
Abstract
We investigated in-vitro lymphoproliferative responses and T-cell subsets in 38 HIV-1-infected patients showing impaired restoration of CD4 T cells despite prolonged viral suppression (discordant), and in 42 HIV-1-infected patients showing positive immunological and virological responses to highly active antiretroviral therapy (HAART) (concordant). In comparison to concordant patients, discordant patients showed poor lymphocyte proliferation, lower secretion of IL-2 and IFN-γ, a lower percentage of perforin and granzyme-B-producing CD8 T cells, and poor differentiation of effector memory CD8 TEM cells into CD8 TEMRA cells in in-vitro stimulation assays, especially against HIV-1 Gag p24 and one of its peptide pools. Functional CD8 T-cell responses of discordant patients after stimulation with recall antigens, Candida albicans, and tetanus toxoid, were also inferior to concordant patients, but comparable to normal healthy controls. Examination of the multifunctional roles of T cells is imperative in describing the overall magnitude of immune responses to HIV-1. Our results suggest that prolonged suppression of plasma viremia alone does not warrant good qualitative and quantitative CD8 T-cell responses to HIV-1, implying that CD4 T cells are required for maintenance of protective CD8 T-cell responses.
Introduction
Insufficient CD4 T-cell numbers can influence the function of CD8 T cells that play an important role in the control of HIV infection and generation of secondary responses to infectious pathogens. On first encounter with viral epitopes, naïve CD8 T cells are capable of generating effector memory (TEM) and central memory (TCM) cells with the aid of CD4 T-helper cells (5,6). Upon subsequent re-encounter with the same antigen, most of the cells from the TEM pool differentiate into CD45RA-expressing TEM (TEMRA) cells, most of which are destined to become effector cells, while some continue to stay in their TEM phenotype (7,8). Antigen-specific memory CD8 T-cell protection is characterized by the ability of CD8 TEM cells to produce a robust secondary response by proliferating rapidly to CD8 TEMRA cells, that are further capable of producing potent effector molecules such as perforins, granzymes, IFN-γ, and TNF-α (9 –12). Though it has been recently demonstrated that CD8 T cells can be primed without CD4 T-cell help, they are incapable of mounting a robust secondary response and die upon re-encountering antigen (6,13). Several studies have also reported selective accumulation of immature HIV-1-specific CD8 memory T cells during HAART, suggestive of inadequate CD4 T-cell help or HIV-1-specific defects (14 –16).
Recently, we observed a group of discordant patients that failed to show any increase in their mean CD4 T-cell counts despite maintaining low to undetectable plasma viral loads for more than 1 y (17). In comparison to the patients that showed good (concordant) immunological and virological response to HAART, the discordant patients had significantly higher numbers of CD8 T cells, and higher percentage of memory CD4 and CD8 T cells. In this study we evaluated the function of CD4 T cells and lymphoproliferative abilities in in-vitro cultures stimulated with HIV-1-specific antigens and recall antigens, Candida albicans, and tetanus toxoid, to understand whether insufficient CD4 T-cell numbers alter functional CD8 T-cell responses in discordant HIV-1-infected patients. The discordant patients showed poor lymphocyte proliferation and fewer perforin- and granzyme-B-producing CD8 T cells following stimulation with HIV-1 Gag p24 and one of its peptide pools compared to concordant patients. At the same time T-cell responses of discordant patients after stimulation with C. albicans and tetanus toxoid were comparable to those of normal healthy controls. When we analyzed variations in in-vitro proliferation of different CD4 and CD8 T-cell subsets, the discordant patients demonstrated an increase in relative percentage of CD8 TEM cells, but poor differentiation into CD8 TEMRA cells, particularly in Gag p24-stimulated cultures.
Materials and Methods
Study subjects and specimens
Peripheral whole blood specimens were collected in sodium heparin and EDTA tubes from 80 HIV-1-infected patients and 12 age- and sex-matched HIV-negative subjects (normal healthy controls) from the University of Miami–Miller School of Medicine outreach clinic, Project Outreach, Florida City, Florida, and Borinquen Health Care Center, Miami, Florida. The whole-blood samples of the recruited subjects were submitted to the Laboratory for Clinical and Biological Studies (LCBS), where the all investigations were performed. We observed the pattern of CD4 and CD8 T-cell counts and plasma HIV-1 RNA levels (recorded every 4–6 mo for over 2 y) of 1100 patients to screen and recruit study subjects that demonstrated a strict discordant or concordant profile and divided them into two groups: discordant (n = 38) and concordant (n = 42). The study was duly approved by the Institutional Review Board of the University of Miami. The criteria for selection of discordant patients were: undetectable plasma HIV-1 RNA (<50 copies/mL) for at least 1 y, with (a) whole-blood CD4 T-cell count <200/mm3, or (b) CD4:CD8 ratio <1.0 with CD4 T-cell counts <400/mm3 and CD4 T-cell increase not greater than 100/mm3 following HAART (Fig. 1) (1,2). Patients showing positive immunological and virological response following HAART (a CD4 T-cell increase >100/mm3 following HAART) with virological control were placed in the concordant group. Patients were excluded for non-adherence to HAART, any acute illness, opportunistic infections, lymphomas, or psychiatric illness.
