
Abstract
Chikungunya virus infection has recently emerged in several countries. The inflammatory response is suggested to be involved in the pathology observed in this infectious disease. Interleukin-18 (IL-18) is an inducer of interferon-γ (IFN-γ) production and has been shown to play a role in several inflammatory diseases. IL-18 binding protein (IL-18BP) is a natural regulator of IL-18. In this study, we determined the levels of IL-18 and IL-18BP in patients with Chikungunya virus infection. Acute and convalescent sera were collected from each patient. The levels of both IL-18 and IL-18BP were measured by ELISA assays. IL-18 and IL-18BP levels were higher in patients than in controls. In addition, the level of IL-18 was higher in convalescent than in acute sera. However, the level of IL-18BP was lower in convalescent than in acute sera. These data suggest that production of both IL-18 and IL-18BP was induced following Chikungunya virus infection. IL-18BP was increased to regulate the activity of IL-18. The ratio of IL-18 to IL-18BP was higher in convalescent than in acute sera. The lower level of IL-18BP in convalescent sera was probably due to loss following IL-18 neutralization. Our data suggest that Chikungunya virus infection promotes the T helper-1 (Th-1) response by inducing IL-18 production. Manipulation of IL-18 and IL-18BP levels could be a promising therapeutic approach to alleviate symptoms in patients with Chikungunya virus infection.
Introduction
Interleukin 18 (IL-18) is a member of the IL-1 family, and it was originally known as an interferon-γ (IFN-γ)-inducing cytokine. IFN-γ enhances macrophage, NK cell, and cytotoxic T-cell activity. For these reasons, IL-18 has been studied as a cytokine involved in host defense, especially in the killing of intracellular organisms and eradication of cancer cells. The predominant cellular sources of IL-18 are macrophages and dendritic cells. However, IL-18 can be produced by other cell types, such as Kupffer cells, T cells, B cells, osteoblasts, keratinocytes, and astrocytes (6).
IL-18 binding protein (IL-18BP) is a natural IL-18 antagonist. It binds IL-18 in the fluid phase (7). Binding of IL-18BP to IL-18 prevents IL-18 from binding to IL-18Rα on the target cell membrane, and thus interferes with high-affinity IL-18R formation. Functional homologues of human IL-18BP encoded by viruses such as orthopoxviruses have been shown to inhibit IL-18 activity (8). There are four isotypes (IL-18BPa, b, c, and d) of human IL-18BP generated from alternative mRNA splicing. Only IL-18BPa and IL-18BPc have intact Ig-like domains and can neutralize IL-18. The role of the other two isoforms remains unknown (9).
It has been shown that IL-18 is one of the cytokines involved in the pathogenesis of inflammatory disease (9,10,11). The IL-18 level was increased in patients with dengue virus infection, and its level correlated with disease severity (12). A role of IL-18 and its natural inhibitor IL-18BP in the pathological process of rheumatoid arthritis has been proposed. An association of IL-18 with the response to treatment of patients with rheumatoid arthritis has also been shown (13). Serum levels of IL-18 were increased in rheumatoid arthritis patients, whereas the level of IL-18BP was decreased (14). Administration of a vector containing IL-18BP and IL-4 genes reduced Th-1 cytokines and IFN-γ-producing T cells in mice with collagen-induced arthritis. These data suggest that besides enhancing Th-2 cytokine production, regulation of IL-18 by its natural ligand could be another promising tool for the development of therapies for rheumatoid arthritis (15).
In this study, we determined the levels of both IL-18 and IL-18BP in sera of Chikungunya patients. The levels were determined in both acute and convalescent sera to investigate whether the amounts of IL-18 and its natural regulator changed during infection. The data obtained will provide further information on the host immune response, which may be involved in the pathologies induced by this infectious disease.
Materials and Methods
The study protocol was approved by the Ethical Committee, Faculty of Medicine, Chulalongkorn University. All specimens were anonymous specimens used for this cytokine study with the permission of the director of Chulalongkorn Hospital.
