
Abstract
The polyomaviruses WUPyV and KIPyV were recently discovered. We expressed their structural proteins VP1, VP2, and VP3, and the corresponding proteins of BKV and JCV, for immunoblotting of IgG antibodies from 115 wheezing young children and 25 asymptomatic adults. Furthermore, nasopharyngeal aspirates (NPA) and sera from the children were examined by PCR for viral DNA. The overlapping minor proteins VP2 and VP3 of WUPyV and KIPyV were more reactive in immunoblots than the major protein VP1; of 100 NPA PCR-negative wheezing children aged ≤4 y, 31 (31%) and 31 (31%) were positive for WUPyV and KIPyV VP2/VP3, compared to only 3 (3%) and 5 (5%) for VP1, respectively. For comparison, the respective WUPyV and KIPyV IgG seroprevalences as determined by immunofluorescence assay (IFA) with nondenatured VP1 were 80% and 54%, respectively, among 50 NPA PCR-negative children aged ≤2 y. This difference shows the importance of conformational VP1 antigenicity. Of the 25 adults, 52% and 68% were IgG-positive in immunoblots for VP2/VP3 of WUPyV and KIPyV, and 8% and 12% were for VP1, respectively. Of the 192 NPA samples studied by PCR, 7 (3.6%) were positive for WUPyV, and 3 (1.5%) were positive for KIPyV DNA. Unlike the NPA samples, none of the corresponding 443 sera contained WUPyV or KIPyV DNA. Together with the high VP2/VP3 IgG prevalence, this points to a paucity or brevity of KIPyV and WUPyV viremias among immunocompetent children. Our results indicate the significance of protein conformation in immunoreactivity of VP1, and show the antigenic importance of the WUPyV and KIPyV minor proteins VP2 and VP3. The high and rapidly increasing IgG prevalence rates observed in this study for WUPyV and KIPyV support the notion that these novel polyomaviruses are widespread and are acquired early in childhood.
Introduction
Two new members of the Polyomaviridae family, provisionally named KI polyomavirus (KIPyV) and WU polyomavirus (WUPyV), were recently discovered in Sweden and in the U.S. by screening of human respiratory samples with large-scale sequencing and bioinformatics (3,11). In genome organization KIPyV and WUPyV are typical polyomaviruses, yet in amino acid sequence their structural proteins are only 15–29% identical to those of BKV and JCV. The “late” regions of KIPyV and WUPyV genomes show 64–71% amino acid sequence identity.
KIPyV and WUPyV, like other polyomaviruses, encode three structural proteins, VP1, VP2, and VP3, and the early region proteins large and small T-antigen. VP3 is an N-terminal truncation of VP2, due to different translation initiation codons and RNA splicing (6). The polyomavirus capsid consists of VP1 pentamers with a single VP2 or VP3 molecule in the center of each pentamer unit. Two recent studies have shown that, in its native form, VP1 of KIPyV and WUPyV is highly immunoreactive; antibody prevalence for the major capsid protein among immunocompetent individuals was found to be 55–70% for KIPyV and 69–80% for WUPyV (16,24). The etiological roles of KIPyV and WUPyV in respiratory disease have yet to be clarified (25).
The aims of this study were: (i) to express and purify the structural proteins of KIPyV, WUPyV, JCV, and BKV; (ii) to compare the immunoreactivities in immunoblotting; and (iii) to study the antibody and DNA prevalences of the novel polyomaviruses in young children with respiratory disease.
