
Abstract
There is limited information on the direct role of the neutralizing antibody responses against hepatitis C virus (HCV) infection or methodologies to study them. Previously we have demonstrated that interleukin-10 (IL-10) administered to chronic hepatitis patients led to a decrease in disease activity, but an increase in HCV viral burden. The mechanism behind this is unknown. The objective of this study was to examine the antibody response in IL-10-treated patients. To establish a neutralization antibody assay, HCV-positive and HCV-negative sera were collected and incubated with HCV strain JFH-1 particles before culture with Huh 7.5 cells. Viral replication was measured a week later by either indirect immunofluorescence assay (iIFA) or real-time reverse transcriptase polymerase chain reaction (RT-PCR). After validation of the methodology, the sera from 30 previously-described subjects of a group previously treated with IL-10 were tested for the neutralization capacity of their antibodies. The amount of total anti-HCV antibody in the sera was also measured by direct staining of HCV full-length replicon cells. With this validated neutralization assay for anti-HCV antibodies we found that HCV-neutralizing antibodies are universally present, but with significantly different titers. In patients who were treated with IL-10, the total anti-HCV antibody titers appear to be constant, but with significantly decreased antibody neutralization activity. Our study validates an assay to quantitatively determine the presence and strength of HCV-specific neutralizing antibodies. We have found that IL-10-treated patients have significantly lower HCV antibodies, but maintain the total anti-HCV antibody titer, suggesting a novel mechanism by which IL-10 treatment increases viral load in patients.
Introduction
The cytokine interleukin-10 (IL-10), produced by macrophages, monocytes, T cells, and B cells, has a significant role in the function and regulation of the immune system and in chronic HCV patients (6 –9). This cytokine is known to suppress the production of Th1 proinflammatory cytokines such as tumor necrosis factor-α (TNF-α), interferon-γ (IFN-γ), and IL-12, which are correlated with overt liver disease, including fibrosis and portal inflammation, and is able to enhance B-cell survival, and proliferation, and to stimulate the production of antibodies (10 –13). Our group previously observed that patients with chronic HCV, who had not responded to previous IFN-based therapy, had decreased disease activity after long-term IL-10 therapy (14). This was based on normalized serum alanine aminotransferase levels, decreased hepatic inflammation, and reduced liver fibrosis, via alterations in immunological viral surveillance, namely a decrease in CD4+ and CD8+ T cells (14,15). Unfortunately, the same treatments also led to an increased viral replication in these patients. Neither the mechanisms of increased viremia, nor the role of anti-HCV antibodies, were explored.
Analysis of antibody-mediated neutralization in individual patients as well as cohorts with well-defined viral isolates has enabled the study of neutralizing responses in the course of HCV infection, and characterization of the impact of neutralizing antibodies on viral infection (4,16 –19). The objective of our study was to examine the role of anti-HCV antibodies using the newly discovered infectious HCV culture system (JFH-1), and to use this technique to examine the anti-HCV antibody titers in the sera of HCV patients, including IL-10-therapy recipients (20 –22). Our study shows the presence of broad cross-reactive neutralization of antibodies from different HCV genotypes, with wide variation in the titers among individuals. We also show that administration of IL-10 in chronic HCV patients appears to reduce neutralizing antibodies, but increase the overall titer of the total anti-HCV antibodies, indicating a potential mechanism for the increased viremia observed during treatment. With this translational study, we provide an insight into the role of neutralizing antibodies in preventing virus entry, and its impact on HCV pathogenesis.
Materials and Methods
Patients
In order to validate our neutralization assay and evaluate cross-genotype differences, sera were collected from 150 HCV-positive and 5 HCV-negative patients that had provided informed consent prior to collection at the University of Florida. The samples were aliquoted into 1.5-mL microcentrifuge tubes before storing at −80°C. Patient information was recorded and each sample was assigned a non-identifiable number.
