GM-CSF, and TNF-α responses (median 17.59 pg/mL, 28.04 pg/mL, and 29.74 pg/mL, respectively), minimal IFN-γ and IL-10 responses (median 8.53 pg/mL and 4.20 pg/mL, respectively), and negligible IL-12p40, IL-4, and IL-5 responses (median 0.00 pg/mL, 0.30 pg/mL, and 0.47 pg/mL, respectively) to rubella stimulation. In addition to cytokine measures we also assessed T-cell responses by IFN-γ and IL-10 ELISPOT assays. As we detected minimal levels of IFN-γ and IL-10 by ELISA, we expected to see similarly modest numbers of spot-forming cells in our ELISPOT assays. IFN-γ ELISPOT responses ranged from −93 to 23, while IL-10 spot-forming counts ranged from −458 to 310. In agreement with the minimal cytokine secretion detected by ELISA, after subtracting background levels of cytokine production from the unstimulated control wells, 52.4% of subjects had positive IL-10 responses, and very few (19.6%) had positive IFN-γ responses. The negative values for the ELISPOT data are a result of subtracting the background levels of cytokine spot-forming cells in the unstimulated wells. In these individuals stimulation with rubella virus led to a decrease in cytokine production compared to baseline levels.
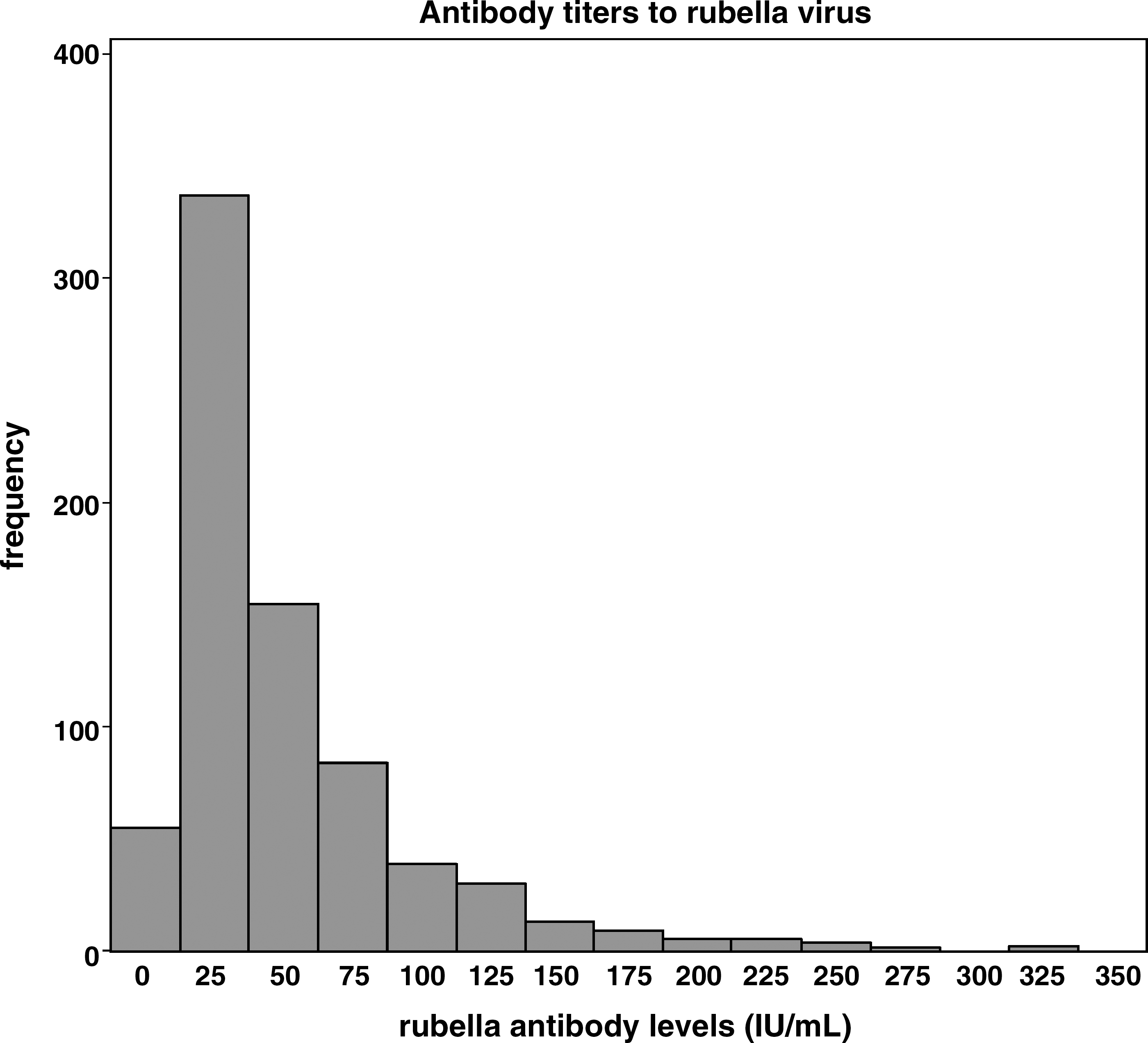
Antibody titers to rubella virus.
IQR, interquartile range.
Correlation between antibody and CMI measures
We found a striking lack of correlation between cytokine production after in-vitro rubella stimulation and serum antibody titers (Table 2). None of the cytokines had significant correlation with antibody response after accounting for age at enrollment, gender, race, age at first rubella vaccination, age at second rubella vaccination, and cohort status (partial correlation). Without adjusting for these variables (simple correlation), IL-6 and IFN-γ had significant correlations with antibody response, but the correlation coefficients (0.078 and 0.077, respectively) were lower than 0.1, consequently it is difficult to assign biological or clinical importance to these differences. Further analysis comparing those who had low antibody levels (<25 IU/mL, n = 255) with those with high antibody levels (≥75 IU/mL, n = 137) did not reveal any significant differences in cytokine response between the two groups (data not shown).
