
Abstract
Infection with porcine reproductive and respiratory syndrome virus (PRRSV) results in a weak antiviral immune response that leads to a persistent infection in a subset of pigs. We investigated the intensity and timing of the early cytokine responses to PRRSV infection to determine their utility as a predictor of persistence. As part of the “Big Pig” project, we evaluated cytokine gene expression in lymphoid tissues collected from pigs for up 202 days post-infection (dpi); serum samples were collected biweekly. Cytokine mRNA levels were compared between pigs that cleared the viral infection from serum and tissues (non-persistent [NP] pigs) to those of persistent (P) pigs, that had viral RNA in their serum for up to 126 dpi. The gene expression studies in the tracheobronchial lymph nodes (TBLN) of all the pigs showed upregulation of interferon-γ (IFN-γ)-associated T-helper 1 (Th-1) markers from 14–84 dpi, and of T-regulatory interleukin-10 (IL-10), but no upregulation of innate markers (IFN-A, IL-1B, and IL-8). At later time points (>112 dpi) these genes were no longer differentially expressed and thus were uninformative for persistence studies. Statistical analyses of serum cytokine levels indicated that innate cytokine (IL-1β and IL-8) levels were upregulated early after infection. Interestingly, serum IL-8 levels in NP pigs were significantly higher than in P pigs at 14 dpi. When analyzed together, variations in all three of the serum cytokines tested (IL-8, IL-1β, and IFN-γ) was significantly correlated with virus level, accounting for ∼84% of the variations observed. These results indicate that while each cytokine individually has minor effects on the length of virus replication, the combination of cytokine activities should be considered when understanding the role of immunity in persistence.
Introduction
Horter et al. (12,13) found that infectious PRRSV is present in most pigs for the first 105 dpi, and suggested that RT-PCR assays of oropharyngeal scrapings was the most effective combination of assay and sample for detecting PRRSV carriers. They noted that antibody response levels did not distinguish PRRSV carrier from non-carrier animals. Serum virus neutralizing antibodies (SNA) are important in PRRSV control during acute infection (18,25), but polyclonal B-cell activation also occurs (15,16,27). Moreover, Batista et al. (2) confirmed that none of an extensive set of parameters (serum ELISA antibody, SNA, and interferon-γ [IFN-γ] levels assessed using ELISpot and flow cytometry, or the proliferative responses of CD4+ and CD8+ T lymphocytes), either alone or in combination, was definitively associated with (or predictive of) viral clearance and sterilizing immunity. PRRSV persists in the serum, despite SNA, and in the lymphoid tissue even after IFN-γ levels have dropped, indicating that some other immune factor is associated with viral clearance and sterilizing immunity after infection (2,20). In field studies, Lowe et al. (19) noted the importance of cell-mediated immunity (as assessed by IFN-γ levels using ELISpot) in preventing piglet losses from sows, although genetic factors also were noted. CD8+ T-cell depletion had no effect on PRRSV resistance (17).
In 2005, a large study known as “Big Pig” was initiated to understand the role of population size on the outcome of PRRSV infection (22,23). One hundred and nine pigs were infected with VR-2332, and a subset of pigs was followed for up to 202 dpi. The samples collected during the Big Pig study provided sera and tissues so that we could determine whether immune factors, and particularly immune cytokines (interleukin-1β [IL-1β], IL-8, IL-10, and IFN-γ), help determine why certain pigs maintain PRRSV replication for extended periods.
Materials and Methods
Animal care, handling, sampling, and database
As previously described by Molina et al. (22,23), for the Big Pig study, 109 Large White/Landrace cross-bred pigs were infected with the North American prototype PRRSV American Type Culture Collection (ATCC) VR-2332, and followed for 202 d. Specifically, on day 0, the pigs (n = 109) were IM inoculated with 1 mL of PRRSV (VR-2332 at 1 × 104.0 TCID50) diluted in minimum essential medium (MEM). Control pigs (n = 56) were inoculated with PRRSV-negative MEM by the same method. Blood samples were collected from the pigs on day 0 before inoculation, at 7 and 14 dpi, and every 14 d afterward up to 202 dpi. All data from the Big Pig studies have now been archived in a new database, the PRRS Host Genetics Consortium database (Fritz et al., manuscript in preparation). This database provides a resource to probe for potential correlations between data from Big Pig samples and levels of immune parameters (e.g., serum cytokines and antibody levels) as discussed here, or with genetic background (SLA class I and II genes), which will be presented separately.
