
Abstract
Recently HIV-infected individuals have virus-specific responses characterized by IFN-γ/IL-2 secretion and proliferation rarely seen in chronic infection. To investigate the timing of loss of HIV-specific T-cell function, we screened cells from 59 treatment-naïve HIV-infected individuals with known dates of infection for proteome-wide responses secreting IFN-γ/IL-2 and IFN-γ alone by ELISPOT. HIV peptide-specific proliferation was assessed by carboxyfluorescein diacetate succinimidyl ester (CFSE) dilution. The contribution of IFN-γ/IL-2 and IFN-γ-only secretion to the total HIV-specific response was compared in subjects infected <6, 6–12, and 12–36 mo earlier. The frequency of IFN-γ/IL-2-secreting cells fell, while that of IFN-γ-only secretion rose with time from infection. HIV peptide-specific proliferative responses were almost exclusively mediated by CD8+ T cells, and were significantly lower in cells obtained from the 12–36 mo versus < 6 mo post-infection groups. By the second year of infection there was a significant difference in these functions compared to those assessed within 6 mo.
Introduction
It has been suggested that T cells with multiple functions, including IFN-γ and IL-2 secretion, and that have the ability to proliferate, are better at controlling experimental viral infections than T cells with more limited function (58). Several studies have associated HIV-specific polyfunctional T cells with better disease control (9,30). Additionally, these cells have been associated with protection from infection with pathogenic Leishmania major, and are induced by vaccinia and yellow fever vaccine (22,53,54). HIV-specific CD8+ T-cell response magnitude and breadth, as detected using IFN-γ secretion assays and tetramer reagents, is compromised in acute infection (6,7,20,38,65,66). HIV-specific T cells able to proliferate and secrete both IFN-γ and IL-2 develop their full breadth and magnitude after the acute phase, and are detected in most infected individuals within the first 6 mo of infection (37,49,66). Despite the presence of these cells, most untreated HIV-infected subjects exhibit disease progression, and polyfunctional responses are rarely seen in subjects with chronic infection, except for a subset of HIV-infected individuals who spontaneously control viremia without treatment (4,9,30,37,51,55,64). In contrast, secretion of IFN-γ by HIV-specific cells, although compromised in acute infection, persists once induced well into the chronic phase of infection (1,6 –8,21,50). This is analogous to what occurs in other chronic viral infections such as murine clone 13 lymphocytic choriomeningitis virus infection, in which antigen-specific IL-2 secretion is one of the first functions of memory T cells lost, whereas IFN-γ secretion is more resistant to extinction (28,63). The cause-and-effect relationship between loss of IL-2 secretion and proliferative capacity and high VL remains a topic of investigation. Although HIV-specific cells able to secrete both IFN-γ and IL-2, or IL-2 only, and to proliferate are lost as disease progresses, there is little information available addressing the timing of this loss.
In this article we sought to assess the functional evolution of HIV-specific responses in terms of the timing of loss of HIV-specific responses characterized by secretion of IFN-γ and IL-2 versus IFN-γ alone. We used a dual-color ELISPOT assay able to simultaneously detect three functional lymphocyte populations: cells secreting IFN-γ/IL-2, IFN-γ only, and IL-2 only. We screened peripheral blood mononuclear cells (PBMCs) from 59 treatment-naïve subjects infected for less than 36 mo for responses directed to the entire HIV proteome. Having previously determined that the contribution of dual IFN-γ/IL-2-secreting cells was greater, and single IFN-γ-secreting cells lower in subjects infected <6 mo versus those in the chronic phase of infection, the timing of change of HIV-specific functional responses characterized by the secretion of IFN-γ/IL-2 and IFN-γ only was ascertained by comparing the contribution of each HIV-specific functional subset to the total response in subjects sampled within 6 mo (n = 35), 6–12 mo (n = 19), and 12–36 mo (n = 35) of infection. Because the proliferative capacity of HIV-specific T cells is linked to their ability to secrete and maintain IL-2, we also assessed proliferation to HIV peptides of cells obtained within 6 mo and between 12 and 36 mo of infection. We found that the contribution of HIV-specific responses secreting IFN-γ and IL-2 and able to proliferate was lower in individuals tested between 12 and 36 mo of infection than in those infected <6 mo, and that this difference was even more important for peptides restricted by HLA alleles not associated with protection.