Specimen collection
Sera were obtained from patients who presented to Narathiwatratchanakharin Hospital, Narathiwat, Thailand, with acute febrile illness and sent to the Center of Excellence in Clinical Virology, Chulalongkorn Hospital, for diagnostic purposes. The remaining sera were kept at −70°C until analysis. Baseline clinical data including the history of the present illness, physical examination findings, and complete blood count, were collected. Sera from patients with suspected Chikungunya virus infection were confirmed by ELISA for anti-Chikungunya IgM, and semi-nested RT-PCR for Chikungunya virus RNA, as previously reported (16).
Paired sera from 32 female and 14 male patients, aged 2–84 y old (mean 39.9 ± 19.8 y), in whom Chikungunya infection was confirmed by IgM ELISA and semi-nested PCR, were investigated in this study. Acute sera was collected 2–6 d after the onset of fever, and convalescent sera was obtained 5–13 d after retrieval of the acute sera.
Sera from 20 donors at the National Blood Center, Thai red Cross Society, Thailand, Bangkok, were used as controls. All donor sera were screened and no infectious agents were detected.
Determination of IL-18 and IL-18BP levels by captured ELISA
Reagents for captured ELISA for IL-18 and IL-18BP were purchased from MBL (Nagoya, Japan) and R&D systems (Minneapolis, MN). The assays were performed according to the manufacturer's instructions. Briefly, for IL-18 determination, sera and standard solution were added to anti-IL-18-coated wells. IL-18 was then captured by anti-IL-18 conjugated with peroxidase. The presence of IL-18 was visualized by adding TMB substrate. The absorbance at 450/620 nm was read by a dual wavelength plate reader. A standard curve was constructed and the concentration of IL-18 in the samples was read from the standard curve.
For IL-18BP, IL-18BPa was determined, as it represents an isoform with widely known IL-18-neutralizing activity. The microtiter plate was coated with anti-IL-18BPa followed by 1% BSA blocking. Sera and standard solution were then added. IL-18BPa was captured by adding biotinylated anti-IL-18BPa. The presence of IL-18BPa was visualized by adding streptavidin-horseradish peroxidase followed by TMB substrate. Absorbance at 450/570 nm was read, and the standard curve was constructed according to the manufacturer's recommendation. The IL-18BPa level was read from the standard curve.
Statistical analysis
The difference between IL-18 or IL-18BP levels in acute and convalescent sera was determined by paired t-test. Unpaired t-test was used to demonstrate the difference between the levels in the control and patient groups. p-values <0.05 were considered statistically significant.
Results
The levels of IL-18 in 20 control sera ranged from 19.6–135.8 pg/mL. The levels in acute and convalescent sera from Chikungunya patients ranged from 78.8–1160.3 and from 79.4–1806.6 pg/mL, respectively. The levels of IL-18 in patient sera were statistically significantly higher than the levels in the sera from the control group (p < 0.001). In addition, IL-18 levels in convalescent sera were higher than those in acute sera (p = 0.03). This suggested an increase in IL-18 levels following infection.
The levels of IL-18BP in the control group ranged from 0.26–0.69 ng/mL. The levels in acute and convalescent sera from the patient group ranged from 1.14–8.09 and 0.82–7.3, respectively. IL-18 BP levels were also significantly higher in Chikungunya patients than in controls (p < 0.001). However, the levels of IL-18BP in convalescent sera were significantly lower than those in acute sera (p < 0.001). The ratio of IL-18 to IL-18BP was significantly higher in convalescent sera than in acute sera (p < 0.001). These data suggest that, although the levels of IL-18BP were increased compared to controls, they were later reduced over the course of the infection.
The results are shown and summarized in Fig. 1 and Table 1.