Viral RNA quantitation using reverse-transcriptase polymerase chain reaction (RT-PCR)
Quantitative, real-time RT-PCR (qPCR) was used to determine viral levels as previously described (34), using a commercially available kit (Tetracore Inc., Rockville, MD). Quantification of sample viral levels was calculated and expressed as the number of RNA copies per milliliter for fluid samples, or the number of RNA copies per gram for solid tissue samples. Virus levels were quantitated for inguinal lymph nodes (IngLN), submaxillary LN (SubLN), and tonsils. More tissues were collected for gene expression work, including lung and tracheobronchial LN (TBLN) (22,23).
Gene expression in tissues
Real-time PCR for immune marker gene detection was performed as previously described (8,29). In the Big Pig study, 5–10 pigs were removed from the study at biweekly time points (22,23). Tissues, including tonsils and LN, were collected and stored in RNAlater at −80°C until use. Our focus was first directed to the TBLN, since these LN drain the lung, the tissues where PRRSV replicates; independent studies have indicated that tonsils would have lower gene expression than TBLN (Lunney et al., unpublished). Gene expression data were determined for RNA isolated from TBLN samples from individual pigs, and compared to a randomly selected set of age-matched control Big Pig TBLN samples collected over the course of the study. Each cDNA sample was analyzed for expression of 23 immune-related genes involved in both innate and acquired immunity, and one housekeeping gene (RPL32). These included a range of immune markers: for Th-1 cell activation (IFNG, IL12B, IL15, IRF1, CT11A, TNF, NOS2A, and CXCL10), Th-2 cell activation (IL5 and IL13), innate immunity (IFNA, CSF2, IL1B, IL6, and IL8), and for T-regulatory (T-reg) (IL10), apoptosis (CASP1, CASP3, CASP8, TAP2, and TGM3), and chemokine (CCL2 and CCL5) signaling. Relative quantification of target gene expression was evaluated using cycle threshold (Ct) values, and recorded with the Applied Biosystems PRISM 7700 Sequence Detection System (Carlsbad, CA). Each individual's data were calculated, then averaged for animals at the same time point, and the group's results were compared to the values for averages of the control pigs using the JMP statistical package (SAS Institute,, Cary, NC), as previously described (8,29). Least squares-means analyses were used to statistically evaluate group means for immune gene analyses. For individual comparisons, each gene's Ct value was compared to the control average for that gene. When gene expression data are presented, the official gene name is noted (e.g., IFNG, not IFN-γ is used for the protein name).
Measurement of cytokines in serum
The cytokines IL-1β, IL-8, and IFN-γ were quantified in sequential serum samples, collected at 0 and 7 dpi and at 14-day intervals thereafter, from 72, 44, and 43 pigs, respectively, of the 109 PRRSV-inoculated animals. Levels of IL-1β and IL-8 were evaluated in samples collected through 42 dpi, and IFN-γ in samples collected through 84 dpi, to cover the peak times of serum expression. The serum samples were assayed with commercial ELISA kits (IL-1β and IL-8: R&D Systems, Minneapolis, MN; IFN-γ: Biosource™ Invitrogen, Camarillo, CA) following the protocols provided by the manufacturers. The substrate, SureBlue™ (KPL, Inc., Gaithersburg, MD), was added and plates were read at 650 nm until the highest standard reached an optical density (OD) of ∼2.0 with no acid added. For all ELISA data, the OD of the medium controls was averaged and subtracted from the OD of each sample, and then the lowest readable concentration was determined from the standard curve. If the sample ELISA OD was below the lowest level, minimum detectable levels were assigned by dividing the lowest readable standard concentration by 2, and then multiplying that number by the dilution factor.