Materials and Methods
Patients and samples
The study was comprised of subjects of three groups. The first group consisted of 278 children aged 3 mo to 15 y (mean 2.3 y, median 1.6 y), admitted to the Department of Pediatrics of Turku University Hospital (Turku, Finland) from September 1, 2000 through May 31, 2002 for acute wheezing (14). A total of 443 serum samples (201 paired, 41 unpaired) from 242 individuals, and 192 nasopharyngeal samples from 192 patients, were studied for the presence of KIPyV and WUPyV DNA by polymerase chain reaction (PCR). The nasopharyngeal aspirates (NPA) were obtained on admission, while the sera were collected on admission and 2–3 wk after discharge. From the sera tested by PCR, 120 samples (100 unpaired, 10 paired) were selected on the basis of age (≤4 y), or nasopharyngeal aspirate (NPA) PCR result (KIPyV or WUPyV DNA-positive) for immunoblotting, and 50 (≤2 y) for immunofluorescence assays (IFA). These children had been studied thoroughly for the presence of respiratory viruses other than KIPyV and WUPyV (4).
The second group consisted of five children aged 0.7–5.6 y (mean 3.0 y, median 2.8 y), hospitalized in Germany and shown to be WUPyV PCR-positive in NPA (23). The sera from these children were taken on the same day (n = 3), or 4 d or 5 mo after the first NPA samples. Hereafter, all children with KIPyV or WUPyV DNA in NPA are collectively designated NPA PCR positive.
The third group consisted of 25 asymptomatic adults aged 20.5–60.7 y (mean 33.5 y, median 27.8 y), mostly laboratory staff, from whom only serum samples were obtained. These adult sera were immunoblotted without PCR testing.
Recombinant protein production
The VP1, VP2, and VP3 genes of the four human polyomaviruses were amplified by PCR using Platinum Taq DNA polymerase (Invitrogen Corp., Carlsbad, CA) from partial or full-length KIPyV (3), WUPyV (11), BKV, and JCV (20) clones. All 12 amplified genes were cloned into the pET SUMO vector (Invitrogen) for protein expression in E. coli. Protein expression utilized an N-terminal fusion tag, a Saccharomyces cerevisiae Smt3 protein (hereafter referred to as SUMO), an 11-kDa homolog of the mammalian SUMO-1. The tertiary structure of this tag is very accurately recognized by a SUMO protease, which cleaves it such that no amino acids remain between the cleavage site and the protein of interest (21). The SUMO expression constructs were transformed into E. coli BL21 (DE3) pLysS (Novagen, Gibbstown, NJ), or Rosetta 2 (DE3) pLysS (Novagen) expression strains, and grown overnight in 500 mL Luria broth (LB) with 50 μg mL–1 kanamycin (BL21 and Rosetta) and 34 μg mL–1 chloramphenicol (Rosetta). At an optical density of 0.6 at 600 nm, protein expression was induced with 1 mM IPTG for 3 h. Inclusion bodies from lysed cells were pelleted by centrifugation, washed twice with phosphate-buffered saline (PBS), and resuspended in 25 mL of lysis buffer (6 M GuHCl, 100 mM NaH2PO4, and 10 mM Tris · Cl, pH 8.0). The rest of the purification was done according to Ni-NTA matrix manufacturer (Qiagen Inc., Valencia, CA) instructions.
The purified proteins were renatured at 4°C by a 4-h dialysis against 2 M (WUPyV and KIPyV) or 3 M urea (BKV and JCV) containing 150 mM NaCl, 5% (v/v) glycerol, and 1 mM DTT in 20 mM Tris/HCl, pH 8.0. Cleavage of the SUMO carriers from the partially unstable WUPyV, BKV, and JCV VP3 proteins was done with 1 U of SUMO protease (Invitrogen) per 2 μg of substrate at 30°C for 2 h. All other proteins were cleaved overnight at 4°C with 1 U of protease per 10 μg substrate. The cleaved WUPyV and KIPyV proteins were passed twice through a Ni-NTA spin column (Ni-NTA spin kit; Qiagen) to remove the hexahistidine-tagged SUMO protein and the SUMO protease.
The WUPyV VP1, VP2, and VP3 proteins were expressed native without a fusion tag in Sf9 insect cells. The genes were cloned in the pCR8/GW/TOPO gateway entry vector and transferred to pDest8 gateway destination vector to create recombinant baculoviruses as previously described (2; Table 1).