To explore the effect of recombinant IL-10 in chronic HCV, specifically the role of neutralizing antibodies in HCV infection, we used sera from a previously published study that was obtained from adult subjects with extensive fibrosis or cirrhosis who had previously failed to respond to IFN-based treatment (15). These patients were enrolled between February 1999 and September 2000. The protocol was approved by the Institutional Review Committee and the Clinical Research Center Scientific Advisory Board, and all study subjects provided written informed consent. Non-response to IFN-based treatment was defined as detectable HCV RNA at the end of 6 mo of IFN therapy. Exclusion criteria included decompensated cirrhosis, hemoglobin <12 g/dL, white blood count <3500 per cubic milliliter, platelets less than 75,000 per cubic milliliter, human immunodeficiency virus infection, pregnancy, and severe non-hepatic illnesses.
Study design and treatment regimens
The IL-10 study, as previously described, was a randomized three-dose trial (15). Briefly, 30 subjects were randomized (1:1:1 ratio; 10 subjects per group) to receive recombinant human IL-10 (rIL-10; Schering Plough Research Institute, Kenilworth, NJ) subcutaneously at a daily or thrice-weekly dose of 8 μg/kg, or a thrice-weekly dose of 4 μg/kg, for at least 12 mo. The participants were monitored as outpatients on a weekly basis during therapy, and 2 and 6 months after treatment. Serum HCV RNA level was assessed by branched-chain DNA assay (Quantiplex HCV RNA version 2.0; Bayer Diagnostics, Emeryville, CA), with a lower limit of detection of 200,000 genome equivalents (Eq)/mL. Negative samples were re-evaluated with a nested real-time reverse transcriptase polymerase chain reaction (RT-PCR).
HCV constructs and viral particle generation
pJFH-1 plasmid was a gift from Dr. Takaji Wakita (Tokyo Metropolitan Institute for Neuroscience, Tokyo, Japan). The linearized DNA was purified and used as a template for in vitro transcription using a MEGAscript kit (Ambio, Austin, TX). In vitro transcribed genomic JFH-1 RNA was delivered into Huh-7.5 cells by electroporation. Transfected cells were transferred to complete Dulbecco's modified Eagle's medium (DMEM) and cultured. Cells were passaged every 3–5 d, and at 21 d the supernatants were filtered and frozen at −80°C. Viral titer (1 × 105 ffu/mL) of JFH-1 stock was determined by the average number of NS5A-positive foci detected at the highest dilutions. Infection was done by adding supernatant or supernatant diluted in complete DMEM for 24 h before exchange. Control was uninfected Huh 7.5 supernatant.
Cells and HCV neutralization assay
The in vitro HCV infection assay uses HCV strain JFH-1 (genotype 2a) and the Huh 7.5 cell line. Five thousand Huh 7.5 cells were seeded onto a 24-well plate (30–50% confluency), and cultured overnight at 37°C and 5% CO2. The sera were incubated at varied dilutions (1:3, 1:5, 1:15, 1:25, 1:125, and 1:625), or at a 1:1 dilution (for IL-10 sera), in a total volume of 200 μL with 100 μL of 105 ffu/mL particles of HCV-JFH1 for 30 min at room temperature. The mixture was added to the Huh 7.5 cultures and incubated for 24 h. The supernatant was then removed and the cells were incubated in fresh medium for 6 more days. On the final day of incubation, the cells were washed with 1× PBS, and either fixed in 5% acetic acid and 95% ethanol at −20°C for 10 min for immunofluorescence, or Trizol was added to isolate RNA. HCV was measured by counting positive cells from immunofluorescence (similar to that described below), or by comparative real-time RT-PCR. The NS5A antibody used was as previously reported (23).