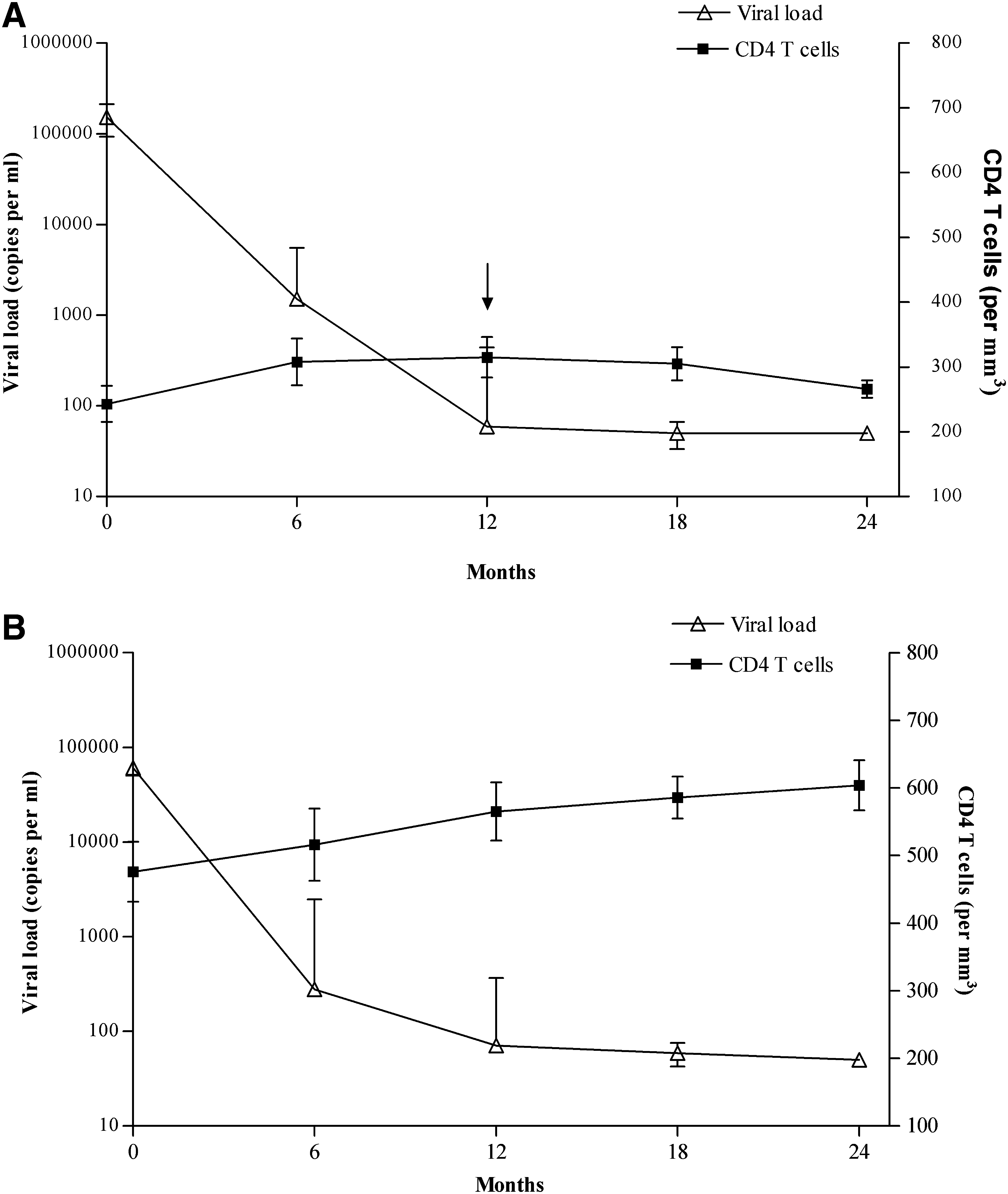
Pattern of CD4 T-cell counts (mean ± SD) and plasma viral load of (
Lymphoproliferative responses and antigens
The white blood counts and percentage of lymphocytes in patient samples were obtained using a Coulter AcT5 5-part differential hematology analyzer (Beckman Coulter, Fullerton, CA). Peripheral blood mononuclear cells (PBMCs) were isolated from heparin-preserved fresh whole blood by density gradient centrifugation with Ficoll-paque Plus™ lymphocyte separation medium (Amersham Biosciences AB, Uppsala, Sweden), washed twice in phosphate-buffered saline, and resuspended in RPMI 1640 with glutamine (Mediatech Inc., Herndon, VA).
A total of 105 PBMCs/well were incubated in quadruplicate in 200 μL of RPMI 1640 medium supplemented with L-glutamine, 10% heat-inactivated human AB serum (Mediatech Inc.), and 1% penicillin, streptomycin (Invitrogen Corp., Carlsbad, CA) in 96-well U-bottom plates (Nunc, Rochester, NY). Negative control wells contained medium alone and cells without any stimulant, while positive control wells contained 5 μg/mL of mitogen, phytohemagglutinin (PHA) (Sigma-Aldrich, St Louis, MO). Test wells contained recall antigens, 10 μg/mL Candida albicans (Greer Laboratories, Lenoir, NC), and 5 μg/mL tetanus toxoid (Sigma-Aldrich), and HIV antigens, 2.5 μg/mL of recombinant HIV-1 Gag p24 antigen (ImmunoDiagnostics Inc., Woburn, MA), and two peptide pools of HIV-1 Gag p24 (pool 1, peptides ARP788.1–11 and pool 2, peptides ARP788.12–22). The peptide pools were comprised of 20mers with 10 overlapping amino acids spanning the complete Gag p24 region. The HIV-1 Gag p24 peptides were provided by the EU Programme EVA Centre for AIDS Reagents, NIBSC, U.K. (AVIP Contract Number LSHP-CT-2004-503487). After 6 d, the wells were pulsed with 1 μCi 3H-thymidine for 6 h. The cells were then harvested onto filter mats using a Tomtec Harvester 96 (Hamden, CT), and counted in a liquid scintillation counter (LKB Wallac 1205 Betaplate Liquid Scintillation Counter, Finland). A stimulation index (SI) of ≥3 was considered to be a positive response. A portion of the harvested cells was used for immunophenotyping of CD4 and CD8 T-cell subsets, including intracellular expression of perforin and granzyme-B in CD8 T cells.
Characterization of CD4 and CD8 T-cell subsets
Immunophenotyping of CD4 and CD8 T-cell subsets was done to analyze the expression of naive (CD45RA+, CCR7+), central memory (TCM; CD45RA−, CCR7+), effector memory (TEM; CD45RA−, CCR7−), and terminally differentiated TEMRA cells (CD45RA+, CCR7−), using specific fluorochrome-labeled monoclonal antibodies in two separate tubes (BD Biosciences, San Jose, CA). These included anti-CD3-APC-Cy7, CD4-PerCP, CD8-FITC, CD45RA-PE-Cy7, and CCR7-PE in tube 1, and anti-CD3-APC-Cy7, CD8-PerCP, CD45RA-APC, and CCR7-PE-Cy7 in tube 2. Isotype control antibodies were added to the control tubes and stained along with the test tubes. A total of 5 × 105 PBMCs were used for staining in each tube, followed by incubation in the dark for 30 min, and a wash with phosphate-buffered saline (pH 7.2). The cells were acquired on a six-color flow cytometer (FACSCanto™), and analyzed using FACSDiva™ software (v 4.1; BD Biosciences). A minimum of 100,000 events were acquired and analyzed for each specimen.
Measurement of intracellular perforin and granzyme-B
Following surface staining (in tube 2), intracellular granzyme-B and perforin were analyzed in the CD8 T cells using specific monoclonal antibodies and isotype control antibodies. The cells were permeabilized using the CytoPerm/CytoFix™ kit per the manufacturer's instructions (BD Biosciences), then stained for 30 min with anti-granzyme-B-FITC and perforin-PE antibodies. The cells were washed twice and suspended in the supplied wash buffer prior to analysis. Acquisition and analysis of the cells were carried out as described above.
Cytokine analysis
Cell-free supernatants from lymphocyte cultures were collected 24 h following stimulation with PHA, Gag p24, and C. albicans. Levels of IL-2 and IFN-γ were measured using a protein biochip array system, Evidence Investigator™ (Randox Laboratories Ltd., Crumlin, U.K.), as described previously (18). Supernatants from cultures without exogenous stimulant were used as controls.