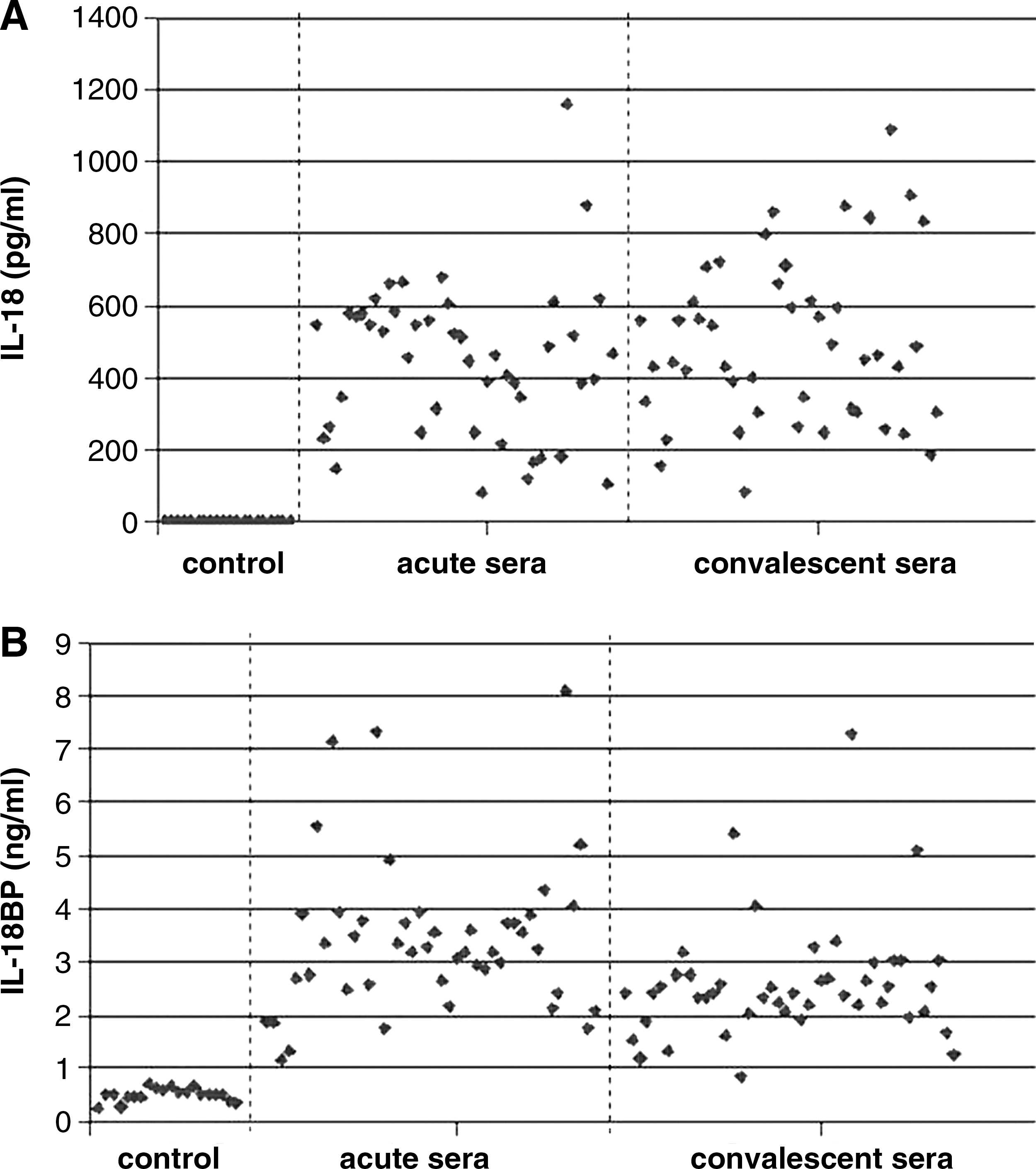
Comparison of IL-18 (
Discussion
Although Chikungunya was discovered more than 50 y ago, there are only a few reports on the mechanism underlying its pathogenesis or the immune response induced by this infectious agent. It has been suggested that the inflammatory response could be the cause of persistent arthralgia in Chikungunya patients. Increased IL-1β and IL-6 levels have been shown to correlate with higher disease severity (5). Both cytokines are involved in the inflammatory response.