Western blotting and immunofluorescence assay
The purifed proteins VP1, VP2, and VP3 were separated by 12% SDS-PAGE and immunoblotted with human sera and horseradish peroxidase-conjugated anti-human IgG. NPA PCR-positive children were also tested for IgM as previously described (15). In immunoblotting, a sample was regarded as seropositive if it had an unequivocal band of correct molecular size, and all samples were scored in blinded fashion. An increase in IgG was defined as a significant elevation in band intensity between the acute and convalescence serum samples, when both were tested identically and simultaneously.
Borderline sera were retested with a lower dilution of serum (1:30). Samples that were still borderline were regarded as negative. The IgG seroprevalences against the SUMO fusion tag, seen with the isolated carrier protein, increased with age from 8% among children to 72% in adults. To avoid false-positivity due to the SUMO fusion partner, both cleaved and uncleaved recombinant proteins were used; those subjects (n = 30) that were reactive with all three uncleaved antigens were re-examined with the cleaved antigens. To account for the possibility that the prokaryotic origin would affect the antigenicity of the recombinant proteins, all sera from 3- to 4-year-old children (n = 50) were also immunoblotted with WUPyV VP1- and VP3-expressing insect cells.
To validate our immunoblot (IB) results and further study the importance of protein folding for the antigenicity of KIPyV and WUPyV VP1, an immunofluorescence assay (IFA) using baculovirus-infected SF9 insect cells (Neske F, Prifert C, Ewald M, and Weissbrich B, unpublished results) was used to test the sera of 54 children for WUPyV and KIPyV VP1 IgG. The samples for the IFA assay were selected on the basis of age (≤2 y, n = 50).
Briefly, SF9 cells infected with recombinant baculoviruses were collected by centrifugation, washed with PBS, mounted on glass slides, and fixed with acetone. Slides with uninfected SF9 cells and SF9 cells infected with a control baculovirus encoding glutathione S-transferase (GST) were prepared in the same manner to control for anti-cellular and anti-baculoviral antibodies. The slides were incubated with 1:10 and 1:40 replica dilutions of each serum for 2 h at 37°C, washed, and incubated with a 1:150 dilution of fluorescein isothiocyanate (FITC)-conjugated goat anti-human-IgG (Jackson ImmunoResearch West Grove, PA) for 1 h at 37°C. The slides were read in blinded fashion.
WUPyV and KIPyV PCR
Nucleic acid extraction from the NPA was performed in a previous study (4). The extracts underwent single-tube, dual-probe, real-time PCR assay for KIPyV and WUPyV DNA as previously described (19). From the sera (50 μL each), DNA was isolated with the QIAamp DNA Blood Mini Kit. PCR with primer set A was performed as described elsewhere (25). The sensitivities of the PCRs were assessed with serial dilutions of plasmids containing the KIPyV and WUPyV VP2 inserts (5 × 103 to 5 × 10–1 copies per reaction).
Results
PCR
Altogether 443 (201 paired, 41 unpaired) sera of 242 young children with respiratory symptoms were studied by PCR for KIPyV and WUPyV DNA. All the sera were negative. Both of the viral target sequences in single copies were readily detectable in control preparations. By contrast, among the 192 NPA samples studied, 3 (1.5%) were positive for KIPyV and 7 (3.6%) were positive for WUPyV DNA. Of the 7 and 3 children positive for WUPyV and KIPyV, 4 and 1, respectively, had a high load of DNA (≥104 copies/mL of the original sample); the DNA loads in the remaining samples were ≤500 copies/mL.