Plasma viral load
HIV-1 RNA levels were determined in the plasma samples of HIV-1-infected patients by a quantitative RT-PCR assay, the Amplicor HIV-1 Monitor Test™ kit (v 1.5; Roche Diagnostic Corp., Indianapolis, IN), per the manufacturer's instructions. Plasma viral load below 50 copies/mL was considered undetectable.
Statistical analysis
The significance of differences between the mean values of all lymphocyte subsets was calculated by the independent samples T-test using SPSS software (v 14.0; SPSS, Inc., Chicago, IL). Welch's correction was applied to t-tests when normal healthy controls were compared to HIV-1-infected patients. Graphs showing comparison of lymphocyte subsets were plotted using GraphPad Prism software (v 4.0; GraphPad, La Jolla, CA).
Results
Patient characteristics and discordant/concordant responses
The discordant patients had similar durations of HIV-1 infection and HAART regimens compared to the concordant patients. However, the discordant patients had a significantly higher plasma viral load and lower CD4 T-cell counts than the concordant group at the initiation of HAART (p < 0.01). The trend of CD4 T-cell counts and plasma viral load of all the patients in the cohort was observed for a period of over 24 mo prior to their immunophenotyping (Fig. 1). The discordant patients demonstrated moderate increases in their CD4 T-cell counts from 0–12 mo. However, from 12 mo onwards, they failed to show any increase in their CD4 T cells, despite maintaining low-to-undetectable plasma viral load. In contrast, the concordant patients demonstrated a good immunological and virological response to HAART, with an inverse relationship between plasma viral load and their CD4 T-cell counts during the entire period of observation. Other demographic and clinical characteristics of patients and controls are described in Table 1.
Age, CD4 and CD8 counts, viral load, duration of infection, and HAART are expressed as mean ± 1 SD and range in parentheses.
Abbreviations: HAART: highly active antiretroviral therapy, NRTI: nucleoside reverse transcriptase inhibitor; NNRTI: non-nucleoside reverse transcriptase inhibitor; PI, protease inhibitor; NA, not applicable.
Discordant patients show poor lymphoproliferative responses to HIV-1 Gag p24
To investigate whether CD4 T cells are required for maintenance of memory T-cell responses, especially TCM responses, we analyzed lymphoproliferation of PBMCs following in-vitro stimulation with HIV-1-associated antigens. Of the various HIV-1-associated antigens, Gag p24 has been reported to be an important target for immune responses (19). Therefore, we cultured the PBMCs of both the groups of patients in the presence of complete recombinant HIV-1 Gag p24, and two peptide pools of HIV-1 Gag p24 (pool 1 and pool 2), separately.
Overall, 22 (57.9%) of the 38 discordant patients showed proliferation in response to Gag p24 antigen, while 33 (78.6%) of the 42 concordant patients showed good responses to Gag p24 (Table 2). Furthermore, when we compared the responders, the concordant patients showed a significantly higher proliferation of lymphocytes than the discordant patients (mean ± SD; SI = 16.8 ± 18.9 and 5.0 ± 3.1, respectively) (p < 0.05) (Fig. 2).
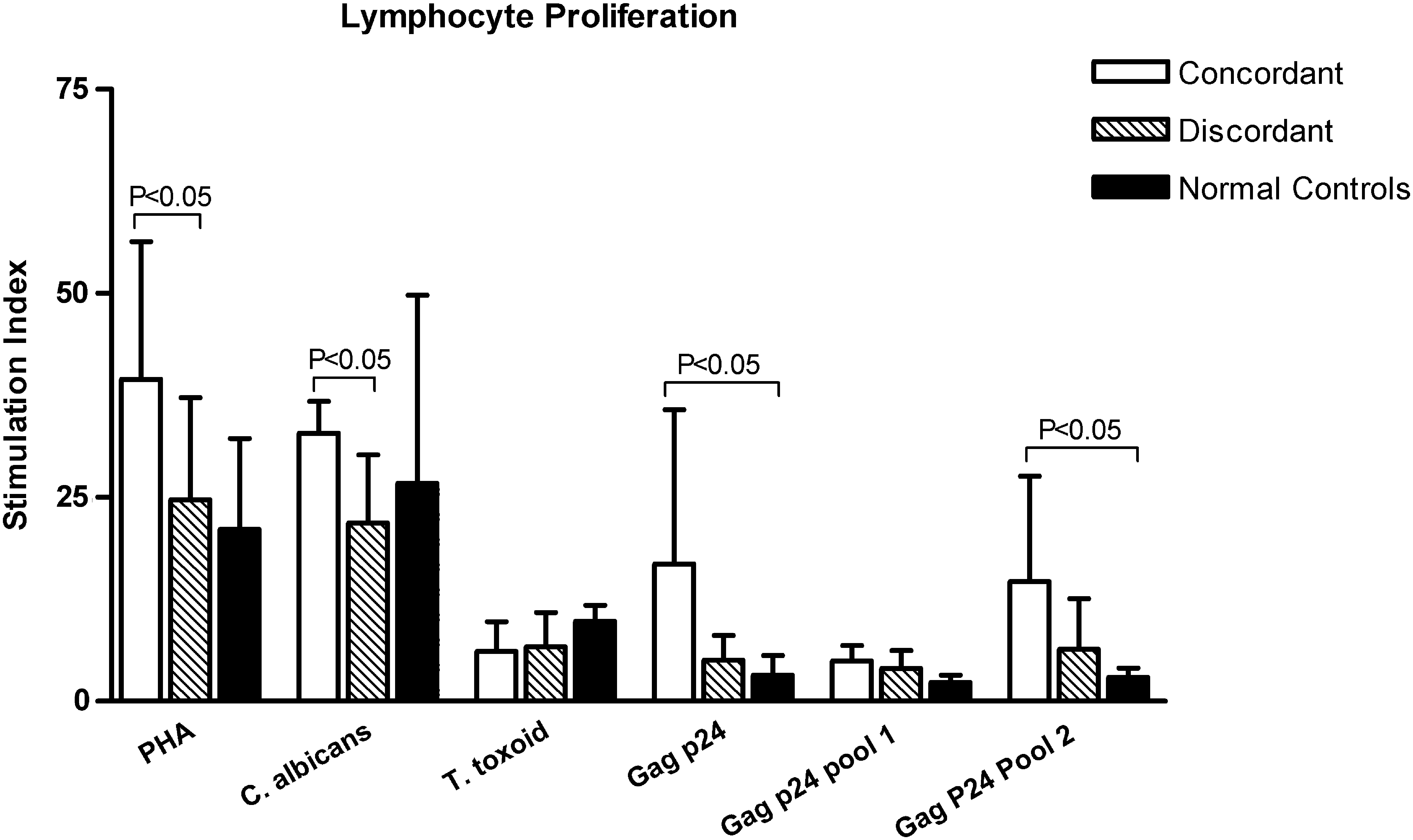
Proliferation (mean ± SD of stimulation indices) of peripheral blood mononuclear cells (PBMCs) isolated from discordant and concordant HIV-1-infected patients in the presence of various antigens. Phytohemagglutinin (PHA) was used as standard mitogen in all the assays, while no stimulant was added to the cells in the control wells. Discordant subjects had a significantly lower proliferation of PBMCs in the presence of Gag p24, peptide pool 2 of Gag p24, and C. albicans, compared to concordant subjects (p < 0.05).