Methods
Study population
The study population included 59 treatment-naïve HIV subtype B-infected subjects. All participants provided informed consent and the institutional review boards of all participating study sites approved this study. All but two study subjects were drawn from the Montreal Primary Infection (PI) cohort, which enrolls individuals within 1 y of infection and follows them for 2 y. The two individuals not enrolled in the Montreal PI cohort, MQ13 and MQ16, who were infected for 12 and 20 mo, respectively, were also treatment-naïve individuals with known dates of infection followed at the Immune Deficiency Treatment Centre clinic of the McGill University Health Centre. The date of infection was estimated for each participant using the algorithm proposed by the Acute HIV Infection Early Disease Research Program sponsored by the National Institutes of Health (5,39,49). Of the 59 individuals studied, 31 were tested more than once, such that there were samples from 35 individuals (33 males) screened within 6 mo, 19 individuals (18 males) between 6 and 12 mo, and 35 individuals (34 males) between 12 and 36 mo of infection. Ten subjects, some with time points in more than 1 study group, i.e., 6 in the <6 mo, 4 in the 6–12 mo, and 7 in the 12–36 mo post-infection groups expressed either the HLA-B27 or HLA-B57/B58 alleles previously reported to be associated with slow HIV disease progression (16,31,62).
Laboratory testing
HIV-1 enzyme immunoassay (EIA) antibody testing was performed at three university hospital sites. Western blot analysis for HIV-1 antibodies and p24 antigen capture assays were performed at the Laboratoire de Santé Publique du Québec (Ste. Anne de Bellevue, Québec, Canada). The less sensitive EIA (detuned assay), which is used to confirm timing of infection, was carried out at the University of California–San Francisco, using the Vironostica HIV-1 EIA (Organon-Teknika, Boxtel, the Netherlands). Plasma viremia was measured using the ultra-sensitive method (Ultradirect; Roche Diagnostics Corp., Indianapolis, IN), with a detection limit of 50 HIV-1 RNA copies/mL of plasma.
Cell preparation
PBMCs were obtained by venipuncture in tubes containing EDTA anticoagulant or by leukapheresis (12). PBMCs were isolated by density gradient centrifugation (Ficoll-Paque; Pharmacia, Uppsala, Sweden) and cryopreserved in 10% dimethyl sulfoxide (Sigma-Aldrich, St. Louis, MO) and 90% fetal bovine serum (Medicorp, Montréal, Québec, Canada).
Stimulatory peptides
Peptides (15-mer with 11-aa overlaps from the National Institutes of Health AIDS Research and Reference Reagent Program, Rockville, MD) corresponding to the HIV consensus clade B sequence spanning the entire HIV proteome were used to stimulate the PBMCs. Pools containing 2–15 peptides were prepared and organized into matrices of Gag, Pol, Nef, Env, and accessory gene peptide pools such that each peptide was present at a final concentration of 4 μg/mL in two pools within each peptide matrix. An initial peptide pool matrix strategy was used to identify candidate stimulatory peptides, followed by a verification assay in which candidate peptides were tested individually at 4 μg/mL to confirm their stimulatory capacity.
ELISPOT assay for dual IFN-γ/IL-2, single IFN-γ, and single IL-2 cytokine release
IFN-γ and IL-2 secretion by HIV-specific cells was quantified using a dual-color ELISPOT assay as described elsewhere (13). Briefly, the wells of a polyvinylidene difluoride-backed microtiter plate (Millipore, Billerica, MA) were coated with optimized concentrations of capture antibodies for both IFN-γ and IL-2 (Becton Dickinson [BD] Pharmingen, San Diego, CA). The PBMCs were thawed and plated at 2 × 105 cells per well and stimulated with peptide pools for 28 h. Cytokine secretion from individual cells was detected by sequential addition, followed by washing with a detection antibody cocktail containing a fluorescein isothiocyanate (FITC)-conjugated anti-IFN-γ antibody (Ab) (1-D1K; Mabtech, Nacka Strand, Sweden), and a biotin-conjugated anti-IL-2 Ab (BD), followed by a cocktail of horseradish peroxidase-conjugated anti-FITC monoclonal Ab (Jackson ImmunoResearch Laboratories, West Grove, PA) and streptavidin alkaline phosphatase (Jackson ImmunoResearch Laboratories). Spots corresponding to the footprint of IFN-γ/IL-2, IFN-γ, and IL-2 cytokine-secreting cells were developed by the addition of Vector Blue (Vector Laboratories, Burlingame, CA), and amino ethyl carbazole (Sigma-Aldrich) substrates. Purple (IFN-γ and IL-2 cytokine secretion), red (IFN-γ secretion), and blue (IL-2 secretion) spots in each well were counted using a CTL series 3B Immunospot analyzer (CTL Analyzers LLC, Cleveland, OH) with software version 4.0 supplied by the manufacturer. Results are expressed as spot-forming cells per million PBMCs (SFC/106 PBMCs) following subtraction of negative controls. To establish the threshold of detection of positive responses for the single and dual cytokine-secreting responses, PBMCs from 8 HIV-seronegative low-risk individuals were stimulated with the same set of stimuli as the test subjects. Uninfected individuals produced a mean of 4.2 ± 10.6 IFN-γ SFC/106 PBMCs to this peptide platform. A positive response was defined as 3 standard deviations above the mean of that seen for uninfected subjects (i.e., >36 SFC/106 PBMCs), and at least threefold greater than the autologous negative control wells. Using the same rationale, the thresholds for IL-2 and dual-cytokine secretion were set at >32 SFC/106 PBMCs and >21 SFC/106 PBMCs, respectively, and at least threefold greater than the autologous negative control wells.