Statistical analysis
Data were analyzed using the PROC-GLM and least squares-means methods of SAS software (SAS Institute Inc., Cary, NC). Initially the data were analyzed with the following statistical model to determine if IL-8, IL-1β, and IFN-γ had an effect on virus levels:
where y is virus level, μ is the general mean, a is the effect of IL-8, b is the effect of IL-1β, c is the effect of IFN-γ, d is the effect of day, a × d is the effect of the interaction of the day and IL-8, and c × d is the effect of the interaction of the day and IFN-γ.
Interest in persistence (a pig was classified as persistent if at euthanasia virus was not cleared from serum or tissues) of viral infection led to an individual analysis of whether or not each of the following—virus level, IL-8 level, IL-1β level, or IFN-γ level—had an effect on whether or not a pig was classified as a persistent carrier of the PRRS virus or not. We used this model:
where y is virus level, IL-8, IL-1β, or IFN-γ level, μ is some mean, a is the effect of the persistence classification (persistent or non-persistent), b is the effect of day, and a × b is the effect of the interaction of the day and the persistence classification.
Results
Assignment of persistence status and viral clearance
Based on viral RNA levels in serum and tissues, the pigs were assigned to either non-persistent (NP) or persistent (P) groups. The NP pigs were identified as those pigs that were negative for PRRSV RNA in their serum by 28 dpi, and were negative for PRRSV in tissues (tonsils, IngLN, and SubLN) when harvested starting at 112 dpi (Table 1). The P pigs were positive for viral RNA in serum at least until 28 dpi (average: 68 dpi), and positive for viral RNA in tissues for up to 202 dpi. The results in Table 1 show that 39 pigs could be clearly assigned to the P or NP group, all of which had TBLN processed for gene expression studies. Of these pigs, 15 met the criteria for the NP group, and 24 could be placed in the P group. Analyses of serum viral levels of NP and P pigs illustrated an important difference between the P versus NP groups of pigs (Fig. 1). Up to 14 dpi there was no statistically significant difference in viral RNA levels between the NP and P groups; however, a statistically significant (p < 0.05) difference was apparent at both 28 and 42 dpi.
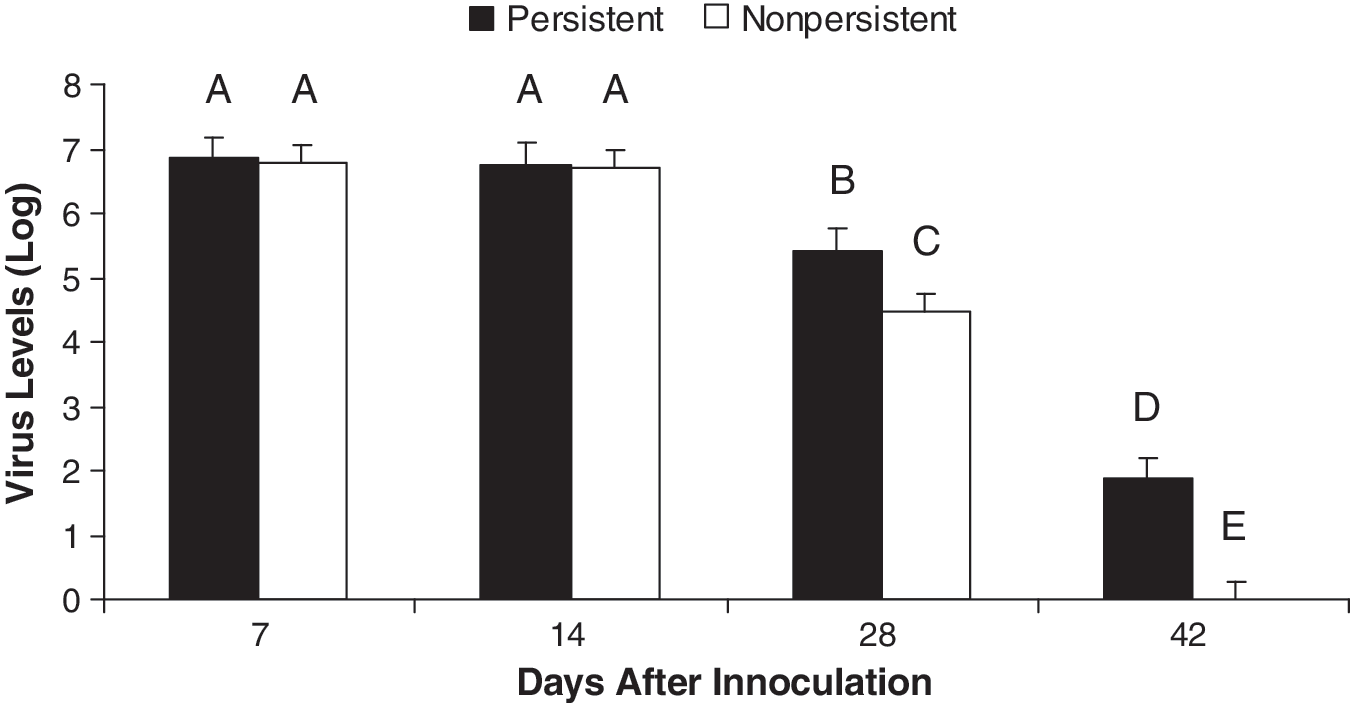
Comparison of viral clearance patterns of pigs with persistent (P) versus non-persistent (NP) PRRSV infection. Virus RNA level estimations for dpi 7, 14, 28, and 42 were compared. Statistically significantly different (p < 0.05) levels are designated with different letters (A–E).