IL-18 is a cytokine of the same family as IL-1β, and it promotes IFN-γ production. IFN-γ enhances the Th-1 response, which has been shown to participate not only in immunity, but also in disease pathology caused by chronic inflammatory responses (17 –20). According to our data, both IL-18 and IL-18BP levels were increased in Chikungunya patients compared with controls. This suggests that Chikungunya infection induces production of IL-18 and IL-18BP. Ng et al. (5) demonstrated that IL-1β and IL-6 levels are related to disease severity. Several proinflammatory cytokines may work together to enhance disease severity in Chikungunya infection. We analyzed data from Ng et al. along with our own, and thus hypothesize the involvement of IL-1β, IL-6, IL-18, and IL-18BP in Chikungunya infection, as shown in Fig. 2. IL-1β and IL-6 are known to be proinflammatory cytokines, and induction of these two cytokines induces the inflammatory response. The increased IL-18 levels seen subsequent to Chikungunya virus infection could lead to IFN-γ production, which may enhance the effect of IL-1β and IL-6, and thus the inflammatory reaction. IL-18BP is a natural regulator of IL-18 and is induced by IFN-γ. This suggests that IFN-γ provides negative feedback for IL-18 suppression by inducing IL-18BP production. The inhibition of IL-18 activity by IL-18BP could be one of the mechanisms that regulates inflammation. An imbalance between IL-18 and IL-18BP production could thus be the underlying cause of inflammatory disease.