SI, stimulation index.
In case of peptide pools of Gag p24, we observed that proliferative responses to pool 1 were relatively weak compared to pool 2 (p < 0.01). Only 5 (13.2%) of the discordant and 8 (18.2%) of the concordant patients responded to pool 1 of Gag p24 peptides (Table 2). The amount of proliferation following stimulation with pool 1 was also less than that of the Gag p24 antigen (mean ± SD; SI = 4.0 ± 2.2 and 4.9 ± 1.9 in discordant and concordant patients, respectively). However, the patients did show good responses to peptide pool 2 of HIV-1 Gag p24, indicating that the majority of the reactive p24 epitopes were in peptide pool 2. There were 20 (52.6%) responders in the discordant group and 33 (78.6%) responders in the concordant group. Just as in the case of Gag p24, the concordant patients showed a significantly higher proliferation of lymphocytes than the discordant patients (mean ± SD; SI = 14.7 ± 12.9 and 6.4 ± 6.2, respectively) following stimulation with peptide pool 2 of HIV-1 Gag p24 (p < 0.05) (Fig. 2). The normal healthy controls demonstrated poor proliferative responses to all HIV-1 antigens.
Lymphoproliferative responses to recall antigens are maintained in discordant patients
The majority of opportunistic infections are caused by the pathogens to which one is exposed during life. Therefore, besides using HIV-1-specific antigens, we used two types of recall antigens, C. albicans, to which the immune system is exposed regularly, and one with rare exposure, tetanus toxoid, as stimulants in the lymphocyte proliferation assays to ascertain whether the discordant patients were able to maintain protective cellular responses to these antigens. Upon stimulation with C. albicans, lymphocyte proliferation was observed in 27 (71.1%) discordant and 35 (83.3%) concordant HIV-1-infected patients (Table 2), and 11 (91.7%) normal healthy controls. The responses to tetanus toxoid were relatively fewer (28 or 34.1% in HIV patients and 8 or 66.7% in healthy controls) and weak compared to that to C. albicans (p <0.01). Among the HIV-1-infected individuals, the discordant patients demonstrated a significantly lower degree of lymphocyte proliferation (mean ± SD; SI = 17.6 ± 6.3) than concordant patients (mean ± SD; SI = 32.9 ± 3.9; p < 0.05), with no significant difference in the percentage of responders, upon stimulation with C. albicans. However, proliferation of lymphocytes of discordant patients was comparable to that of normal healthy controls in C. albicans–stimulated cultures, indicating no compromise in their proliferative capacity. In response to tetanus toxoid also, both discordant and concordant individuals had similar amounts of lymphocyte proliferation compared to normal healthy controls (Fig. 2).
Discordant patients have a higher relative percentage of CD4 TCM, CD8 TCM, and a lower percentage of naïve CD4 T cells in peripheral blood
In order to examine the differences in the phenotypes of T cells in the peripheral blood of recruited subjects, we characterized CD4 and CD8 T-cell subsets on the basis of surface expression of CD45RA and CCR7 (7,20). Compared to normal healthy controls, the HIV-1-infected patients had a significantly higher percentage of CD4 TCM cells (mean ± SD; 41.2 ± 10.0% versus 32.6 ± 4.2%) and CD8 TEM cells (mean ± SD; 30.3 ± 9.1% versus 20.1 ± 2.2%; p < 0.05). The relative percentage of naïve CD4 T cells was lower (mean ± SD; 32.4 ± 8.2% versus 41.9 ± 7.5%), and CD8 T cells expressing perforin were higher (mean ± SD; 20.8 ± 8.5% versus 10.7 ± 4.9%) in HIV-1-infected patients.
Among the HIV-1-infected patients, the discordant patients had a higher relative percentage of CD4 and CD8 T cells with the TCM phenotype than the concordant HIV-1-infected patients (CD4+CD45RA−CCR7+: 48 ± 7.6% versus 34.4 ± 7.3%; CD8+CD45RA−CCR7+: 23.3 ± 5.3% versus 15.8 ± 5.2%; p < 0.05) (Fig. 3A and B). Naïve CD4 T cells were significantly lower in discordant patients (mean ± SD; 26.6 ± 3.8%) than in concordant patients (mean ± SD; 38.8 ± 7.2%; p < 0.05). Both the groups had similar relative percentages of naïve and terminally differentiated CD8 TEMRA cells. The relative percentages of CD8 TEM cells were also similar in both the groups (mean ± SD; 31.4 ± 9.3% in concordant and 27.4 ± 4.5% in discordant patients).