The persistence status was determined based on viral RNA qPCR analyses of serum and tissues, tonsils, inguinal lymph nodes (IngLN), and submaxillary LN (SubLN) (Molina et al., 2008a [22]). Tissue viral loads were noted as positive if one, two, or three of the tonsils, IngLN, or SubLN tested PRRSV-positive by qPCR analysis.
NP = non-persistent pigs, or those with cleared infection. These pigs were defined as negative for evidence of PRRSV by viral RNA qPCR analysis of serum at 28 dpi or later, and tissues at the time of euthanasia (between 112 and 202 dpi).
P = persistent pigs, or those with proven persistent infection. These pigs were defined as positive for evidence of PRRSV by viral RNA qPCR analysis of at least one, if not all three, lymphoid tissues and/or tonsils at the time of euthanasia (between 112 and 202 dpi). They were serum virus qPCR-positive at least until 28 dpi (average: 68 dpi), and several were serum virus qPCR-positive until 98–126 dpi.
Immune-related gene expression in tracheobronchial lymph nodes
The original goal of the Big Pig persistence studies was to determine whether gene expression for immune markers in local tissues could identify factors associated with persistence. To this end, RNA prepared from infected TBLN from pigs removed at different time points were tested for expression of a panel of immune markers (Fig. 2A). Samples from each time point were tested, Ct values were averaged, and the gene expression data for each time point were averaged and compared to the average Ct for control tissues. The results showed upregulation of expression of IFNG-associated Th-1 markers, with the highest upregulation seen from 42 to 70 dpi. Importantly, there was no significant upregulation of most innate markers (IFNA, IL1B, and IL8), since an impaired innate response is considered a key factor in the poor response to PRRSV infection. In addition, there was significant upregulation of the T-reg marker IL10, appearing as early as 14 dpi, and lasting until 84 dpi. Several apoptosis markers were upregulated, peaking at 42 dpi. By 112 dpi, expression levels of most immune markers were lower than the averaged control tissues. The results in Fig. 2B show the same gene expression data reported on TBLN RNA, but from individual pigs in the NP and P groups. The data indicated no or low differential gene expression for most genes, and no apparent difference between individual NP and P pigs in TBLN immune gene expression.