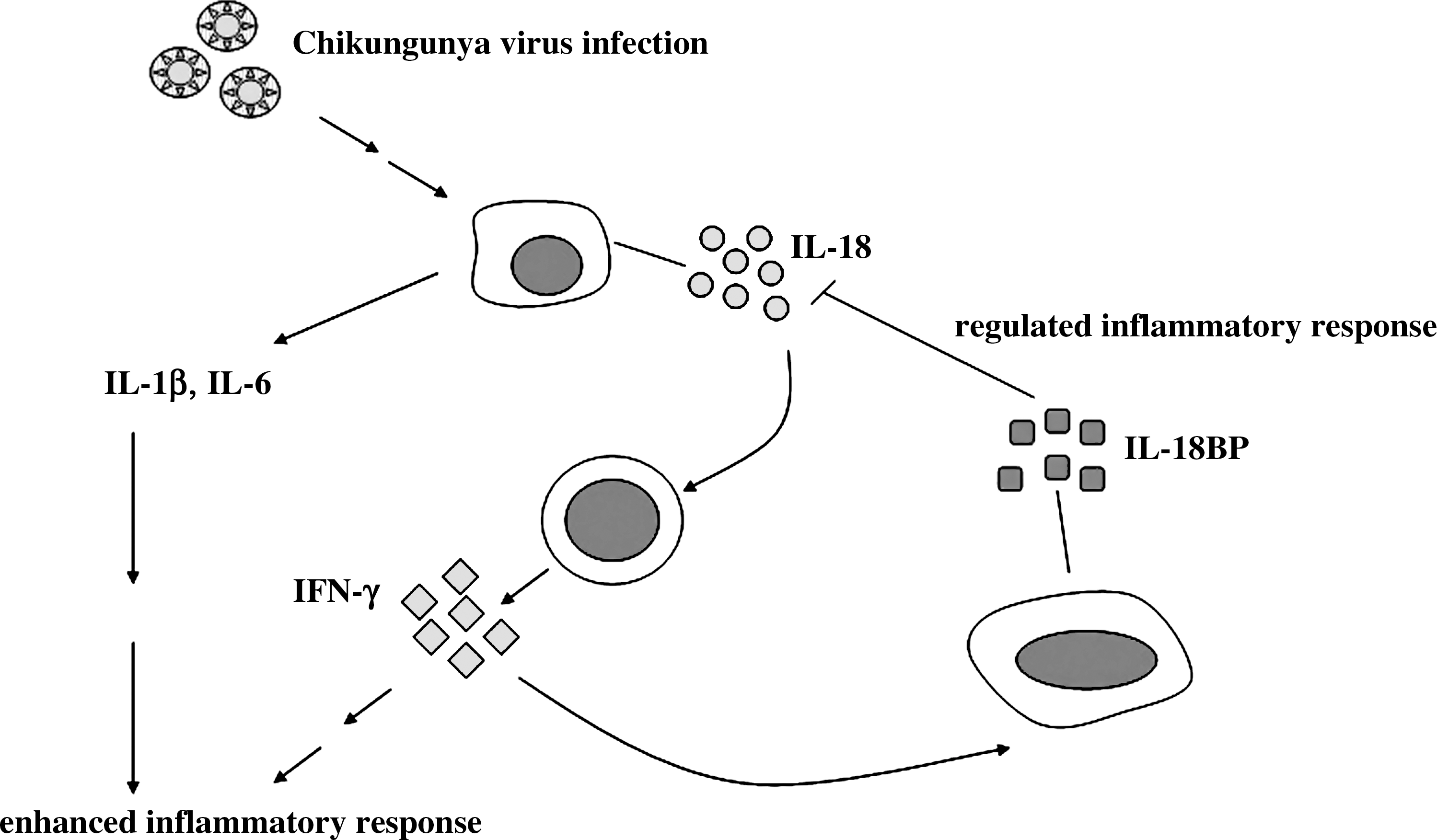
The roles of IL-1β, IL-6, IL-18, and IL-18BP in enhancing and regulating the inflammatory response. Chikungunya virus infection induces IL-1β, IL-6, and IL-18 production, and IL-1β and IL-6 induce the inflammatory response. IL-18 induces production of IFN-γ, which further enhances the inflammatory reaction. IFN-γ negatively regulates IL-18 by inducing IL-18BP production. Neutralization of IL-18 activity by IL-18BP regulates the inflammatory response.
While the levels of IL-18 in convalescent sera were higher than those in acute sera, the levels of IL-18BP were lower in convalescent sera than in acute sera. The lower level of IL-18BP in convalescent sera was probably due to a loss in the process of IL-18 neutralization. Although IL-18BP production was enhanced compared to controls, its amount was still insufficient to reduce the level of IL-18 over the course of infection. While IL-18BP was reduced, the level of IL-18 was constant. The ratio of IL-18 to IL-18BP was decreased in convalescent sera, due to the significant reduction in IL-18BP, but not IL-18. The proposed mechanism behind the lower level of IL-18BP is shown in Fig. 3. An imbalance of IL-18 and IL-18BP has been shown in patients with lupus nephritis. An elevated ratio of IL-18 to IL-18BP was also shown in lupus nephritis, but not in primary nephritic syndrome, compared to normal controls (21).