5,6-Carboxyfluorescein diacetate succinimidyl ester (CFSE) dilution assay
PBMCs were thawed, washed, resuspended in cRPMI supplemented with 2% human AB serum (Wisent Inc., St-Bruno, Quebec, Canada), and labeled with 5 μM CFSE (Molecular Probes, Eugene, OR). CFSE-labeled PBMCs were stimulated in cRPMI containing 15% human AB serum with 2 μg/mL of HIV consensus B peptides identified during the screening process using the ELISPOT assay. Stimulation with media alone served as a negative control, whereas stimulation with 1 μg/mL of staphylococcal enterotoxin B (SEB) and 1 μg/mL of human cytomegalovirus lysate (Advanced Biotechnologies Inc., Columbia, MD) were used as positive control stimuli. Following 6 d of incubation at 37°C and 5% CO2, PBMCs were stained for cell surface markers using anti-CD3-peridinin chlorophyll protein, anti-CD8-allophycocyanin, and anti-CD4-phycoerythrin mAbs (BD). One hundred thousand lymphocyte-gated events were acquired per sample using a FACSCalibur flow cytometer, and analyzed with FlowJo software version 8.7.1 (Tree Star, Inc., Ashland, OR). Information on the percentage and number of CFSElo cells in the CD4 and CD8 compartments was collected. Responses were defined as positive if after subtracting background they were above 0.1%.
Statistical analysis
Statistical analysis and graphical presentation were performed using InStat 3.05 and GraphPad Prism 4.01, respectively (GraphPad Software, San Diego, CA). The Kruskal-Wallis test with Dunn's post-test for multiple comparisons was used to assess between-group differences in age, CD4 counts, CD8 counts, VL, response magnitude, and contribution of each functional subset to the total HIV-specific response. The significance of between-group differences in peptide-specific proliferation of samples from two time points obtained within 6 mo and later in infection (between 8 and 17 mo of infection) was assessed using a Mann-Whitney U test for non-matched samples, and a Wilcoxon matched-pairs signed-rank test for paired samples. A Fisher's exact test was used to assess proportional between-group differences. Statistical significance was set at p < 0.05.
Results
Study population
Table 1 shows for each group the median and interquartile range (IQR) of time from infection at which samples were obtained. Also shown in Table 1 are the median (IQR) age, absolute CD4 and CD8 counts, and HIV log10 VL measures of the subjects included in each time-interval-from-infection group. There were no between-group differences in age, absolute CD8 counts, and VL (p > 0.05 by Kruskal-Wallis test). Compared to subjects who were infected <6 mo, those infected 12–36 mo had lower CD4 counts (p < 0.001 by Dunn multiple comparisons test).
Median (interquartile range).
M, male; F, female.
Changes in the percentage contribution of HIV-specific IFN-γ/IL-2- and IFN-γ-only-secreting cells with increasing time from infection
Results generated using a dual-color ELISPOT assay screening for proteome-wide HIV-specific responses provided information on the specificity, the breadth as defined by the number of individual stimulatory peptides recognized, and the magnitude of HIV-specific immune responses as defined by the sum of SFC/106 PBMCs induced by each stimulatory peptide for each of the cytokine secretion profiles detected by the assay. In previous work we showed that HIV-specific cells secreting IL-2 were only present at low frequencies and targeted few epitopes, even in individuals infected <6 mo, and were rarely detected in individuals during chronic infection (49). For this reason, in this article we focused on dual IFN-γ/IL-2 and single IFN-γ responses, and sought to assess their functional evolution during the first 3 y of infection. The magnitude of HIV-specific responses characterized by IFN-γ/IL-2 secretion and single IFN-γ secretion did not differ significantly between the three groups defined by time from infection (Fig. 1A and B; p > 0.05 by Kruskal-Wallis test). However, the proportion of HIV-specific cells secreting IFN-γ and IL-2 decreased with time, from a median (IQR) of 21.5% (12,29) to 11% (2,20.5), and 6% (0,22) for the groups infected <6 mo, 6–12 mo, and 12–36 mo, respectively (Fig. 1C; p = 0.019 by Kruskal-Wallis test). The proportion of HIV-specific cells secreting IFN-γ alone increased with time from infection from 78% (71,88) to 88% (78.5,98), and 94% (78,100) in the groups infected <6 mo, 6–12 mo, and 12–36 mo, respectively (Fig. 1D; p = 0.01 by Kruskal-Wallis test). The contribution of HIV-specific responses to the total response for IFN-γ/IL-2-secreting cells was significantly greater, and for IFN-γ only responses significantly lower, for comparisons between the groups infected <6 mo versus 12–36 months (Fig. 1C and D; p < 0.05 for both comparisons by Dunn's multiple comparisons test).
