
Abstract
This report describes the identification of a novel linear B-cell epitope at the N-terminus of the avian infectious bronchitis virus (IBV) nucleocapsid (N) protein. A monoclonal antibody designated 2D2 was prepared against the IBV N protein, and a series of 23 partially overlapping, tagged fragments of the IBV N gene were expressed. These peptides and five heterogeneous IBV strains were used to identify the epitope recognized by 2D2 using enzyme-linked immunosorbent assay (ELISA) and Western blotting analysis. The linear motif 11DSPAPIIKLGG21, located at the N-terminus of the N protein, was identified as the epitope bound by 2D2. ELISA and Western blotting also showed that this epitope was recognized by IBV-positive chicken serum. We propose that this motif is a linear B-cell epitope of N protein, and is the core sequence for reactivity, as demonstrated by binding of 2D2 to the truncated peptides and different IBV strains. Alignment and comparison of the 2D2-defined epitope with other coronavirus sequences showed that the epitope is conserved among avian coronaviruses. The identified epitope might be useful for clinical applications and as a tool for further study of the structure and function of the IBV N protein.
Introduction
The N protein, which is the most abundant viral structural protein, is associated with the viral RNA genome (29). Although its primary function is formation of the viral ribonucleoprotein complex, several studies indicate that it is multifunctional. Its intracellular localization suggests that it is a likely component of the coronavirus replication and transcription complex. Although one study found that N is not involved in immune protection (12), it can induce high titers of cross-reactive antibodies and induce cell-mediated immunity that protects chickens from acute infection (12,38,44), so it is used as a target protein in designing infectious bronchitis (IB) vaccines (45). The N protein is highly conserved, showing 91.0–96.5% similarity among different IBV strains (47). It is abundantly expressed during infection. These features make the IBV N protein a frequent target of diagnostic applications (9,33).
To understand antigenicity and virus-antibody interactions at the molecular level for vaccine design and other clinical applications, the location of viral protein epitopes must be identified. However, few B-cell epitopes associated with the above functions have been mapped in the IBV N protein. Seah et al. (37) expressed the full-length N protein as 12 overlapping fragments in Escherichia coli (E. coli), and found that seven contained B-cell epitopes, with the most antigenic regions between amino acids 175–241, 310–409, and 360–409. Ignjatovic and Sapats (13) recently identified a conformation-independent epitope (amino acid residues 72–86) that induced a delayed-type hypersensitivity response, but in spite of being antigenic, it did not confer protection against IBV. All epitopes identified in IBV N protein have not been finely mapped, and the core sequences of the identified and other epitopes need to be investigated.
Monoclonal antibodies (mAbs) are widely used as powerful tools for identifying linear epitopes, or for mimicking epitopes of a variety of infectious agents (8,18). In this study, we prepared a mAb against N protein from the IBV strain CK/CH/LDL/97I, and used it to screen for linear B-cell epitopes. The results provided important information for vaccine design, the development of specific serological diagnostics of IBV infection, and for better understanding of the antigenic structure of N protein.
Materials and Methods
Virus propagation in specific pathogen-free embryonated eggs
IBV strain CK/CH/LDL/97I (25) was used for N gene cloning and for preparation and identification of the mAb. This IBV strain is a novel IBV strain type isolated in 1997 from the swollen proventricular tissues of infected chickens in Dalian, Liaoning province, China. It has been associated with some IB outbreaks in recent years in China (25). For Western blotting, five heterogeneous IBV strains composed of four field strains and one commercially available vaccine strain were used to represent different IBV types (Table 1) (24,26). The five heterogeneous IBV strains were used to determine if the identified epitope was conserved among the different IBV serotypes circulating in China. All IBV strains were propagated once in 9- to 11-day-old specific pathogen-free (SPF) embryonated chicken eggs, and the presence of viral particles in the allantoic fluid of inoculated eggs was confirmed with a negative contrast electron microscope as previously described (23).
Country (province) where the viruses were isolated.
Year when the viruses were isolated.
Fertile white Leghorn SPF embryonated chicken eggs were obtained from the Laboratory Animal Center, Harbin Veterinary Research Institute, Chinese Academy of Agricultural Sciences, Harbin, China.