Expression and purification of polyomavirus structural proteins
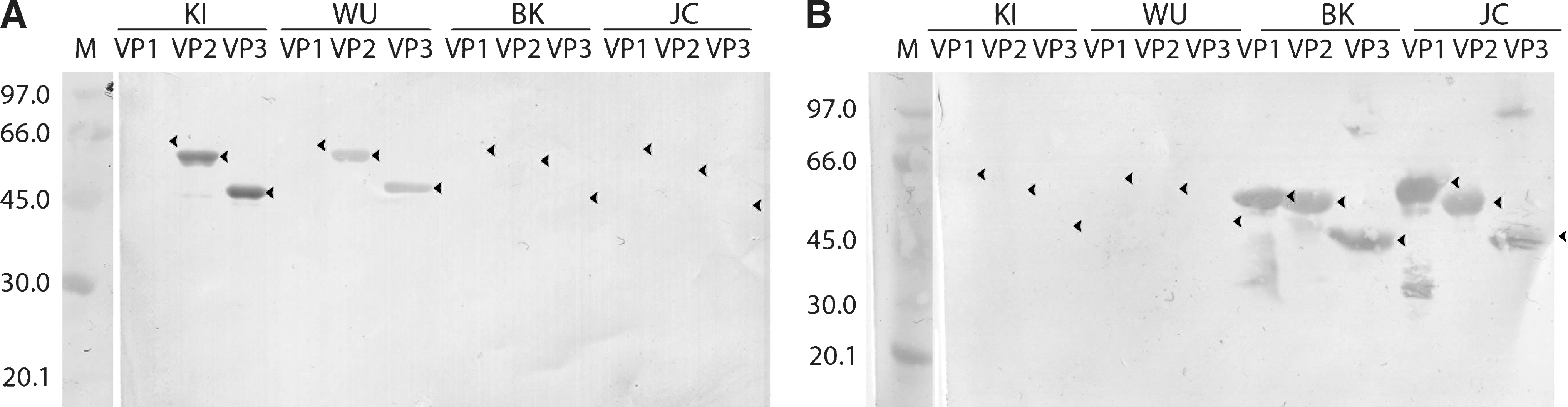
The structural proteins VP1, VP2, and VP3, of KIPyV, WUPyV, BKV, and JCV, each were expressed with the SUMO expression system in two E. coli strains. The expression levels in the Rosetta strain were in general higher, most notably for WUPyV VP1, which was expressed at levels 3–4 times higher than with the BL21 (DE3) strain (Fig. 1b). Protein yield was high in all cases, with 1 L of culture resulting in 20–25 mg of purified protein. Due to low solubility, all proteins were purified under denaturing conditions to >95% purity as estimated by SDS-PAGE (Fig. 1). Expression of WUPyV VP1 and VP3 in SF9 cells resulted in major bands in SDS-PAGE (Fig. 2), whereas VP2 was undetectable (data not shown).

SDS-PAGE of prokaryotically expressed, purified, and cleaved KIPyV (KI;

SDS-PAGE of Sf-9 insect cells infected with baculoviruses encoding WUPyV VP1 and VP3 (M, molecular weight; N, noninfected SF9 cells). Arrowheads indicate the observed positions of the recombinant proteins.
SUMO protease cleavage
The Ni-NTA affinity-purified proteins were cleaved with SUMO protease in urea that was found to be essential for antigen solubility. Virtually 100% of all of the WUPyV and KIPyV proteins, and of BKV and JCV VP1, were cleaved in 2 M urea (Fig. 1b and d), while 10–20% of BKV VP2 and of JCV VP2/VP3 remained uncleaved in 3 M urea (Fig. 1f). The detached SUMO tag and the protease were removed by affinity subtraction (Fig. 1b, d, and f).