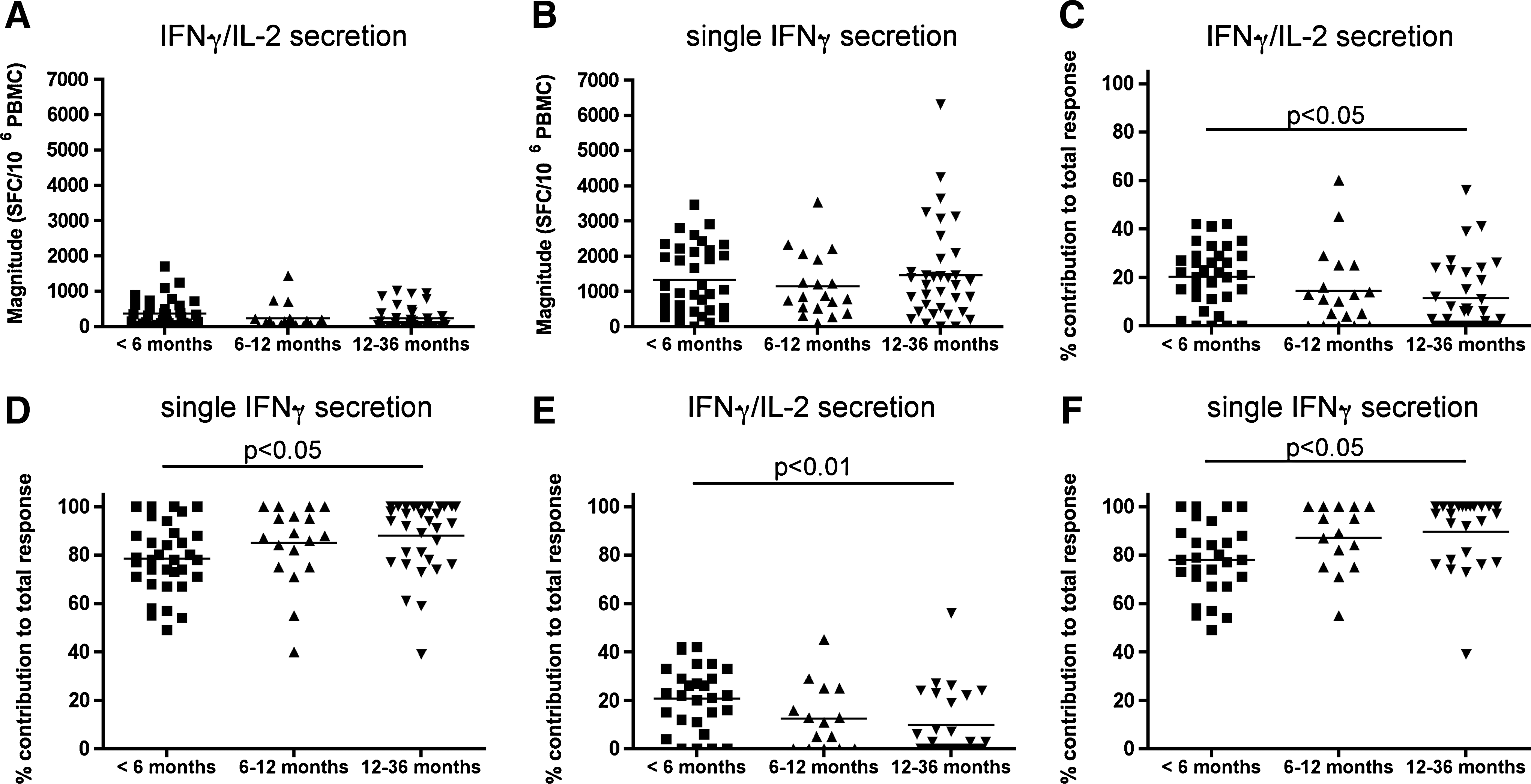
HIV-specific functional subsets in subjects categorized by duration of infection. Thirty-five subjects infected <6 mo, 19 infected between 6 and 12 mo, and 35 infected between 12 and 36 mo were screened for immune responses to the entire HIV proteome using a dual-color ELISPOT assay. The magnitude of proteome-wide HIV-specific responses characterized by secretion of IFN-γ/IL-2 is shown in panel
The study population included 10 individuals expressing either the HLA-B*27, HLA-B*57, or HLA-B*58 alleles, which are associated with slow HIV disease progression (16,31,44,62). These alleles are often present in individuals who exhibit effective VL control, and who are able to maintain HIV-specific proliferation and responses characterized by the secretion of multiple cytokines, including IL-2 (4,9,25,43,46). We reasoned that inclusion of subjects with these HLA alleles may introduce a bias towards the maintenance of HIV-specific cells able to secrete IFN-γ and IL-2. To address this, we re-analyzed the data, excluding the subjects expressing these MHC class I alleles. Fig. 1E shows that the contribution of HIV-specific cells secreting IFN-γ/IL-2 to the total responses fell from 22% (11.75,30) to 11% (0,20.5) and 3% (0,22), for subjects infected for 6–12 and 12–36 mo, respectively (p = 0.02 by Kruskal-Wallis test), while for IFN-γ only responses it rose from 78% (70,88.25) to 89% (78.5,100) and 97% (78,100) in the three study groups defined by increasing time from infection (p = 0.006 by Kruskal-Wallis test; Fig. 1F). Between-group differences were statistically significant for those infected <6 mo versus 12–36 mo for both cytokine secretion patterns (Fig. 1E and F; p < 0.01 and p < 0.05 for HIV-specific cells secreting both IFN-γ/IL-2 and IFN-γ alone, respectively, by Dunn's multiple comparisons test). In contrast, for the 6, 4, and 8 subjects infected for <6, 6–12, and 12–36 mo expressing one of the protective HLA alleles, the contribution of IFN-γ/IL-2-secreting cells to the HIV-specific response was 19% (13,25), 12% (9,25), and 12% (6,21), changes that were not significantly different (p = 0.79 by Kruskal-Wallis test).