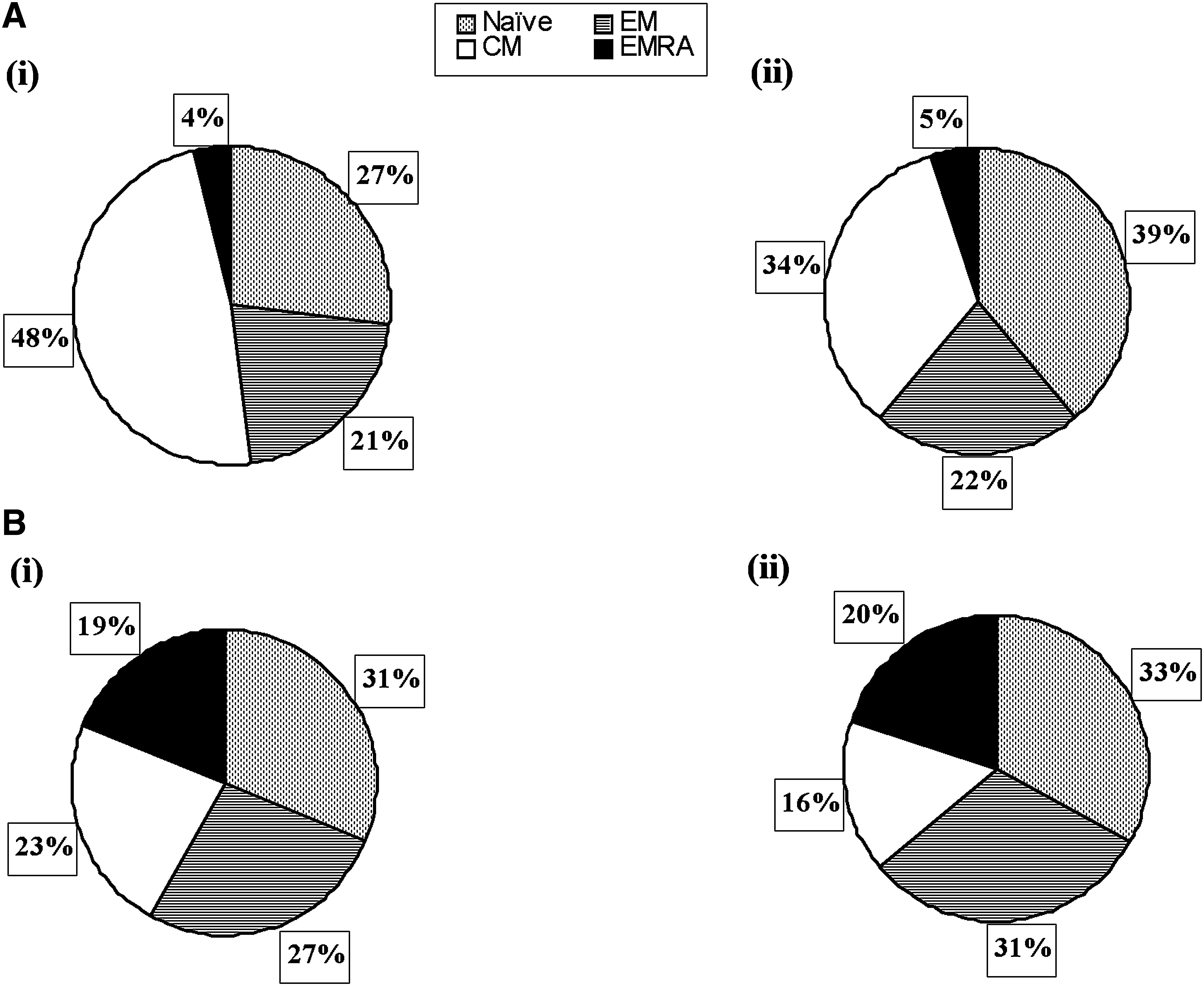
Distribution of (
Discordant patients show lower percentages of perforin- and granzyme-B–producing CD8 T cells in HIV-1 Gag p24–stimulated in-vitro cultures
Besides measuring proliferation of PBMCs after stimulating them with antigens, we analyzed the percentage of CD8 T cells for intracellular production of perforin and granzyme-B as markers of their function. Prior to stimulation, CD8 T cells expressing perforin were significantly higher in concordant patients (mean ± SD; 24.5 ± 8.1% versus 14.1 ± 5.4%; p < 0.05), while the expression of granzyme-B was relatively the same in both the groups (Fig. 4A). Following stimulation with complete Gag p24 antigen, CD8 T cells from concordant patients expressed a significantly higher percentage of perforin (mean ± SD; 30 ± 7.1% versus 14.9 ± 10.1%), and granzyme-B (mean ± SD; 51.1 ± 10.3% versus 32 ± 24.9%; p < 0.05) (Fig. 4B). In response to stimulation with peptide pool 2 of Gag p24, concordant subjects again demonstrated significantly higher perforin-producing CD8 T cells than did discordant subjects (mean ± SD; 32.5 ± 9.1% versus 17.8 ± 9.7%). Granzyme-B-producing cells were also higher in concordant subjects following in-vitro stimulation with peptide pool 2 of Gag p24, though the difference was statistically insignificant (mean ± SD; 52.2 ± 15.2% versus 34 ± 21.5%; p = 0.101) (Fig. 4C). However, in case of peptide pool 1 of Gag p24 and the recall antigens C. albicans (Fig. 4D) and tetanus toxoid, there were insignificant differences in numbers of CD8 T cells producing perforin and granzyme-B between the discordant and concordant HIV-1 infected patients.
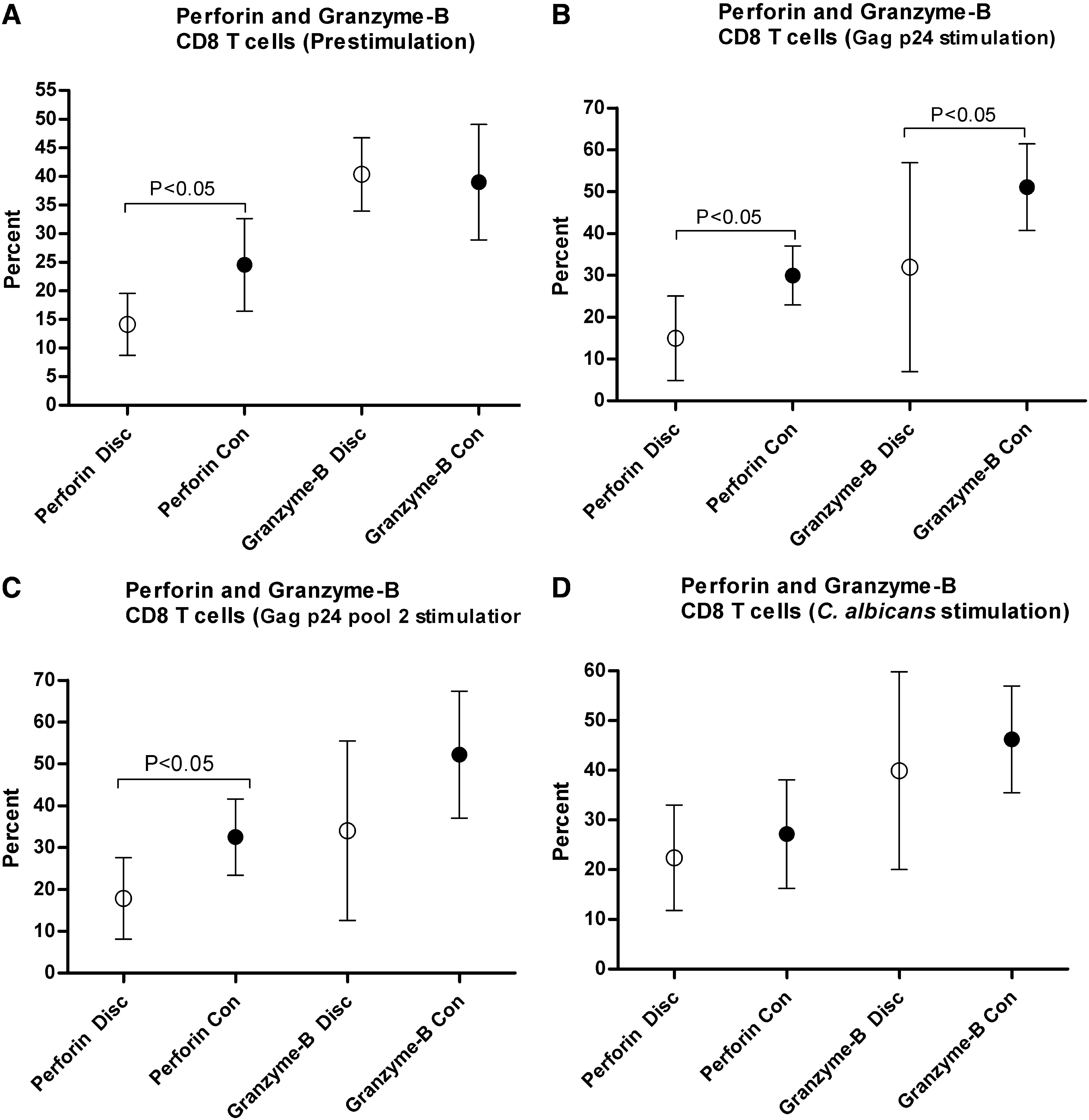
Relative percentage of perforin- and granzyme-B–producing CD8 T cells before and after 6-day stimulation with test antigens: Gag p24, peptide pool 2 of Gag p24, and C. albicans, in in-vitro cultures. (
Poor differentiation of terminally differentiated CD8 TEMRA cells in discordant patients following in-vitro stimulation is associated with a decline in CD4 TCM and CD8 TCM cells
Generation of effector CD8 T cells in response to an antigen is an indicator of an effective protective cellular immune response. Production of the effector molecules perforin and granzyme-B is mainly mediated by effector CD8 T cells differentiating from CD8 TEMRA cells. Such protective responses are also accompanied by an accumulation of CD4 TCM cells and CD8 TEM cells (21). After having observed the distribution of various CD4 and CD8 T-cell subsets prior to stimulation with antigens, we further investigated the relative proliferation of CD4 and CD8 T-cell subsets in response to various antigens.