Generation of mAbs
Two 8-week-old BALB/c female mice were immunized subcutaneously with condensed IBV CK/CH/LDL/97I virus-infected allantoic fluids (48) mixed with Freund's complete adjuvant, followed by two booster immunizations. Protocols used for the preparation of mAbs and ascetic fluids were as previously described (36,46,48). All hybridomas were cloned by at least three rounds of limiting dilution. Primary screening of hybridomas was by enzyme-linked immunosorbent assay (ELISA) using IBV CK/CH/LDL/97I virus particles as coating antigen. mAb class and subclass were determined using an SBA Clonotyping System/HRP kit (Southern Biotechnology Associates, Birmingham, AL).
Construction of expression vectors and expression of fusion proteins
The N gene and all truncated N gene fragments (Fig. 1) were amplified from the IBV CK/CH/LDL/97I strain as previously described (25). Genes and gene fragments were cloned into the pGEX-6P-1 vector (Pharmacia, Brussels, Belgium) using EcoR I and Hind III sites that were introduced into the sense and negative-sense primers, respectively. A collection of expression clones corresponding to the entire N gene or truncated fragments was constructed and sequenced. Sequences and locations of primers are shown in Table 2.
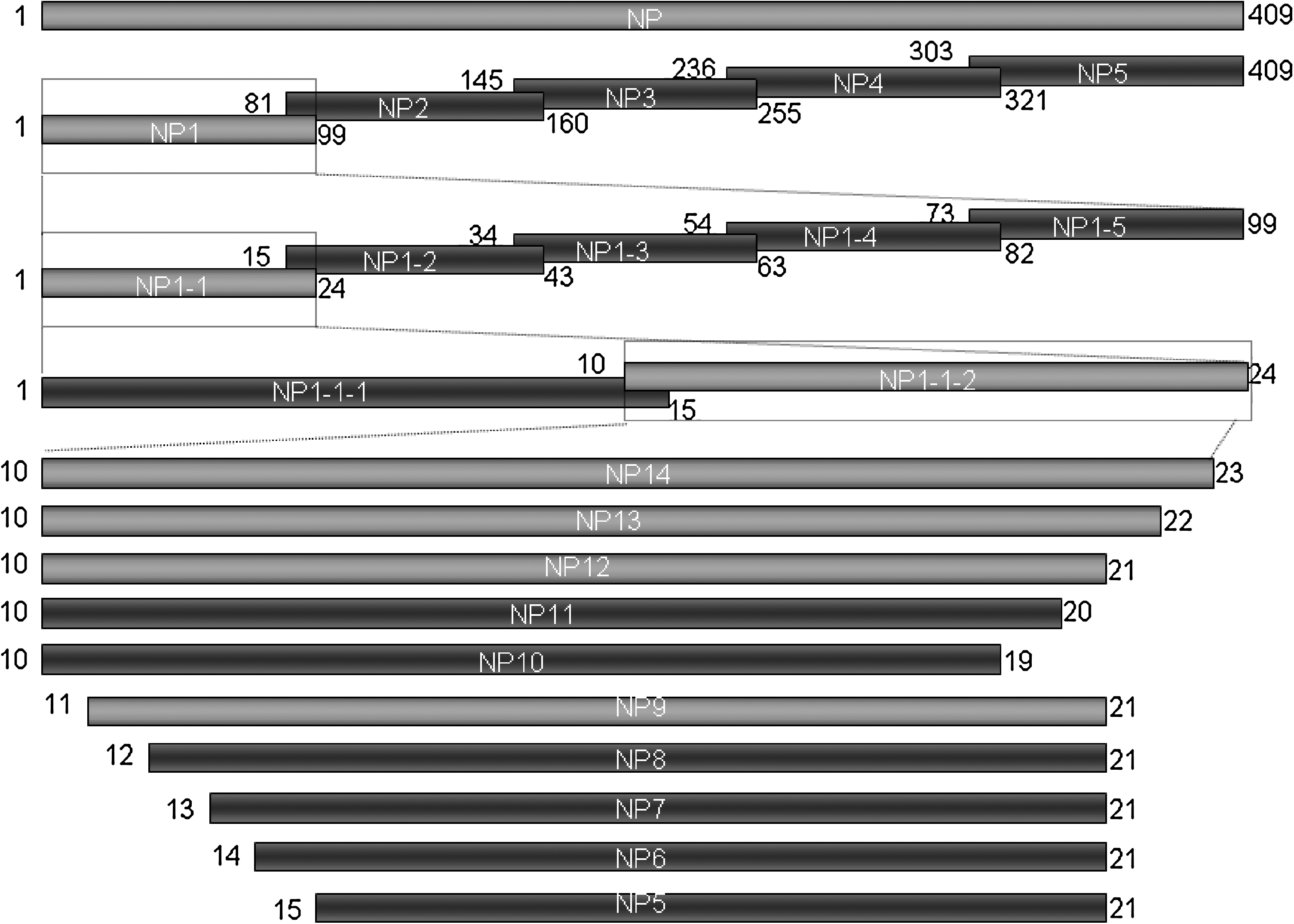
Schematic diagram showing the truncated fragments from the IBV CK/CH/LDL/97I N protein and their relative positions. Letters represent the amino acid positions. The names of the peptides are shown in Table 2. Bars represent truncated N peptides. Peptides that were negative by Western blotting with mAb 2D2 are in dark grey, and peptides that were positive are in light grey.