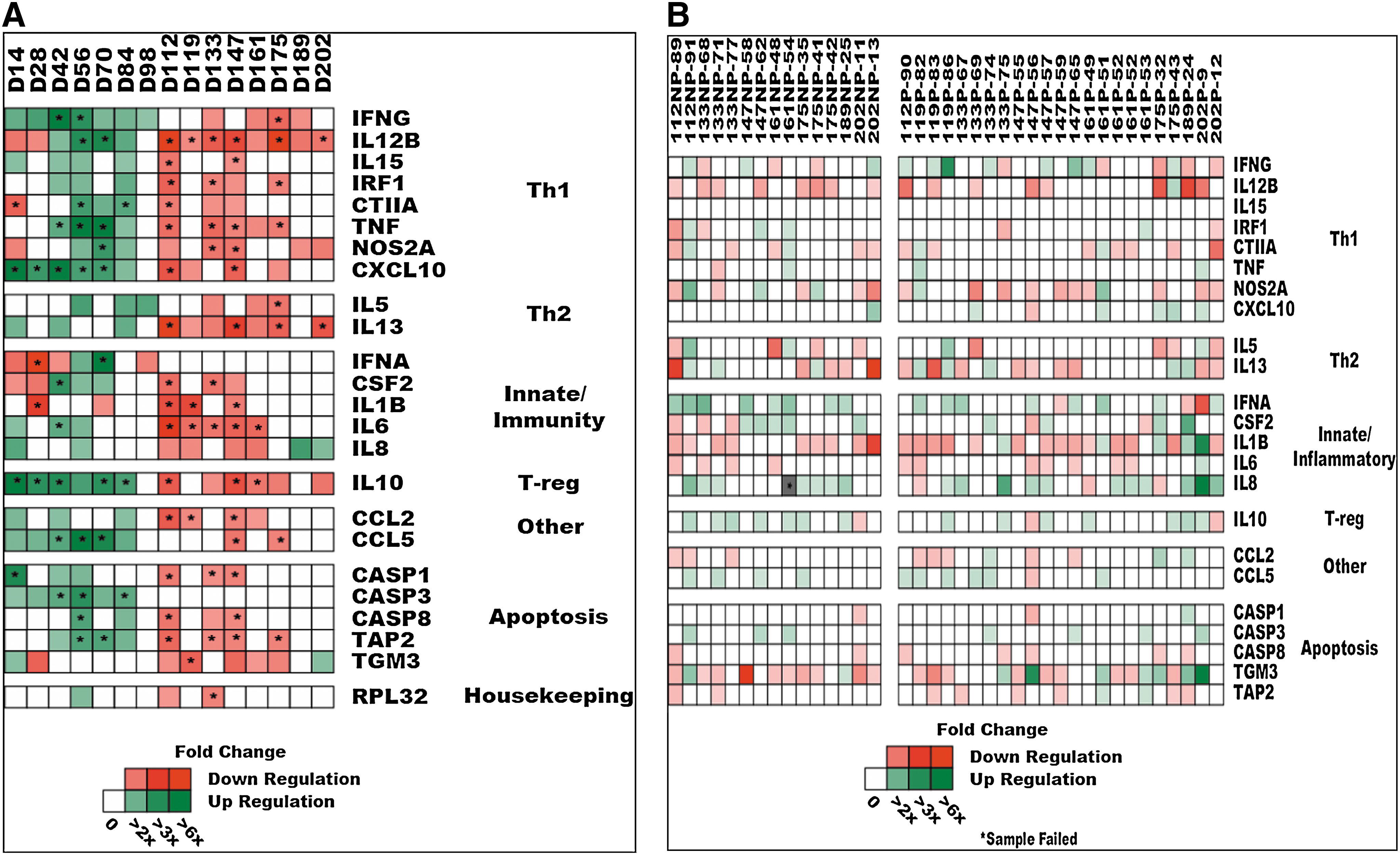
Gene expression of immune markers from RNA prepared from tracheobronchial lymph node (TBLN) samples from PRRSV-infected pigs collected during the Big Pig studies. (
Serum cytokine protein concentration in serum
Unlike tissues, which were collected only once, serum samples were collected biweekly until termination for the Big Pig study. Therefore we could analyze early cytokine protein events in each pig. Data from 44 PRRSV-infected pigs were analyzed for effects of serum cytokine levels on PRRSV RNA levels using the PROC-GLM function of SAS with the full model. For the NP/P comparisons complete serum sample sets were available for all but 1 NP and 3 P pigs. The model was significant (p < 0.0001) with an R2 = 0.84 (Table 2). This indicates that variation in all three of the cytokines tested (IL-8, IL-1β, and IFN-γ) had a significant correlation with virus level in all PRRSV-infected pigs, accounting for ∼84% of the variation observed.
p Values for model and effects of cytokines and/or day post-infection (Day) on virus levels for all animals, or separate analyses of persistent (P) and non-persistent (NP) animals. All p values <0.05 are in bold type.