Overall, the discordant patients showed a significant increase in the relative percentage of naïve CD4 T cells following stimulation with all antigens used in the study (p < 0.05) (Fig. 5A). The increase in naïve CD4 T cells was accompanied by a concomitant decline in the relative percentage of CD4 and CD8 T cells with the TCM phenotype (p < 0.05) (Fig. 5B and E). The decrease in the percentage of CD8 TCM cells was associated with an increase in the relative percentage of CD8 TEM cells (p < 0.05) (Fig. 5F). In contrast, the concordant patients demonstrated a marginal increase in CD4 TCM cells, and a significant increase in CD8 TEMRA cells in response to stimulation with the different antigens used in the study (p < 0.05). The variations in differential proliferation of CD8 T-cell susbsets between discordant and concordant patients following stimulation were more evident in the case of the Gag p24 antigen and peptide pool 2 of Gag p24, for which concordant patients showed a significantly higher percentage of CD8 TEMRA cells (mean ± SD; 41.3 ± 17.9% versus 17.3 ± 12.3%, and 47.2 ± 10.8% versus 24.8 ± 14.6%, respectively; p < 0.05) (Fig. 5G). The discordant patients did show an increase in CD8 TEM cells from baseline; however, the generation of terminally differentiated CD8 TEMRA cells was poor compared to that seen in the concordant patients. The discordant patients only showed a significant increase from pre-stimulation levels in CD8 TEMRA cells following stimulation with C. albicans (p < 0.05) (Fig. 5G). CD4 TCM cells were also higher in concordant subjects following stimulation with C. albicans, Gag p24, and peptide pool 2 of Gag p24, though the differences were statistically insignificant.
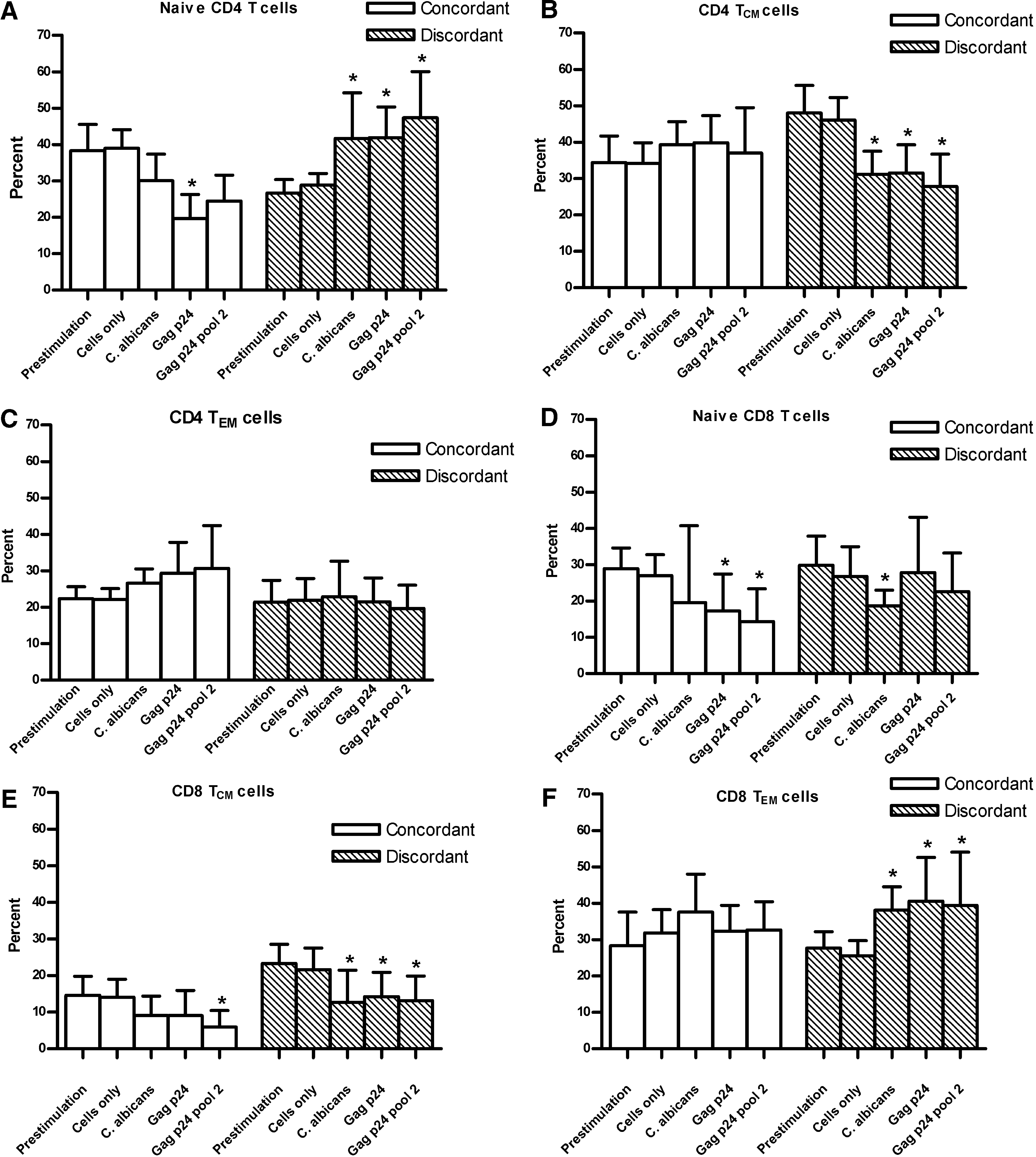
Changes in the relative percentage of CD4 and CD8 T-cell subsets in concordant and discordant HIV-1-infected patients following in-vitro stimulation with C. albicans, complete Gag p24 antigen, peptide pool 2 of Gag p24, and in the absence of exogenous stimuli (cells only). (

Discordant patients show defective production of IL-2 following stimulation with Gag p24
To further investigate the strength of cellular immune responses elicited against Gag p24 and C. albicans, we measured release of IL-2 and IFN-γ into the cell culture supernatants 24 h following stimulation of peripheral PBMCs. In general, PBMCs isolated from HIV-1-infected patients produced smaller amounts of IL-2 and IFN-γ following stimulation with PHA than did normal healthy controls, indicating inadequate CD4 T-cell help or persistence of HIV-specific defects in the infected patients. In contrast, in-vitro stimulation with Gag p24 induced higher IL-2 and IFN-γ production by HIV-1-infected patients (p < 0.05). Following stimulation with C. albicans, mean levels of IL-2 were lower in the discordant group than in the concordant group, though the differences were statistically insignificant. Importantly, the discordant patients produced smaller amounts of IL-2 following stimulation with Gag p24 (p < 0.05), reflective of their insufficient CD4 TCM and TEM numbers. However, there were no differences in the production of IFN-γ in both the discordant (mean ± SD; 17.1 ± 7.9 pg/mL), and the concordant groups (mean ± SD; 17.3 ± 9.0 pg/mL) following stimulation with Gag p24 (Fig. 6).