HIV peptide-specific proliferation of PBMCs from subjects infected for defined time intervals
We used a CFSE dilution assay to assess the proliferative capacity of PBMCs specific for HIV peptides able to induce secretion of IFN-γ/IL-2 or IFN-γ only in the dual-color ELISPOT assay, to establish whether HIV-specific proliferative capacity differed in cells obtained from time points collected within 6 mo versus those taken at later times post-infection.
Flow cytometry plots showing the proliferative capacity of CD4+ and CD8+ T cells following no stimulation as a negative control, stimulation with SEB and cytomegalovirus lysate (CMV) as positive controls, and the HIV peptide pol 5543 for a study subject infected <6 mo is shown in Fig. 2A. A panel of 103 HIV peptides recognized by 17 different subjects tested within 6 mo of infection was used to induce proliferation of cells from the same study subject time point. Eighty-one HIV peptides stimulated IFN-γ/IL-2 secretion, while 22 peptides stimulated IFN-γ-only secretion in the dual-color ELISPOT assay. Of the 81 peptides stimulating IFN-γ/IL-2 secretion, 19 (23.5%) and 61 (75.3%) induced proliferation in the CD4+ and CD8+ T-cell compartments, respectively (Fig. 2B). Of the 22 peptides inducing single IFN-γ secretion, 3 (13.6%) and 11 (50%) stimulated proliferation in the CD4+ and CD8+ T-cell compartments, respectively. A larger proportion of peptides stimulating secretion of both IFN-γ/IL-2 than IFN-γ only induced CD8+ T-cell proliferation (p = 0.03 by Fisher's exact test). This comparison was not significantly different for CD4+ T-cell proliferation, possibly due to the smaller number of responses mediated by CD4+ T cells. If only the peptides inducing above-background proliferation of CD8+ T cells were considered, the median (IQR) percentage of CFSElo cells was 1.85% (0.73,5.16) and 1.09% (0.52,2.58) for peptides inducing IFN-γ/IL-2 and IFN-γ alone, respectively, a difference that was not statistically significant (p = 0.21 by Mann-Whitney U test). Therefore, HIV-specific proliferative responses detected within 6 mo of infection were generally mediated by CD8+ T cells, although some peptides induced proliferation of both CD8+ and CD4+ T cells, or of only CD4+ T cells. A larger proportion of peptides able to stimulate IFN-γ/IL-2 than IFN-γ alone were also able to induce above-background levels of CD8+ T-cell proliferation.