Reverse transcription and real-time reverse transcriptase polymerase chain reaction
Total cellular RNA was isolated from cells using Trizol (Invitrogen, Carlsbad, CA). Reverse transcription to obtain cDNA was performed using the Superscript II (50 U reverse transcriptase per reaction) first-strand synthesis for RT-PCR kit (Invitrogen), primed with oligo(dT) (Invitrogen), according to the manufacturer's instructions. Quantitative real-time fluorophore-labeled LUX primers and their unlabeled counterparts were obtained from Invitrogen. The primers for HCV were 5′-CGCAGGAGAGCCATAGTGGTCTG [FAM] G-3′, 5′-GAGCGGGTTGATCCAAGAAAG-3′; and for GAPDH were 5′-CGACCGGAGTCAACGGATTTGGT [JOE] G-3′, 5′-GGCAACAATATCCAGTTTAGCA-3′. The PCR conditions were: 50°C, 2′; 95°C, 2′ (95°C, 15′'; 60°C (HCV) or 62°C, (GAPDH) 30′'; and 72°C, 1 min) for 45 cycles. The reactions were conducted in a spectrofluorometric thermal cycler (MJ Research DNA Engine Opticon® 2 thermal cycler; Bio-Rad Laboratories, Inc., Hercules, CA). Fluorescence was monitored during every PCR cycle at the annealing step. The results were analyzed with MJ Opticon Monitor 3.1 software from Bio-Rad. Results are represented as means ± SEM.
Immunofluorescence for titration of anti-HCV antibody in the patient sera
Huh 7.5 cells transduced with HCV-JFH1 virus were grown on 24-well plates. The cells were fixed with 5% acetic acid in ethanol for 10 min at −20°C. After washing, the cells were first blocked with goat serum at a 1:50 concentration, washed with 1× PBS, and incubated with different dilutions (1:1, 1:10, and 1:50) of patient sera. Goat anti-human IgG conjugated with Alexa 488 (Southern Biotechnology Associates, Birmingham, AL) was used as the secondary antibody. Nuclei were counterstained with 4′,6-diamidino-2phenylindole (DAPI; Vector Laboratories Inc, Burlingame, CA), and cells were preserved with 50% glycerol. Examination was done with a fluorescence Leica DMIRE2 microscope, and images were analyzed with Open Lab 3.1.5 software. Positive cells and total cells were counted in the images to determine HCV-specific antibody titers in patient sera at 0 and 12 months post-treatment with IL-10. Three pictures were taken for each slide, for a total of three slides per serum, for a total of nine fields counted.