Number of infected pigs for which sera from 0, 7, 14, 28, and 42 dpi were evaluated for IL-8 and IL-1β levels by ELISA. For IFN-γ, sera from additional time points (56, 70, and 84 dpi) were evaluated with 1 fewer NP pig.
Denotes interaction between cytokine and day.
Statistical analyses indicated that innate cytokines (IL-1β and IL-8) were increased earliest, followed by IFN-γ (Fig. 3A, C, and E). The main effects of IL-8, IL-1β, and day on viral levels were significant (p < 0.01), while the main effect of IFN-γ was not significant (p > 0.10). The interaction of day with IL-8 was significant (p < 0.05), while the interaction of day with IFN-β was suggestive of an effect on viral levels (p < 0.10) (Table 2).
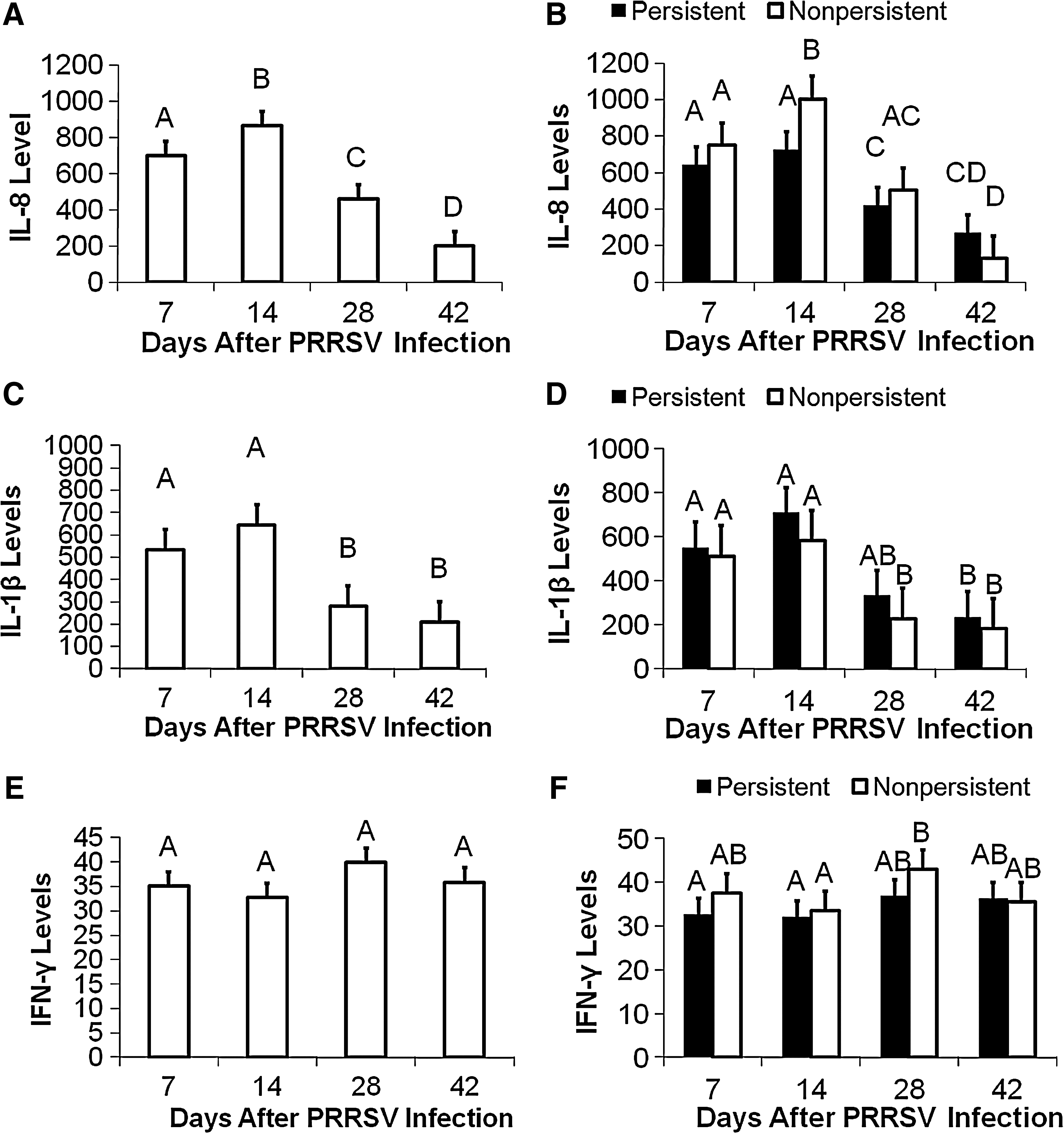
Comparison of serum cytokine levels after PRRSV infection. (
The infected pigs exhibited high serum IL-1β until 14 dpi. When IL-8 was examined at 14 dpi, NP pigs had higher concentrations of IL-8 than P pigs (Fig. 3B). There appeared to be no changes in serum IL-10 associated with infection (data not shown). Overall, pigs with non-persistent PRRSV infections appeared to have earlier and higher serum IL-8 levels than pigs with persistent infections.
Effect of persistence
Data from the PRRSV-infected pigs were run through the PROC-GLM function of SAS using the single trait model. The model was run using virus, and serum IL-8, IL-1β, and IFN-γ levels. The single trait model was significant for virus. The main effects of persistence and day, as well as the interaction of persistence with day, were all significant for virus level (p < 0.01; Fig. 1). The virus level significantly decreased on dpi 28, and further on dpi 42. Comparisons revealed significant differences between P and NP pigs on dpi 28 and 42 (Fig. 1).