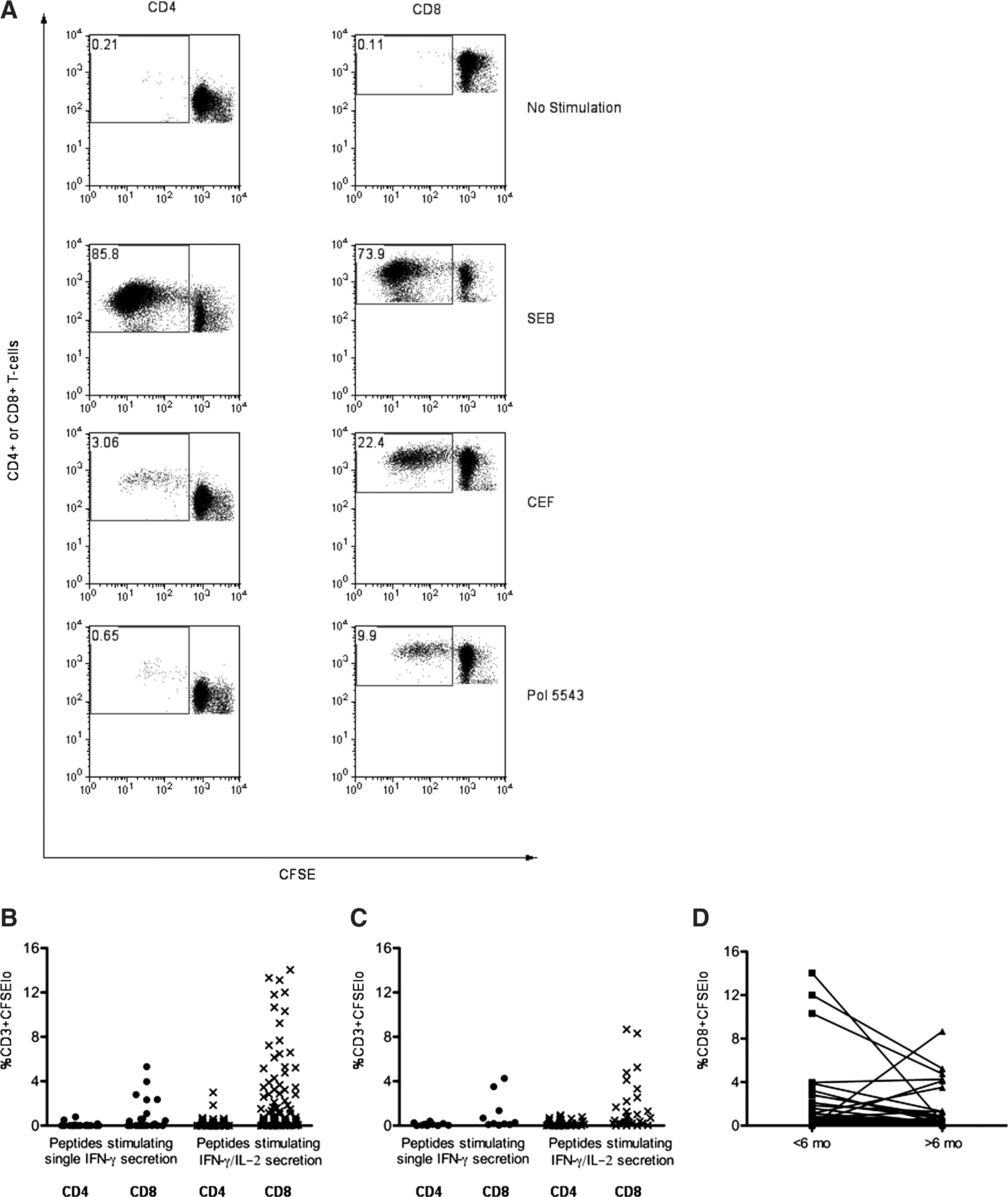
HIV-specific proliferation of CD4+ and CD8+ T cells in samples obtained at times within 6 mo and after 6 mo of infection to peptides stimulating either IFN-γ/IL-2- or IFN-γ-only secretion. Panel
Thirty-eight peptides recognized by six subjects tested after 6 mo of infection by dual-color ELISPOT were screened for their ability to stimulate proliferation of cells from the same study subject time point. These six subjects were infected 241, 356, 368, 372, 448, and 501 d at the time of testing. Twenty-eight and 10 peptides induced IFN-γ/IL-2- and IFN-γ-only secretion, respectively, in the dual-color ELISPOT assay. Of the 28 peptides inducing IFN-γ/IL-2 secretion 15 (53.6%) and 24 (85.7%) stimulated proliferation in the CD4+ and CD8+ T-cell compartments (Fig. 2C). Of the 10 peptides inducing IFN-γ-secretion only, 5 (50%) and 9 (90%) induced proliferation in the CD4+ and CD8+ T-cell compartments, respectively. There was a non-significant trend toward the percentage of CD8+ CFSElo cells being higher for responses to peptides stimulating IFN-γ/IL-2- than IFN-γ-only secretion (0.94% [0.30,2.72] and 0.29% [0.18,1.35] for peptides inducing IFN-γ/IL-2 and IFN-γ alone, respectively; p = 0.13 by Mann-Whitney U test). This was also the case for proliferation of CD4+ T cells (0.46% [0.22,0.67] and 0.19% [0.13,0.26] CFSElo cells induced by peptides stimulating IFN-γ/IL-2 and IFN-γ only, respectively; p = 0.07 by Mann-Whitney U test).