Underlined letters indicate additional restriction enzyme sites (Bam HI and Sal I) introduced into each primer. The boxed ATGs were the start codon of the IBV CK/CH/LHLJ/04V M gene, and the TTAs were the stop codon of the N gene or additions introduced into the negative-sense primers of the N fragments.
The nucleotide positions correspond to those in the sequence of the IBV CK/CH/LDL/97I N gene (GenBank accession no. EF602445).
Each recombinant expression construct was transformed into E. coli BL21 (DE3) (Novagen, Gibbstown, NJ). A series of fusion proteins with the expected molecular weights were induced by IPTG as previously described (48), and stained with Coomassie blue after SDS-PAGE, also as previously described (48). For preparation of purified proteins, inclusion body proteins were separated by SDS-PAGE, the proteins of interest were excised, and gel slices were crushed and added to an appropriate volume of sterilized PBS. The extracted proteins were used for Western blotting and ELISA.
SDS-PAGE and Western blotting
The specificity and reactivity of the mAbs were determined by Western blotting using recombinant N protein, IBV strain CK/CH/LDL/97I, and the five heterogeneous strains (Table 1). Purified recombinant N protein and condensed IBV infectious allantoic fluids were separated by denaturing SDS-PAGE. For Western blotting, the proteins were transferred onto nitrocellulose membranes, and detected as previously described (48). Purified GST protein and the sterilized allantoic fluids of SPF eggs were used as negative antigen controls. SPF chicken sera were used as a negative serum control.
Indirect ELISA
The specificity of the mAb was determined by indirect ELISA using IBV CK/CH/LDL/97I virus particles as the coating antigen as previously described (48). A commercial total antibody ELISA (IDEXX Corp., Westbrook, ME) was used with some modifications to the manufacturer's instructions, specially, the tested mAb replaced the serum samples, and HRP-conjugated sheep anti-mouse IgG was used to replace the goat anti-chicken IgG. Antibodies against IBV were the positive control, and SPF sera were the negative control. All assays were performed in triplicate. The reactivity of the mAbs with different truncated recombinant N proteins was determined by ELISA, as previously described (48). All assays were performed in triplicate and the serum-to-positive ratios (S/P ratios) were calculated as previously described (39,40,41).
Comparison of the epitope-containing sequence with other coronaviruses
The sequences containing the epitope were assembled, aligned, and compared with coronavirus strains using the MEGALIGN program in DNAStar. The sequences of the reference coronaviruses were from the GenBank database, with the GenBank accession number in Fig. 2.

Alignment of the epitope motif in 20 group 3 coronaviruses. Epitope sequences are underlined and amino acid residues in the epitope region that are shared by different coronaviruses are in grey. Deleted amino acid residues are represented by dashes (TCoV, turkey coronavirus). GenBank accession numbers are in parentheses.
Results
Expression of recombinant proteins
The strategy for expressing IBV N and truncated N fragments is shown in Fig. 1. The entire N gene and truncated fragments were expressed as GST fusion proteins in E. coli BL21 (DE3). All the proteins were successfully expressed and examined by SDS-PAGE of cell lysates after induction with IPTG. The recombinant N protein reacted with a chicken antibody against IBV by both ELISA (Fig. 3A) and Western blotting (Fig. 3B).