Serum IL-8 levels were significantly affected by dpi (p < 0.01; Fig. 3A). Interleukin-8 levels peaked on dpi 14, and then significantly decreased to 28 dpi, and continued to decrease to 42 dpi. In contrast, the main effect of persistence and the interaction of persistence with day were not significant for serum IL-8 (p > 0.10; Fig. 3B). Days post-infection also significantly affected IL-1β level (p < 0.01; Fig. 3C), while the main effect of persistence and the interaction of persistence with day were not significant (p > 0.10; Fig. 3D). Interleukin-1β levels were similar for 7 and 14 dpi, and were significantly higher than both 28 and 42 dpi. The number of days post-infection, persistence, and the interaction between persistence and day did not significantly affect IFN-γ levels (p > 0.10; Fig. 3E and F). Serum IFN-γ levels were moderately increased at 28 dpi, and were statistically significantly different only in the NP pigs at 28 dpi versus the 14 dpi values; no differences were detected at 42 dpi (Fig. 3F).
Using the full model, data from P pigs were run through the PROC-GLM function of SAS. Variations in serum IL-8, IL-1β, and IFN-γ, along with the interactions between IL-8 and day, and IFN-γ and day, accounted for a significant amount of variation in viral levels (p < 0.0001; R2 = 0.75; Table 2). The main effect of day was significant (p < 0.01), while the main effect of IL-1β was only suggestive (p < 0.10), and the main effects of serum IL-8 and IFN-γ on viral levels were not significant for P pigs. The interaction of day with IL-8 was significant (p < 0.05), while the interaction of day with IFN-γ was not significant (p > 0.10).
When sera from NP pigs were analyzed, variations in serum IL-8, IL-1β, and IFN-γ, along with the interactions between IL-8 and day, and IFN-γ and day, accounted for a significant effect on viral levels (p < 0.0001; R2 = 0.89; Table 2). The main effect of day was significant (p < 0.01), while the main effect of IL-1β was suggestive for NP pigs (p < 0.10). In contrast, the main effects of IL-8 and IFN-γ were not significant (p > 0.10). The interaction of day with IL-8 was not significant (p > 0.10), and the interaction of day with IFN-γ was suggestive for correlation with viral levels in NP pig serum (p < 0.10) (Table 2).