CD8+ T cells from subjects within 6 mo of infection had a greater capacity to proliferate than CD8+ T cells from individuals at later times post-infection to peptides inducing IFN-γ/IL-2 secretion (p = 0.03 by Mann-Whitney U test). This was not the case for the peptide panel inducing IFN-γ-secretion only, which stimulated similar levels of CD8+ T-cell proliferation from subjects at both time intervals post-infection (p = 0.42 by Mann-Whitney U test). As seen in Fig. 2B and C, proliferation of CD4+ T cells was of a lower magnitude than that of CD8+ T cells for both peptide panels and time-interval-from-infection groups. Comparison of the percentage of CFSElo CD4+ T cells induced by peptides stimulating IFN-γ/IL-2 revealed a non-significant trend toward greater proliferative capacity of samples collected before 6 mo versus later time points post-infection (p = 0.06 by Mann-Whitney U test), which was not seen for peptides stimulating IFN-γ secretion only (p = 0.77 by Mann-Whitney U test). In summary, this cross-sectional analysis of HIV-specific proliferative capacity showed that both CD8+ and CD4+ cells responding to peptides able to stimulate IFN-γ/IL-2 secretion in a dual-color ELISPOT assay was lower in samples collected after 6 mo of infection compared to those collected within 6 mo of infection. In contrast, the proliferative capacity of CD8+ and CD4+ cells responding to peptides stimulating IFN-γ secretion only was similar at both time intervals from infection.
For five study subjects exhibiting HIV disease progression two time points were available for testing, one from within 6 mo of infection and the other from a later time, 241, 356, 372, 448, and 501 d from infection. Twenty-seven stimulatory peptides were used to induce proliferation in paired samples tested in the same experiment. This stimulatory peptide panel induced a median (IQR) percentage of CFSElo cells of 1.09% (0.30,2.82) and 0.48 (0.20,1.33) in paired samples collected at early and late time points, respectively (p = 0.05 by Wilcoxon matched pairs signed-ranks test) (Fig. 2D). These results confirm in paired samples collected within 6 mo and later in infection that proliferative capacity to the same epitope declines with time from infection.
Discussion
In this report we assessed the contribution of proteome-wide HIV-specific responses characterized by secretion of both IFN-γ and IL-2 and IFN-γ only at three time intervals post-infection in order to address the timing of loss of HIV-specific responses able to secrete IFN-γ/IL-2. Thirty-one of the 35 (88.6%) subjects tested within 6 mo of infection had HIV-specific cells capable of secreting IFN-γ and IL-2 simultaneously. While the absolute magnitude of HIV-specific responses secreting both IFN-γ and IL-2 decreased, and that secreting IFN-γ alone increased with time from infection, this change did not achieve statistical significance when a Kruskal-Wallis test was used to assess the significance of differences in subjects tested within 6 mo versus those tested 6–12 or 12–36 mo post-infection. However, the relative contribution of responses characterized by IFN-γ/IL-2 secretion fell, while that for IFN-γ-only secretion rose with increasing time intervals post-infection. Using panels of HIV peptides able to stimulate either IFN-γ/IL-2 or IFN-γ alone to induce proliferation of cells from the same time point used for the ELISPOT assay, over 75.3% of peptides stimulating IFN-γ/IL-2 secretion and over 50% of peptides stimulating IFN-γ-only secretion also induced proliferation of CD8+ T cells. A lower proportion of these peptides stimulated proliferation of both CD8+ and CD4+ T cells, or stimulated only CD4+ T-cell proliferation. Proliferative capacity was greater for CD8+ than CD4+ T cells, and was generally higher in cells from time points collected within 6 mo of infection than from times spanning 241–501 days from infection, particularly for peptides able to induce IFN-γ/IL-2 secretion. The observation that proliferation was greater for CD8+ than for CD4+ T cells could be due to the use of 15-mers as a stimulus. Although 15-mers are known to induce CD4+ T-cell responses, the use of 20-mers may be more appropriate to accurately measure their immunological response profile (24). An alternative possibility is that the low level of proliferation to HIV peptides in the CD4 compartment may reflect a defect in CD4+ T-cell proliferative capacity.
The rationale for choosing to examine HIV-specific IFN-γ secretion in this study population is that this T-cell function is resistant to functional exhaustion in a setting of high viremia, and it is the cytokine most frequently assessed in standard ELISPOT assays that measure HIV-specific immune responses (60,63). The decision to assess HIV-specific IL-2 secretion was based on its sensitivity to functional exhaustion in a setting of high viremia (30,63,68). HIV-specific IFN-γ/IL-2-secreting cells are present in most individuals in early infection, but are uncommon in the chronic phase of progressive infection (37,49). By monitoring HIV-specific IFN-γ/IL-2-secreting cells in samples collected at defined times from infection it is possible to address when after infection the response characterized by this functional profile begins to wane. Although we only assessed IFN-γ and IL-2 secretion and proliferation, it is likely that the IFN-γ/IL-2 functional profile is a surrogate for cells with a more multifunctional profile. Most of the HIV-specific responses detected in long-term non-progressors that were able to secrete IL-2 also mediated the secretion of other cytokines and/or chemokines (9). The functional importance of IL-2 secretion relates to its role in the maintenance of CD8+ T-cell responses, proliferation, and differentiation into effector T cells when CD4+ T-cell help is limited (29,30,37).