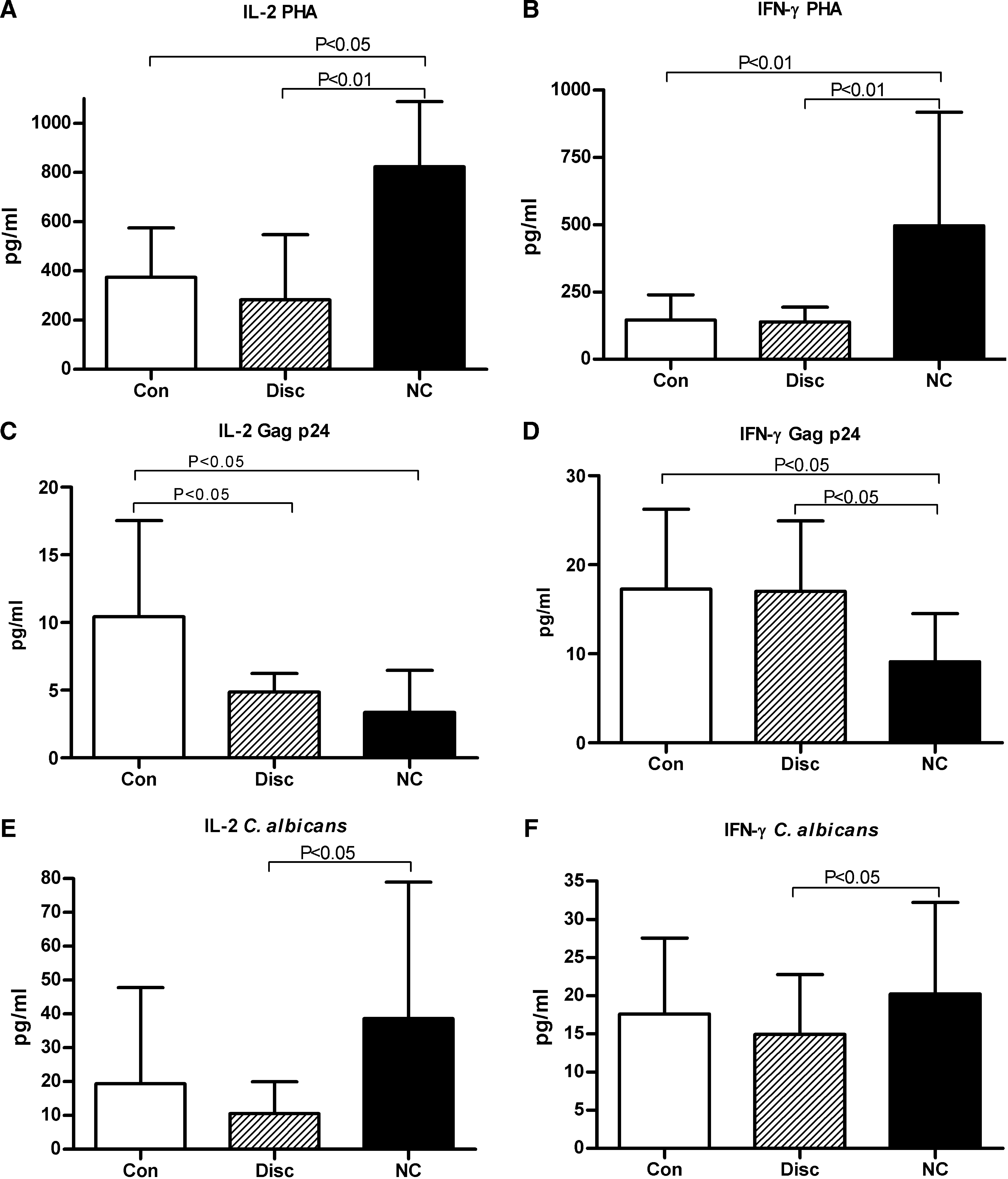
Production of IL-2 and IFN-γ from PBMCs 24 h following in-vitro stimulation with PHA (
Discussion
There is growing evidence that CD4 T cells are required to maintain the memory CD8 T-cell population in chronic HIV-1 infection (22,23). Prospective evaluation of T-cell responses in discordant patients provided an opportunity to assess the requirement of CD4 T cells in the generation of CD8 T-cell responses to HIV-1 and the antigens with regular or rare exposure. We selected a sufficient number of discordant and concordant patients in a controlled cohort and investigated functional memory CD8 T-cell responses using three approaches. At first we measured lymphocyte proliferation following stimulation with antigens; second, we measured the cytolytic potential of CD8 T cells in terms of perforin and granzyme-B production; and third, we measured the increase in the relative percentage of CD8 TEMRA cells in in-vitro cultures stimulated with Gag p24 and recall antigens. In addition, we quantitated production of IL-2 and IFN-γ in culture supernatants following antigenic stimulation. Robust Gag p24-specific CD8 T-cell responses have been shown to be associated with lowering of viremia, regardless of the HLA type (19). In our study the discordant patients showed poor proliferative responses and fewer perforin- and granzyme-B-producing CD8 T cells in in-vitro cultures stimulated with Gag p24 and one of its peptide pools, despite suppressed plasma viremia for long periods of time. Additionally, the discordant patients also showed defects in differentiation of CD8 T cells into effector phenotype following stimulation with the same antigens. The observed differences in discordant patients do not seem to be associated with distinct characteristics of virological response to HAART and drug resistance, since plasma viral loads were undetectable.