Immunoblotting and IFA
In all, the 100 NPA PCR-negative children showed a steady increase in VP2 and VP3 IgG seropositivities with age, reaching a combined seroprevalence of 64% for KIPyV and WUPyV (Table 2) by the age of 4 y. Of the 25 adults, 80% of the subjects were IgG-positive for one or more of the KIPyV or WUPyV antigens. A representative immunoblot is shown in Fig. 3. Of note, the IB reactivity of VP2/VP3 was superior to that of VP1; the WUPyV and KIPyV VP1 seropositivities were only 4% and 4% in children aged 0–1 y, and 8% and 12% in adults, respectively. To confirm our results, all 3- to 4-year-old children were also immunoblotted with insect cell-derived WUPyV VP1 and VP3 proteins. The results were identical to those seen with the prokaryotically-expressed proteins. In the entire study, only two serum samples with KIPyV/WUPyV IgG were reactive with all three capsid proteins. In contrast, all VP2 IgG-positive subjects (53 WUPyV and 52 KIPyV) in the study were also VP3-positive, and vice versa.
Representative results of IgG immunoblotting of 12 human polyomavirus proteins with sera of (
Data are no. (%) of patients.
IB, immunoblot; IFA, immunofluorescence assay; ND, not determined.
Only children aged ≤ 2 y were tested with IFA, whereas adults were not tested by PCR. KIPyV and WUPyV VP2/VP3 results have been combined due to apparent antigen cross-reactivity.
Among the 50 children included in IFA testing, all 18 samples positive for WUPyV/KIPyV VP2/VP3 by immunoblotting were also positive for the corresponding conformational KIPyV or WUPyV VP1 IgG in IFA. However, the VP1-IFA IgG prevalences for KIPyV and WUPyV were significantly higher than those in immunoblotting with any of the viral antigens. Of the 50 NPA PCR-negative children aged 0–2 y, 37 (74%) were IFA IgG-positive for WUPyV VP1, and 25 (50%) were positive for KIPyV VP1 (Table 2; Fig. 4).
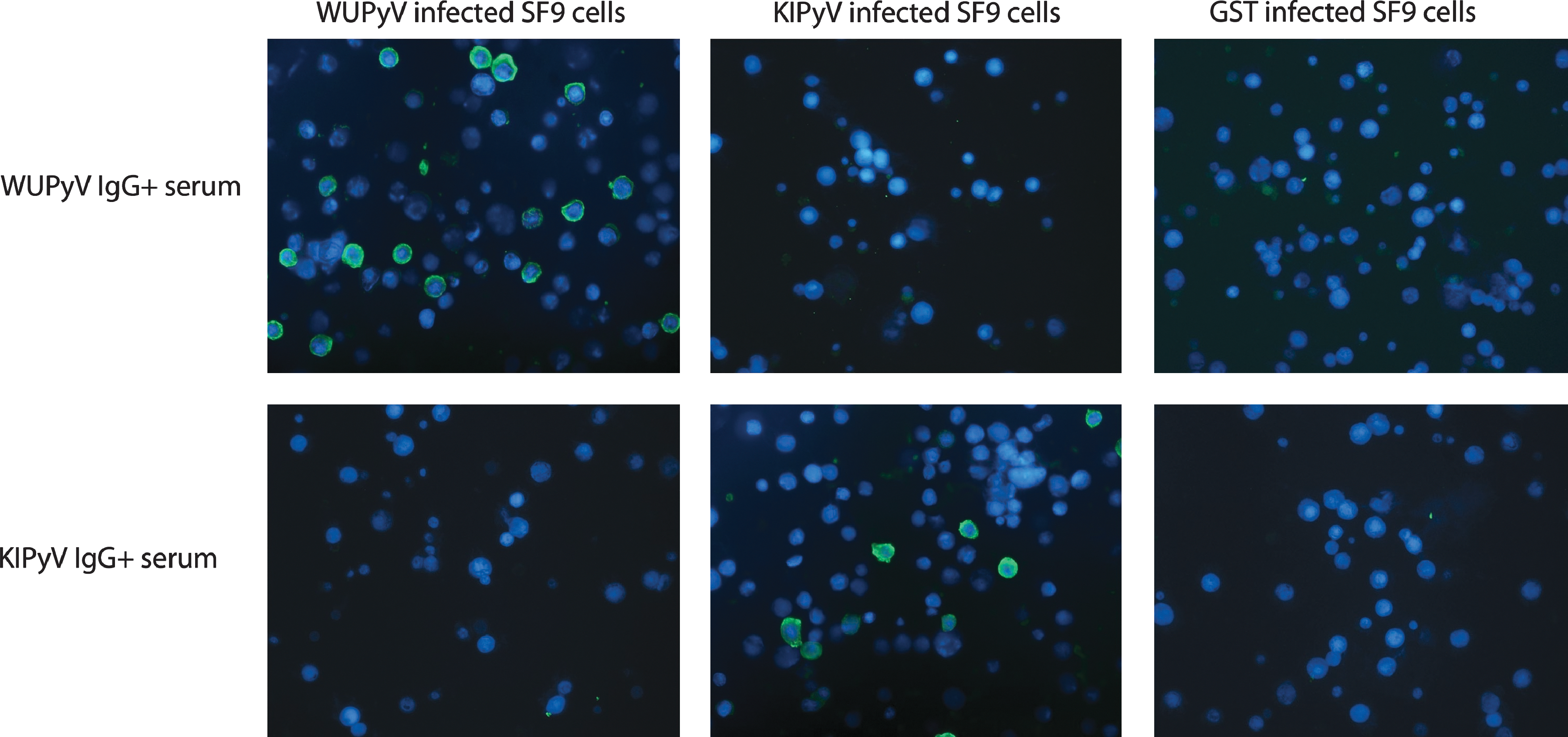