Our observation that cells from individuals in early infection have a higher proliferative capacity than cells from later times in infection is likely to be related to changes in the functional avidity and clonal make-up of HIV-specific responses with increasing time from infection (38). Lichterfeld et al. found that responses characterized by CD107a expression and secretion of IFN-γ and MIP-1β were of a higher functional avidity in individuals in acute infection with early disease, such as the subjects tested here infected <6 mo, than those to the same peptides seen in subjects infected for more than 3 y (38). Some responses were no longer detected at later times in infection, or their contribution to the total response was reduced due to the expansion of T cells bearing T-cell receptor clonotypes having low to medium avidity for their cognate antigen (38). Alternatively, a recent study has shown that the persistent antigenic load that characterizes chronic infections drives T-cell functional exhaustion through sustained antigen presentation (48).
An important proportion of the peptides inducing single IFN-γ secretion in the dual-color ELISPOT assay were also able to induce proliferation. In general, responses that include the ability to secrete IL-2 rather than IFN-γ only are correlated with proliferative capacity (37,67). The reason underlying the finding that many peptides stimulating secretion of IFN-γ only also stimulated proliferation may be related to the sensitivity of the dual-color ELISPOT assay. In other words, these responses may include cells able to secrete enough IL-2 to support proliferation, but at a frequency or in a quantity insufficient to produce a response above the cut-off for background in the dual-color ELISPOT assay. This potential limitation of the dual-color ELISPOT assay may also apply to detection of cytokine-secreting cells by intracellular cytokine staining, which has a lower threshold of detection of 0.02% specific T cells (61).
Lichterfeld et al. reported that the magnitude of responses characterized by the secretion of not only IFN-γ, but also MIP-1β and CD107a/b expression, was of a lower magnitude in subjects infected <6 mo compared to those infected for more than 3 years (38). We did not observe differences in the magnitude of either IFN-γ or IFN-γ/IL-2 secretion for the three study groups categorized by time from infection studied here. This discrepancy may be explained by the early-infection group studied by Lichterfeld et al., which included a larger proportion of individuals infected <3 mo than the <6-mo-infected population studied here. We and others have reported that the breadth and magnitude of responses in individuals infected <3 mo is compromised compared to that seen in individuals in later stages of primary infection (6,7,21).
By the second half of the first year of infection it is already possible to detect a decline in the contribution of HIV-specific cells secreting both IL-2 and IFN-γ. The contribution of IFN-γ/IL-2-secreting HIV-specific cells continues to fall in the second year of infection, but appears to plateau thereafter, as we did not detect significant differences in the contribution of IFN-γ/IL-2- and single IFN-γ-secreting cells between individuals tested between 12 and 24 mo and 24 and 36 mo of infection (data not shown). Secretion of IL-2 is linked with proliferative potential, which is lost in the majority of viremic individuals during chronic HIV infection (37,51,55,67). The loss of IL-2 secretion and persistence of IFN-γ secretion are a functional phenotype for an effector population having limited self-renewal potential. This likely has negative consequences for control of persistent viral replication. HIV-specific cells secreting IL-2 and able to proliferate also have enhanced cytolytic ability, adding to the protective potential ascribed to polyfunctional lymphocytes more likely to be seen in early infection (37,38,43,59,63).
In summary, we show that most individuals infected <6 mo develop HIV-specific IFN-γ/IL-2 responses in addition to single IFN-γ responses. These bifunctional responses peak within the first 6 mo of infection and begin to wane soon thereafter.
Footnotes
Acknowledgments
We wish to acknowledge Ms. Chantal Grignon and Ms. Melanie Lemay for nursing support for the leukapheresis infrastructure, Mr. Mario Legault, coordinator of the Montreal Primary Infection cohort, and Ms. Nancy Simic and Ms. Marie-Pierre Boisvert for expert technical assistance. We acknowledge Dr. Bernard Lessard, Dr. Anne Talbot, Dr. Marc-André Charon, Dr. Martin Potter, Dr. Marina Klein, Dr. Norbert Gilmore, Dr. John MacLeod, Dr. Graham Smith, Dr. Benoit Trottier, Dr. Louise Charest, Dr. Danielle Legault, Dr. Michel Boissonnault, Dr. Claude Fortin, Dr. Julian Falutz, and Dr. Julie Bruneau for study subject referral, enrollment, and clinical follow-up. We are also grateful to participants in the Montreal PI cohort. This work was supported by grants from the National Institutes of Health #AI043261-04, and the Réseau du Sida et Maladies Infectieuses du Fonds de la Recherche en Santé du Québec (FRSQ). Y.P. and S.B. were the recipient of a scholarship from the FRSQ, and J.-P.R. and was the recipient of a Chercheur Boursier scholarship from the FRSQ.
Author Disclosure Statement
No competing financial interests exist.