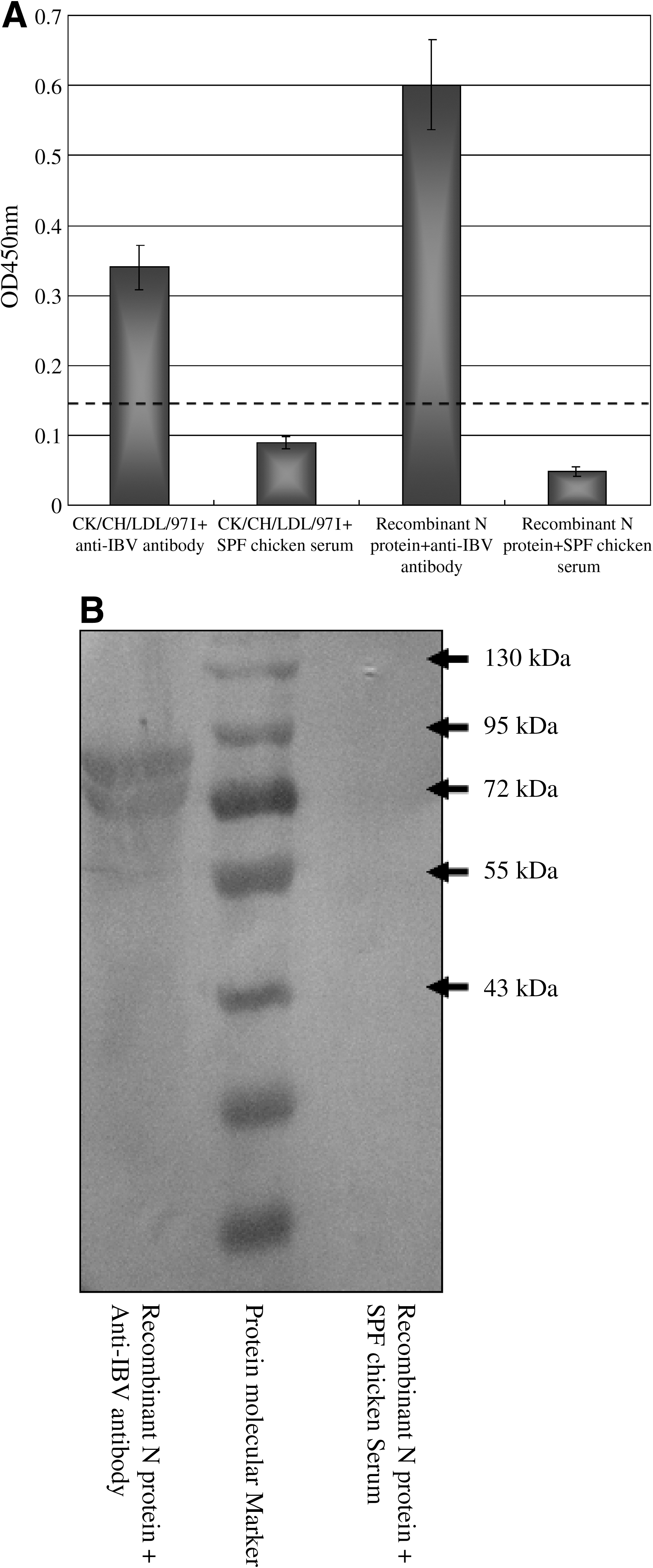
Reactivity of recombinant N protein with chicken antibody against IBV by ELISA (
Production of IBV N mAb 2D2
The hybridoma 2D2 was found to secrete mAbs against the IBV CK/CH/LHLJ/04V N protein. The produced mAbs recognized both recombinant N protein and native IBV CK/CH/LDL/97I antigen by Western blotting (Fig. 4A). The reactivity and specificity of mAb 2D2 were confirmed using both a commercial ELISA and an ELISA in which whole IBV CK/CH/LHLJ/04V virus particles were used as the coating antigen (Fig. 4B). The mAb 2D2, which was determined to be IgG1 (κ), was used for additional studies.