SF9 cells infected with WUPyV VP1, KIPyV, or GST encoding recombinant baculoviruses were treated with sera from wheezing children and visualized with FITC-labeled anti-human IgG (green channel) and DAPI counterstaining (blue channel). A representative set of images with all three types of cells are shown for KIPyV or WUPyV IgG-positive sera. Color images available online at
Although the numbers are too small to be statistically interpretable, IgG in IB for WUPyV or KIPyV was found more frequently in subjects with low loads of viral DNA (n = 10), than in subjects with high loads of viral DNA (n = 5; Table 3). Of the 8 and 2 PCR-positive children with WUPyV or KIPyV IgG by IB, 3 and 1 showed an IgG rise, respectively. Of these 4 IgG elevations, 3 were for VP2/VP3, and only 1 was for VP1 (WUPyV). Of the NPA PCR-positive children, only one tested positive for WUPyV VP1 IgM. VP2/VP3 IgM was not detected in this group of children.
High viral load was defined as ≥104 copies/mL of sample material, and low viral load was defined as <104 copies/mL.
Not the same child.
In order to minimize the number of true double infections, cross-reactivity between WUPyV and KIPyV in immunoblotting was only studied for the 50 youngest children (≤2 y). The number of WUPyV VP2/VP3 IgG-positive cases was significantly higher (p = 0.002 by Fisher's exact test) among the KIPyV VP2/VP3 IgG-positive samples (n = 9; 6 WUPyV IgG-positive cases), than among the KIPyV IgG-negative samples (n = 41; 5 WUPyV IgG-positive cases), and vice versa. In the entire cohort of 140 serum samples including all children and adults, all 16 serum samples strongly seropositive for WUPyV were also positive for KIPyV. Similarly, all 14 serum samples strongly seropositive for KIPyV were also positive for WUPyV. Fig. 3a shows a typical IgG IB for serum that was strongly positive for KIPyV VP2/VP3, and also positive for WUPyV VP2/VP3, albeit with reduced band intensity. In contrast, only 1 of 14 WUPyV or KIPyV VP1 IgG-positive subjects showed IgG for both viruses.