Spearman correlation coefficients. Simple correlations are unadjusted. Partial correlations account for age at enrollment, gender, race, age at first rubella vaccination, age at second rubella vaccination, and cohort status.
Discussion
The continued incidence of CRS globally, occasionally in the face of detectable humoral immunity against rubella, highlights the need for a deeper understanding of markers of protective immune response to vaccination, and the need for new vaccine development. The importance of cellular immunity to the development of protection against other viral pathogens such as herpes simplex virus, influenza virus, varicella virus, and hepatitis B virus has been demonstrated (14,15,17,34). The role of cellular immunity in rubella is as yet poorly defined. One of the goals of the present study was to more clearly characterize cell-mediated immune responses to rubella virus, and to determine potential associations between humoral and cellular immune responses to rubella vaccination. Vaccine success and rubella immunity is typically assessed by quantitation of rubella-specific antibody titers, or even neutralizing antibody activity against the virus. Far less is known about the role of cellular immunity, including cytokine production, in protection against rubella infection.
In this study of 738 adolescent Olmsted County children with documentation of two rubella vaccinations, the vast majority (87%) of subjects demonstrated protective rubella antibody levels (≥15 IU/mL) (31). PBMC from these subjects had robust IL-6 cytokine secretion, and moderate GM-CSF and TNF-α secretion following stimulation with rubella virus. The cytokine secretion assays suggested a stronger Th-1 response than Th-2 response, with moderate IL-2 and IFN-γ response, and negligible IL-12p40 response, compared to a minimal IL-10 response and a negligible IL-4 and IL-5 response. Our results indicate that cytokine production by memory T cells in response to rubella virus is fairly weak and is skewed toward the production of inflammatory and Th-1-type cytokines, with relatively little production of Th-2 cytokines such as IL-4, IL-5, and IL-10. This Th-1 skewing has also been seen in mouse models of rubella infection (10).