Reactivity of mAb 2D2 with IBV strain CK/CH/LDL/97I and recombinant N protein by Western blotting (
Localization of the mAb 2D2-defined epitope
The GST-fusion proteins were used for fine mapping of the epitope of the IBV CK/CH/LDL/97I antigen that is recognized by 2D2 (Fig. 1). Western blotting showed that the minimal recognition sequence appeared to be 10SDSPAPIIKLGG21, because deletion of S10 or G21 destroyed the binding of mAb 2D2 to the GST-fusion peptides (Fig. 5A). Therefore, by Western blotting the linear epitope recognized in the N protein of IBV strain CK/CH/LHLJ/04V by 2D2 was 10SDSPAPIIKLGG21. However, the minimal sequence identified by ELISA was 10SDSPAPIIKLGGP22, using the truncated peptides as coating antigens (Fig. 5B). Thus the linear B-cell epitope in the N protein of IBV recognized by mAb 2D2 appeared to be localized within 10SDSPAPIIKLGGP21.

Fine localization of the mAb 2D2 epitope. Reactivity of mAb 2D2 with truncated recombinant N proteins by Western blotting (
Localization of 2D2-bound epitope by cross-reactivity of heterogeneous IBV strains
We examined the cross-reactivity of mAb 2D2 with five IBV strains, representing five different serotypes. As illustrated in Fig. 6, all heterogeneous IBV strains cross-reacted with mAb 2D2, with Western blotting of infected allantoic fluids showing a pronounced band of the expected molecular weight except CK/CH/LHLJ/04V, which had an additional small band. The sequence consistently shared by the CK/CH/LDL/97I strain and the five heterogeneous IBV strains was 11DSPAPIIKLGG21, as indicated in Fig. 6. Thus the linear B-cell epitope in the IBV N protein recognized by mAb 2D2 appeared to be precisely localized within 11DSPAPIIKLGG21, which is located at the N-terminus of the IBV N protein.

Reactivity of mAb 2D2 with five heterogenous IBV strains by Western blotting, and alignment of the epitope motif with the five strains. Epitope sequences are underlined and amino acid residues in the epitope region that are shared by different coronaviruses are in grey. Deleted amino acid residues are represented by dashes. The GenBank accession numbers are as shown in Table 1.
Reactivity of the identified epitope with anti-IBV antibody
The peptide 11DSPAPIIKLGG21, which corresponded to the epitope defined for mAb 2D2, was used as the coating antigen in ELISA assays, which demonstrated that this peptide was also recognized by a chicken anti-IBV antibody (Fig. 7A). The peptide showed similar reactivity by Western blotting analysis (Fig. 7B), further confirming the reactivity of this epitope.

Reactivity of the identified epitope (11DSPAPIIKLGG21) with antibodies against IBV. The peptide corresponding to the mAb 2D2-defined epitope was the coating antigen for ELISA, and NP12 and NP8 were used as the positive and negative controls, respectively. Anti-IBV chicken antibody was the primary antibody, and SPF chicken sera were the negative controls (
Alignment of the 2D2-defined sequence with other coronaviruses
Only a limited number of heterogeneous IBV strains could be used in 2D2 cross-reactivity tests, and no other group 3 coronavirus strains were available in our laboratory. Therefore, we aligned and compared the regions containing the sequence DSPAPIIKLGG from 11 heterogeneous strains, representing 11 additional IBV serotypes from different global regions, and 9 other group 3 coronaviruses that infect three different hosts. The results showed that 9 of the 11 residues in the mAb 2D2-defined sequence were highly conserved among chicken, turkey, pheasant, and pigeon group 3 coronaviruses (Fig. 2). This indicated that the 11DSPAPIIKLGG21 sequence represents a relatively conserved epitope on the avian coronaviruses' N proteins. Sequences from other coronaviruses from different groups or subgroups were used for comparison, but no sequences corresponding to the 2D2-defined epitope were found (data not shown).
Discussion
The three major coronavirus structural proteins are the S and M glycoproteins, and the nucleocapsid N protein (29). These are the main targets for vaccine design, and for the development of immunity-based prophylactic, therapeutic, and diagnostic techniques for controlling coronavirus. To date, identification of B-cell antigenic epitopes for the IBV S, N, and M proteins has been limited, although some of the epitopes induce the production of virus-neutralizing antibodies, and provide protection against virulent viruses (1,12,13,15,17,19,20,21,30,34,37,38,48). In this study, we discovered a novel linear B-cell epitope at the N-terminus of the IBV N protein.