To study possible interrelations between the immunoreactivities of WUPyV/KIPyV and the two prototypical human polyomaviruses, BKV and JCV, 40 children aged ≤2 y were chosen for IgG IB. The panel included 5 sera with IgG for KIPyV/WUPyV VP1, and 13 sera with IgG for WUPyV/KIPyV VP2/VP3. Only two of these sera with KIPyV/WUPyV IgG showed a positive result with the corresponding BKV or JCV antigens. On the other hand, of 24 sera that were IgG-negative for KIPyV and WUPyV, 13 showed IgG for BKV/JCV VP1. Of these 13 VP1-positive sera, 8 were IgG-positive for all three BKV/JCV proteins, indicating a lack of serological cross-reactivity between BKV/JCV and KIPyV/WUPyV proteins. Of note, none of the sera with BKV and/or JCV IgG reacted only with VP2/VP3.
Discussion
We describe a method for the prokaryotic expression and purification of all three structural proteins of each of four human polyomaviruses, including the recently discovered WUPyV and KIPyV, with high yield and >95% purity. The proteins were expressed in E. coli using SUMO protein as a fusion tag to enhance expression levels. The SUMO fusion partner was efficiently cleaved from the KIPyV and WUPyV VP1, VP2, and VP3 to yield fully tag-free recombinant proteins, and was then depleted by a second affinity purification. The proteins were immunoblotted for antibodies of wheezing young children and asymptomatic adults, and the children's sera and NPA were examined for KIPyV and WUPyV DNA by PCR. For comparison, serum samples from the youngest children were further examined by IFA with non-denatured VP1 proteins.
Our immunoblot results showed a strong increase in WUPyV and KIPyV VP2/VP3 seroprevalence with age. By contrast, the VP1-IgG prevalence with the same method was very low, with little difference between the age groups. Interestingly, Nguyen et al. (24) previously reported that some randomly selected sera positive for KIPyV or WUPyV VP1 IgG by ELISA were also positive by immunoblotting. Whether this difference is due to a different tagging system (SUMO versus GST), technical factors, or potential restoration of antigen-antibody binding (9) is unknown. Antigen conformation has previously been shown to affect antibody recognition of polyomavirus antigens (5,9,28). Babé et al. (5) examined five monoclonal antibodies for SV40 VP1 that neutralized viral infectivity and were unreactive in Western blotting, strongly suggesting that they recognize only conformational epitopes. More recently, Bodaghi et al. (7) demonstrated that there are at least two populations of anti-VP1 IgG: the major population that almost exclusively recognizes the VLP form, and the minor population that recognizes the undenatured monomeric form.
The apparent importance of VP1 protein conformation for antibody binding renders immunoblotting an inadvisable method for VP1-based polyomavirus serodiagnostics, and underlines the requirement of correctly folded virus-like particles (VLPs) or VP1 pentamers in sensitive and virus-specific antibody detection. Indeed, conformational VP1 pentamers were recently shown to be highly reactive in WUPyV and KIPyV EIAs (16). Therefore, compared with VP1, it is interesting that the minor proteins VP2 and VP3 were found in our study to be immunogenic, even in denatured form. Based on crystallographic studies, VP2 and VP3 are thought to line the interiors of the polyomavirus pentamers (12). The minor proteins are probably inaccessible to antibodies unless (i) the polyomavirus capsid structure, and hence the tertiary structure of the antigens, is disassembled, or (ii) the peptide sequence is partially exposed (e.g., through the 12.5-Å gap in the pentameric axial cavity) (8). The steric accessibility of the minor proteins could indeed be reflected by the ability of antibodies to recognize their non-conformational (linear) epitopes in immunoblotting. On the other hand, the fact that all our KIPyV or WUPyV VP2 IgG-positive sera were also VP3 IgG-positive, and vice versa, raises the possibility that the VP2 unique region itself does not contain immunodominant B-cell epitopes, as VP3 is an N-terminal truncation of VP2. Our IFA data disclosed a ubiquitous IgG prevalence even in young children, for conformational WUPyV and KIPyV VP1. This contrasts with the immunoblot results, in which the VP2/VP3 seroprevalences increased with age. It is tempting to hypothesize that the KIPyV and WUPyV VP2/VP3 seroreactivities might evolve by repeated exposure to these viruses, possibly including reinfections or reactivations.