The proliferative responses of discordant patients following stimulation with recall antigens (especially C. albicans) were lower than those seen in concordant patients, but comparable to normal controls. However, the differences in perforin- and granzyme-B-producing CD8 T cells and generation of effector CD8 T cells between discordant and concordant patients were insignificant following stimulation. Additionally, the discordant patients did not have any active opportunistic infections at the time of analysis. Taken together, these observations indicate that protective CD8 T-cell responses to recall antigens were not perturbed to a large extent in the discordant patients. Earlier reports have shown regeneration of responses to recall antigens, but a lack of HIV-1-specific responses in patients treated with HAART (24). Similarly positive lymphoproliferative responses to C. albicans were reported in a small number of patients, similarly to our discordant group, with higher CD4 T-cell counts compared to HAART non-responders (25). Though such responses were seen in patients showing positive responses to HAART, our results demonstrate that protective CD8 T-cell immunity to recall antigens in discordant patients was largely unaffected, despite low numbers of circulating CD4 T cells. Thus it appears that discordant patients may not be vulnerable to opportunistic infections, unlike patients failing therapy, until their CD4 T-cell counts drop below a critical level. Furthermore, it confirms that restoration of protective CD8 T-cell responses following HAART is pathogen-specific.
Besides measuring overall lymphocyte proliferation and CD8 T-cell function, we observed differences in the proliferation of CD4 and CD8 T-cell subsets following stimulation with different antigens. Interestingly, the discordant patients had higher relative percentages of peripheral CD4 TCM and CD8 TCM cells than the concordant patients. Though these individuals had low-to-undetectable viral loads, some levels of underlying viral replication in the lymphoid compartment is sufficient to provide antigenic exposure, if required, for maintenance of memory T cells (26). However, after stimulation with Gag p24 and recall antigens, there was decline in the relative percentage of CD4 TCM cells, indicating that most of these cells were non-protective in nature. HIV-1 controllers usually preserve their CD4 TCM function in the absence of viral load, which appears to have been lost in the discordant patients (27 –29). The rate of memory CD4 or CD8 T-cell differentiation from TEM into the self-renewing and long-lived pool of TCM is influenced by signals received during priming, including antigen levels, clonal competition, and/or the duration of infection (21,30). Studies have demonstrated that CD8 TCM cells also mediate effector function and are more effective in conferring protective immunity (31,32). Though it is difficult to obtain information on the generation of TCM subsets during the initial stages of infection in human subjects, we have observed that despite the absence of other active infection and suppression of viremia, the discordant patients showed poor proliferative responses and lower IL-2 secretion in the presence of Gag p24. Thus poor restoration of CD4 T cells can also affect cytokine-dependent CD8 T-cell-mediated immune responses to certain antigens, such as HIV-1 Gag p24, as seen in this study.
Whether effector CD8 T cells are generated from CD8 TEM cells alone, or both lineages are generated independently, is still unclear. However, it is well known that TEMRA cells have the ability to reactivate effector functions besides undergoing vigorous proliferation upon subsequent challenge with the same antigen (33). The percentage of CD8 TEM cells in the discordant patients was comparable to that seen in the concordant patients. However, we observed generation of functionally impaired CD8 T cells upon in-vitro stimulation, especially in case of Gag p24, which again reflects upon the incompetent state of CD8 TEM cells. One of the hallmarks of controlled HIV-1 infection is the production of fully differentiated HIV-1-specific effector CD8 T cells (34 –37). Despite maintenance of low viral load, the discordant patients demonstrated characteristics of a progressive HIV-1 infection in terms of differentiation of CD8 TEMRA cells. It seems likely that the persistence of low CD4 T-cell numbers affected the differentiation of CD8 TEM cells, resulting in insufficient and dysfunctional HIV-1-specific CD8 TEMRA cells.
The discordant patients were depleted of naïve CD4 T cells despite HAART-mediated viral suppression. This condition generally arises either due to failure in CD4 T-cell reconstitution, or due to HIV-1 replication-mediated destruction of CD4 T cells, making HIV-1-reactive CD8 T cells non-functional (38 –40). There was no active viral replication in the discordant patients, as indicated by viral load and expression of CD38, CD69, and HLA-DR on CD4 T cells (17). Prior to initiation of HAART, the discordant patients indeed had significantly higher baseline viral load and low CD4 T-cell counts. Higher baseline plasma viral loads and low CD4 T cell counts have been reported to predict loss of Gag-specific responses in chronic HIV-1 infection (41). In addition, we have earlier reported that there is loss in number and function of plasmacytoid dendritic cells in discordant patients, reflective of the immune damage inflicted by high levels of HIV-1 replication in these individuals in the past (42). Reconstitution of peripheral CD4 T cells is biphasic, involving a rapid increase in memory CD4 T cells, followed by a slow increase in naïve CD4 T cells (43). The presence of higher percentages of CD4 TCM and CD8 TCM cells in the discordant patients indicates that most of the discordant patients might be in a pre-naïve CD4 T-cell expansion phase. Thus the impairments in the memory T-cell responses observed in the discordant patients may be attributable to poor CD4 T-cell reconstitution. However, there are various factors associated with the dynamics of T-cell reconstitution that must be considered before drawing any conclusions.
Conclusion
To conclude, whether the mechanisms that underlie immunological and virological discordance are merely a consequence of greater immune damage before initiation of antiretroviral therapy, or whether they represent a different potential for immune reconstitution remains unclear. In this study we have shown that while functional CD8 T-cell responses to recall antigens are maintained in discordant patients, the intensity of HIV-1-specific responses is diminished, largely due to inadequate CD4 T-cell help despite maintenance of undetectable viral loads and high CD8 T-cell numbers. With increasing access to antiretroviral therapy worldwide, discordant responders to HAART are expected to increase further in numbers. The information obtained from this study might be useful for understanding T-cell differentiation, and aid in the design of immunotherapeutic approaches against HIV-1 and opportunistic pathogens in these patients.
Footnotes
Acknowledgments
This work was supported by the UM-LCBS educational and training funds. The HIV-1 Gag p24 peptides were provided by the EU Programme EVA Centre for AIDS Reagents, NIBSC, U.K. (AVIP contract number LSHP-CT-2004-503487). The authors thank Prof. Michael Lederman, Director, Center for AIDS Research, Case Western Reserve University, for critical evaluation of the manuscript and valuable suggestions.
Author Disclosure Statement
No conflicting financial interests exist.