Discussion
Numerous studies have affirmed PRRSV persistence in tissues for extended time periods (1,2,14,27,35,36). The Big Pig data have confirmed that persistence occurs until 147 dpi, based on viral RT-PCR (22,23). The underlying mechanism supporting or preventing PRRSV persistence in tissues has not yet been identified. In this article we have presented data that support the hypothesis that serum cytokine levels detected early after PRRSV infection may help determine whether the virus will persist in blood and tissues. Our original hypothesis, that mucosal respiratory tissue (TBLN) immune gene expression level at the day of euthanasia would be correlated with persistence, proved wrong and revealed no differential reactivity of RNA from P versus NP pigs.
Sera collected early after PRRSV infection did show differential reactivity of P versus NP pigs. Statistical analyses indicated that innate cytokines were increased earliest. Levels of serum IL-8 and IL-1β at 14 dpi, followed by Th-1 and IFN-γ at 28 dpi, indicated that the P pigs had a higher concentration of IL-1β and lower serum IL-8 than the NP animals. Thus serum immune cytokine protein expression early after infection was associated with determining which pigs would have persistent PRRSV infections.
The results of running the single trait model on virus level showed that persistence, day, and the interaction between day and persistence were all significant. This indicated that there is indeed a statistically significant difference in virus levels between pigs classified as P and NP. The factor of day being significant indicates that virus levels varied throughout the period of examination, as would be expected, that virus levels increase after infection and subsequently decrease as the pigs mounted an immune response. The finding that the interaction of day with persistence is significant indicates that there is a difference between P and NP pigs at different time points (Fig. 1). Thus, P pigs do not clear the virus completely or as effectively as NP pigs, indicating differences in their immune responses.
There was a day effect for both IL-8 and IL-1β, which means that these levels changed over time (Fig. 3). However, there was no persistence or effects of the interaction of day with persistence for single cytokines. However, serum cytokine protein levels indicated that the NP pigs had earlier and higher serum levels of the innate cytokine IL-8 than the P pigs. This immune cytokine trend was correlated, with NP pigs having lower serum and tissue viral loads, indicating that the NP immune response was more effective than that of P pigs, and possibly prevented the NP pigs from becoming persistently infected with PRRSV. These data confirm the work done by Petry et al. (26), who found that pigs with a higher pre-infection serum IL-8 level were more resistant to PRRSV infection. Overall, in our study pigs with non-persistent PRRSV infections appeared to have earlier and higher IL-8 levels than pigs with persistent infections. However, more information is needed to determine if this is actually the case, or if this is the result of a small dataset.
While individual serum cytokine levels did not appear to have an effect on persistence, they did have a significant effect on virus level, whether directly or indirectly. When the full model was run using all PRRSV-infected pigs, the main effects of serum IL-8, IL-1β, and day were all significant for correlation with viral levels, while the effects of the interactions of IL-8 with day and IFN-γ with day were only suggestive (Table 2). Overall, 84% of the observed variation in virus levels was significantly correlated with levels of not one, but all three, of the serum cytokines tested (IL-8, IL-1β, and IFN-γ). This indicates that since virus level was significant when run through the single trait model based on persistence, the cytokines together could possibly have an effect on persistence, or be used to predict persistence. Indeed, numerous cytokines have been reported to influence responses to PRRSV infection (4,7,9,10,11,21,30 –33). In fact, it is likely that cytokines other than IL-1β, IL-8, and IFN-γ influence viral persistence status (e.g., innate IFN-α and Th-1-associated IL-12).
Footnotes
Acknowledgments
This work was funded by the U.S.D.A. Agricultural Research Service, and by grants from the U.S.D.A. CSREES #2004-35604-14580 PRRS CAP1, and National Pork Board grant #07-233. The authors would like to thank Dr. M. de Jong, Wageningen, The Netherlands, for his suggestions on data analyses.
Author Disclosure Statement
No competing financial interests exist.