Results
HCV neutralization assay
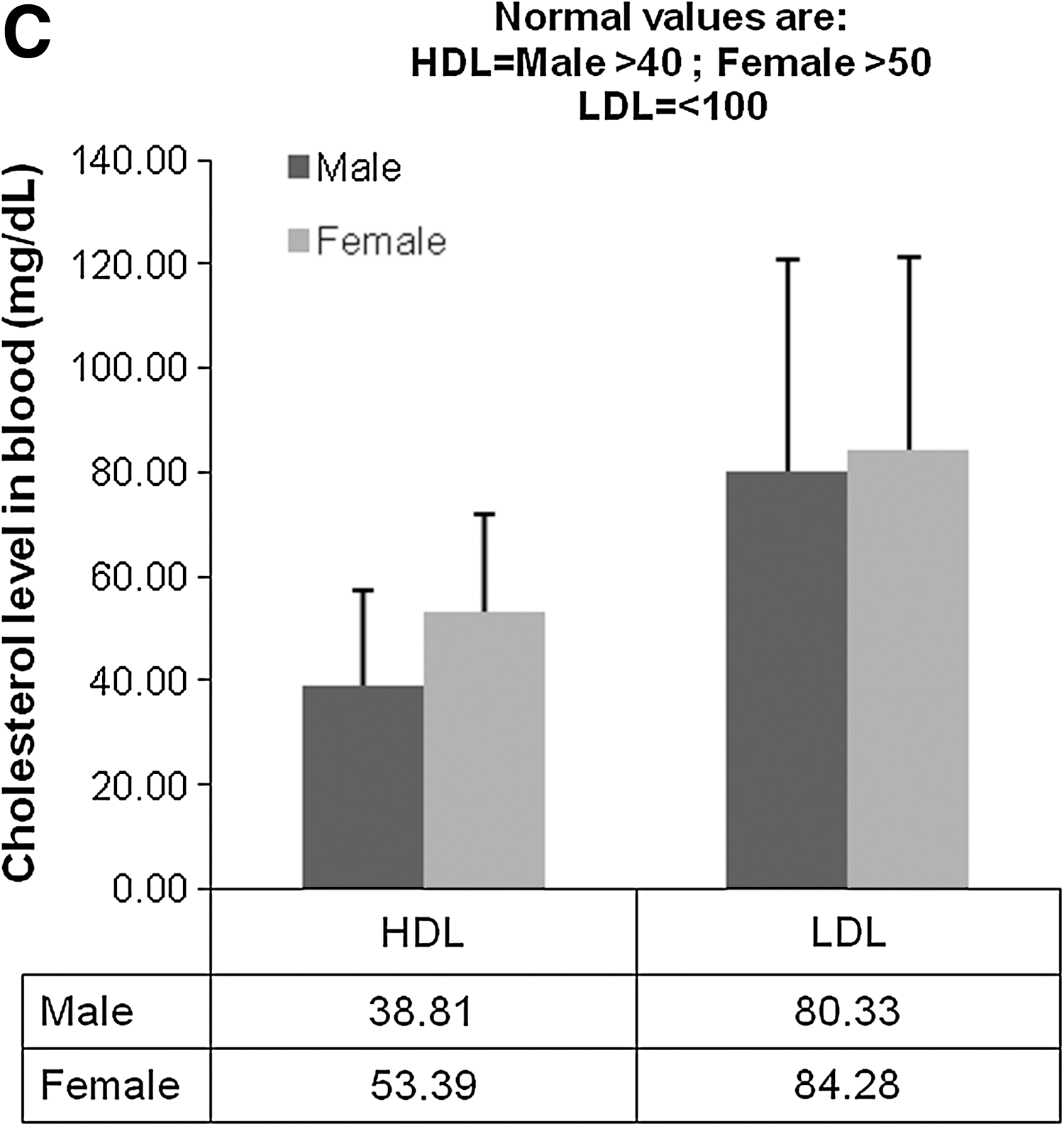
In order to study the neutralization activity of anti-HCV neutralizing antibodies in patient sera, we utilized direct neutralization of the fully replicative HCV strain JFH1 (genotype 2a). Initially, various dilutions of virus were used until an optimal amount, relative to the number of cells, was determined (i.e., 100 μL). Then 100 μL of 1 × 105 ffu/mL of virus stock was co-cultured with an equal volume of serum (or subsequent dilutions of this stock), and incubated for 30 min before culturing with the HCV-permissive Huh 7.5 cells. This allowed sufficient time for association between the virus and potential neutralizing antibodies before infection. After a week of culture, the cells were fixed for an indirect immunofluorescence assay (iIFA), or collected for RNA isolation. Representative images of the dilutions from one patient's serum are depicted in Fig. 1. Patient characteristics, including viral load and genotype, are tabulated in Table 1. As expected, patient sera showed strong neutralizing activity against the virus in a titer-dependent manner (Fig. 2A). The HCV-negative sera of lowest dilution was not able to reduce the amount of HCV-positive cells to the levels of the highest dilution of the positive sera (Fig. 2B). High-density lipoprotein (HDL) has been shown to attenuate the neutralizing activity of antibodies in HCV patient sera (24 –26). Since we used whole sera instead of isolated immunoglobulin, we correlated our results with the HDL levels in the patients tested, to corroborate that changes in our assays are not due to an artifact. While lower than normal, HDL levels were comparable among all patients (Fig. 2C).