In this study, we generated the mAb 2D2, and found that it reacted specifically against native IBV CK/CH/LDL/97I particles, as well as against recombinant N protein. However, an additional small band was also observed by Western blotting analysis using recombinant N protein to react with both mAb 2D2 and anti-IBV antibody, which may be cleaved by bacterial cellular proteases (4,7,31,32,42). To study the specificity in more detail and to finely map the epitope bound by mAb 2D2, a series of 23 partially overlapping fragments covering the N gene of IBV CK/CH/LDL/97I were expressed with a GST tag, and used to screen for the minimal epitope. Five heterogenous IBVs were also used to identify the minimal epitope sequence by Western blotting and ELISA. ELISA using recombinant proteins as coating antigens showed that the core sequence was 10SDSPAPIIKLGGP22, while Western blotting indicated that the minimal sequence was 10SDSPAPIIKLGG21. This discrepancy may be attributed to the sensitivity of the methods (27,28). Interestingly, we found that mAb 2D2 reacted with five heterogenous IBVs by Western blotting, although the proteins recognized by the mAb 2D2 in most of the viruses produced very faint bands. The bound sequence shared by CK/CH/LDL/97I and the five heterogenous IBV strains was 11DSPAPIIKLGG21, which was one amino acid shorter than the sequence recognized using the truncated recombinant N proteins as antigens. This may be because the recombinant N protein expressed in E. coli had lower antigenicity compared to the native protein in the IBV particles. In addition, the N-terminal GST moiety might interfere with binding of mAb 2D2 to the recombinant protein. Compared to our previous reports (43,48), the positive ELISA test values using recombinant proteins as coating antigens were slightly lower (not more than 0.5) (Fig. 5B). Taken together, we conclude that the core sequence of the epitope defined by mAb 2D2 in the IBV N protein is 11DSPAPIIKLGG21.
To date, the identified linear and conformation-independent B-cell epitope regions of IBVs have been at amino acid positions 72–86, 175–241, 310–409, and 360–409 (13,37). In this study, mAb 2D2 identified a novel linear B-cell epitope at positions 11–21. To our knowledge, this is the first finely-mapped B-cell epitope in the IBV N protein. Furthermore, we found that this epitope is highly conserved among avian coronaviruses. The coronaviruses isolated from different avian species (chicken: IBV; turkey: TCoV; pheasant: PhCo; and pigeon: pigeon coronavirus) have been proposed to be distinct species (6). In different avian species, the S gene of coronaviruses does not have more than 57% identity. However, the N genes show higher identity among coronaviruses of avian species (5,10,16), and little has been known about antigenic epitopes within the structural proteins of avian coronaviruses, except for the IBVs. The conserved epitope shared by the avian coronaviruses' N proteins is important, not only because the avian coronaviruses have economic impact on the poultry industry, but also because they also show antigenic and biological similarities to other coronaviruses.
Viral upper respiratory diseases such as IBV (23), Newcastle disease (ND) (22), subtype H9 avian influenza virus infection (14), and infectious laryngotracheitis (35) are a serious problem in the field farms of China. Detection of IBV infection in poultry flocks is a major challenge because of the difficulty in differentiating this condition from other upper respiratory diseases. Appropriate diagnostic methods are needed and are an important tool for taking appropriate preventive steps. Another problem complicating IBV detection and diagnosis is the existence of at least nine IBV types co-circulating in vaccinated and non-vaccinated chicken flocks (23). The mAb 2D2 and its epitope identified in this study will be useful for clinical applications, and as a tool for further study of IBV detection and diagnosis. Since epitopes were mapped via expression in prokaryotes, the subclones allowed rapid screening of large numbers of mAbs, and more precise definition of the epitope's location. The truncated recombinant proteins generated in this study may be useful tools for mapping other epitopes on IBV N proteins in future studies.
Footnotes
Acknowledgment
This work was funded by a grant from the National Key Technology R&D Program of the Ministry of Science and Technology of the People's Republic of China (no. 2006BAD06A03).
Author Disclosure Statement
No competing financial interests exist.