We found minimal correlations between markers of cellular immunity, as measured by cytokine secretion and serum antibody levels. For the only cytokines in which there was a significant correlation, IL-6 and IFN-γ, the correlation coefficients were lower than 0.1, casting doubt on the biological relevance of these correlations. Furthermore, the correlations were no longer significant after accounting for age at enrollment, gender, race, age at first rubella vaccination, age at second rubella vaccination, and cohort status. This is similar to what we have previously reported for measles (24), but contrasts with other studies examining individual T-cell epitopes and neutralizing antibody titers, or total T-cell numbers and humoral immunity (HI) titers (16,35).
There are several potential reasons why no associations were found between markers of humoral and cellular immunity following rubella vaccination. We studied total IgG levels, and it may be that cytokine production could be correlated with individual isotype levels. A finer dissection of both humoral (HI versus neutralizing) and cellular (quantitation of rubella-specific T-cell numbers and individual epitope responses) may be necessary to identify associations between humoral and cellular immunity to rubella. It is possible that the level of cytokine response to vaccination was too low to detect statistically significant differences because of the timing of the study in relation to when subjects had received their rubella vaccine.
It is known that humoral and cellular immune processes are integrally linked with each other during the initial induction of immunity to a pathogen. Cytokines are involved in immunoglobulin isotype switching and controlling B-cell development and differentiation. The presence of protective antibodies can affect antigen concentration and reduce subsequent antigen presentation. As time passes following initial antigenic challenge, however, humoral and cellular memory responses may not be interconnected nearly as much as primary responses. In fact, B and T memory cells use different survival factors and as such, B-cell and T-cell immunity decline at different rates, and individual cell lifespans may differ. This disconnect between memory T cells and neutralizing Ab titers has been seen with other viral pathogens such as vaccinia and hepatitis B (6,11). To better discern correlations between humoral and cellular markers of immunity to rubella at the time of initial antigenic challenge, a subsequent study should be conducted shortly after initial rubella vaccination, when primary T- and B-cell responses are at their peak.
Currently, the primary measure of rubella vaccine efficacy and protection is antibody titer. Our data suggest that measuring cytokine secretion after ex-vivo virus stimulation several years after vaccination will not provide any additional information on vaccine efficacy. Similarly, the low levels of cytokines detected in our cohort may indicate waning of cellular immunity. Although our data did not show correlations between cytokine response and antibody response to rubella vaccination, a deeper understanding of the relationship between cytokine response and antibody response to pathogens is integral to the future use of cytokines as markers of immunity or as potential adjuvants to vaccines, and may inform new vaccine development in directing cellular responses in order to optimize vaccine efficacy. In addition, negative studies such as this do provide useful insights and inform the next series of experiments designed to further advance our rubella immunology knowledge base.
Footnotes
Acknowledgments
We would like to thank the clinical nursing and study coordinators of the Mayo Vaccine Research Group for their efforts in recruitment, obtaining consent, and biospecimen acquisition. We also acknowledge the kind assistance of the Rochester, Minnesota public school district, and the support of the Rochester Epidemiology Project and the Mayo Center for Translational Science Activities. This work was supported by National Institutes of Health (NIH) grants AI 48793, AI 33144, and 1 UL1 RR024150-01 for the National Center for Research Resources (NCRR), a component of the NIH, and the NIH Roadmap for Medical Research. Its contents are solely the responsibility of the authors and do not necessarily represent the official view of the NCRR or the NIH.
Author Disclosure Statement
Dr. Poland is the chair of a Data and Safety Monitoring Board for novel non-rubella vaccines undergoing clinical studies by Merck Research Laboratories. Dr. Jacobson serves on a Safety Review Committee for a post-licensure study of Gardasil for Kaiser-Permanente. No competing financial interests exist for the other authors.