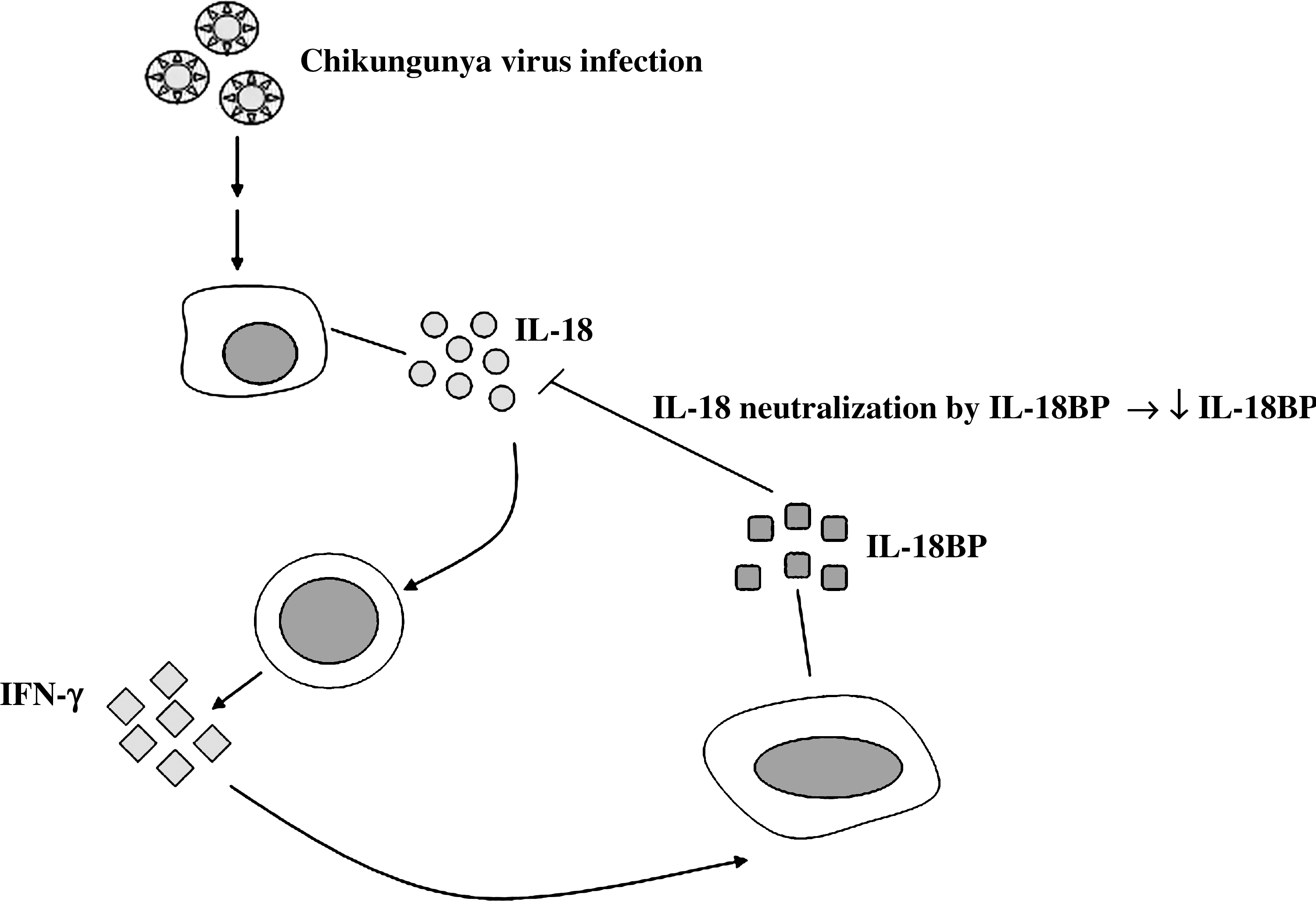
Decreased IL-18BP level via IL-18 neutralization. Both IL-18 and IL-18BP were induced during the acute stage of infection. IL-18BP neutralizes the activity of IL-18, resulting in the decrease in the level of IL-18BP seen in the convalescent samples.
Conclusion
Further investigation of the mechanisms behind the regulation of IL-18 and IL-18BP and their balance could help elucidate the molecular mechanisms at work in the pathology induced by Chikungunya virus infection, and aid in the development of therapies to treat this infection.
Footnotes
Acknowledgments
This study was supported by the Higher Commission on Education, Ministry of Education, Rachadapisek Sompoch Research Fund, Faculty of Medicine, Chulalongkorn University, King Chulalongkorn Memorial Hospital, the Center of Excellence in Clinical Virology, and the CU Centenary Academic Development Project. We also would like to thank all of the staff members of the Narathiwatratchanakharin Hospital who participated in this project, and Ms. Petra Hirsch for reviewing the manuscript.
Author Disclosure Statement
No conflicting financial interests exist.