In all, our data strongly suggest cross-reactivity between KIPyV and WUPyV with regard to IgG for denatured VP2/VP3. However, in light of the similarity of the WUPyV and KIPyV genoprevalences, the frequent detection of IgG for VP2/VP3 in this study, and of IgG for soluble VP1 reported here and previously (16), increasing evidence is accumulating that these novel polyomaviruses circulate widely. As virtually all our sera with IgG for BKV or JCV VP2/VP3 were unreactive with the KIPyV and WUPyV VP2/VP3 antigens in immunoblotting, it is unlikely that the antibodies of the former would have affected our results. Although our attempts to verify the absence of cross-reactivity between KIPyV/WUPyV and BKV/JCV using competition by soluble proteins were not successful (data not shown), probably due to the immunoreactive linear epitopes being cryptic in solution, our results are in accordance with those of a previous study showing no cross-reactivity between conformational VP1 of these two virus pairs (16).
We showed that antibodies against the yeast SUMO protein are ubiquitous in the adult population, whereby we specifically removed the tag from our antigens. Although not proven, it is unlikely that many people would have antibodies against the human counterpart of this fusion tag, which in protein expression should provide the same benefits as the yeast analogue. On the other hand, the option to generate fully tag-free proteins by SUMO protease treatment should permit the use of these proteins in structural basic research, such as in vitro assembly of VLPs (26). However, yeast SUMO technology, as opposed to GST tagging (16,24), does not appear to offer advantages for the production of soluble and correctly-folded WUPyV or KIPyV VP1.
No overrepresentation of IgG was observed among children with a high load of viral DNA in NPA, although the number of DNA-positive patients in this study alone is too small to draw definitive conclusions. For comparison, in a previous publication we demonstrated that the level of human bocavirus (HBoV) DNA in the nasopharynx correlates strongly with the acuteness of HBoV infection, as shown serologically by immunoblotting (15), which inspired us to use similar methodology in this study. With KIPyV and WUPyV, high viral loads may indicate primary infection, together with slow appearance of IgG post-infection. Alternatively, low or moderate levels of viral DNA in NPA may reflect latent or persistent infection, rather than exclusively ongoing primary infection (1).
Our PCR results, together with the high prevalence of IgG antibodies shown here and elsewhere (16,24), suggest that KIPyV or WUPyV primary infections in immunocompetent individuals may not in general be viremic, or that the viremias may be of very short duration. Indeed, our results agree with those of a recent study of a mixed population of immunocompetent and immunocompromised individuals, that showed WUPyV viremia in only one patient with less than 1010 copies/mL in NPA, and with an unknown immune status (22).
Conclusion
We demonstrated an efficient method for the expression of KIPyV and WUPyV structural proteins for immunological and serodiagnostic purposes. Comprehensive serodiagnosis of KIPyV and WUPyV when using denaturing techniques calls for the inclusion of VP2 and/or VP3 in addition to VP1. KIPyV- and WUPyV-related viremias occur rarely or are short-lived in immunocompetent individuals.
Footnotes
Acknowledgments
We wish to thank Dr. Kristina Dörries of the Julius Maximilian University for the kind gift of BKV and JCV clones. This work was supported by the Helsinki University Central Hospital Research & Education and Research & Development funds (to A.L.), the Centre for International Mobility (CIMO; to A.K.), the Sigrid Jusèlius Foundation, the Finnish Funding Agency for Technology and Innovation, and the Academy of Finland (to K.K., M.S., K.H., and M.S.V.; project code 1122539), the University of Helsinki Funds, the Paulo Foundation, and the Research and Science Foundation of Farmos and BONFOR (to P.N.; grant number 0-151.0021), the Swedish Research Council, and the Torsten and Ragnar Söderberg Foundation (to T.A.).
Author Disclosure Statement
No competing financial interests exist.