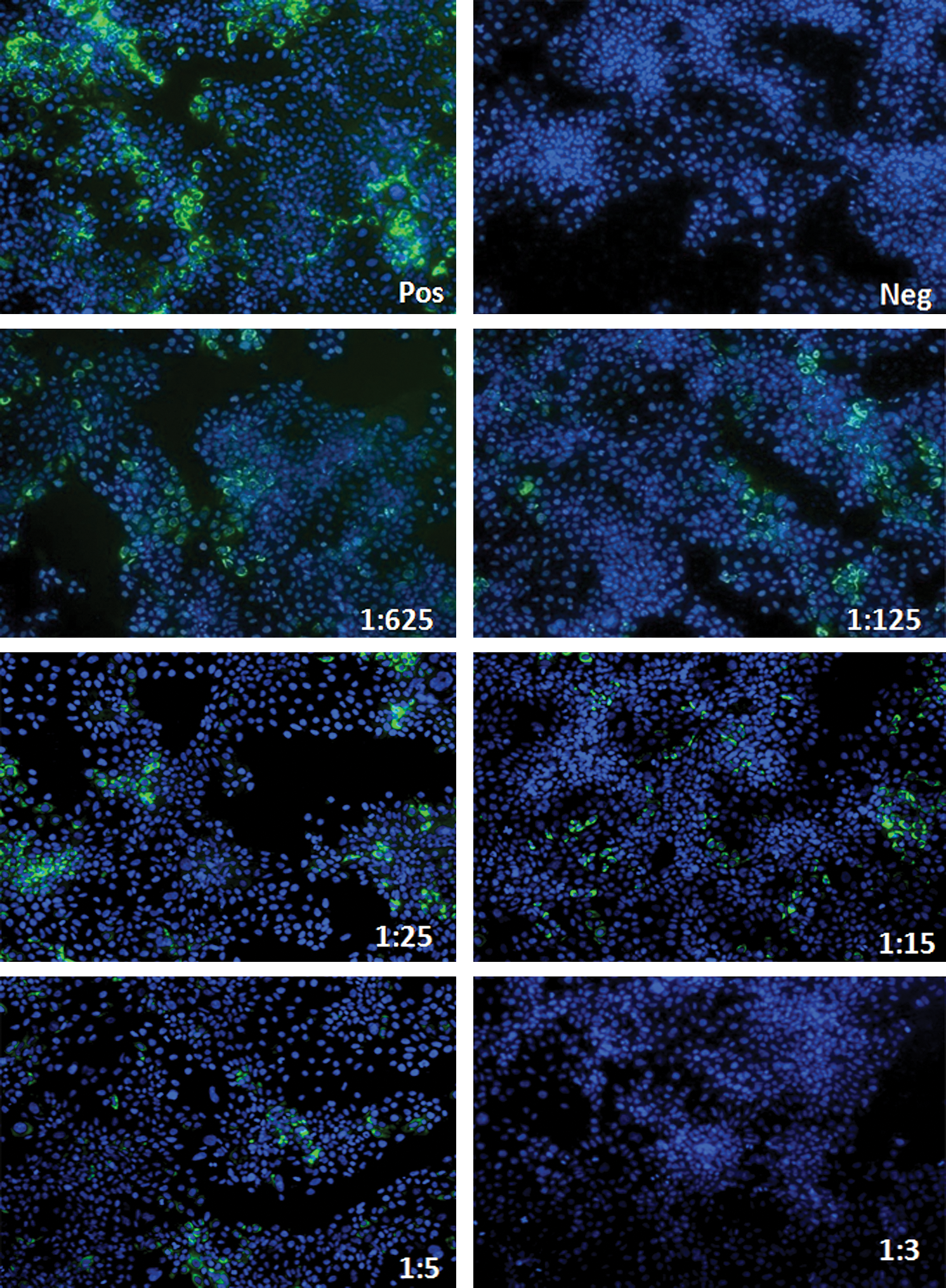
Representative iIFA images of HCV neutralization with patient serum. Cells were seeded at 5000 cells per well in a 24-well plate and infected with either 100 μL of virus/100 μL diluents (positive control), 200 μL diluents (negative control), or 100 μL virus with 100 μL of sera at different dilutions. Cellular nuclei are stained with DAPI (blue), and HCV NS5A protein is stained with FITC (green). Representative figure out of 150. Color images available online at
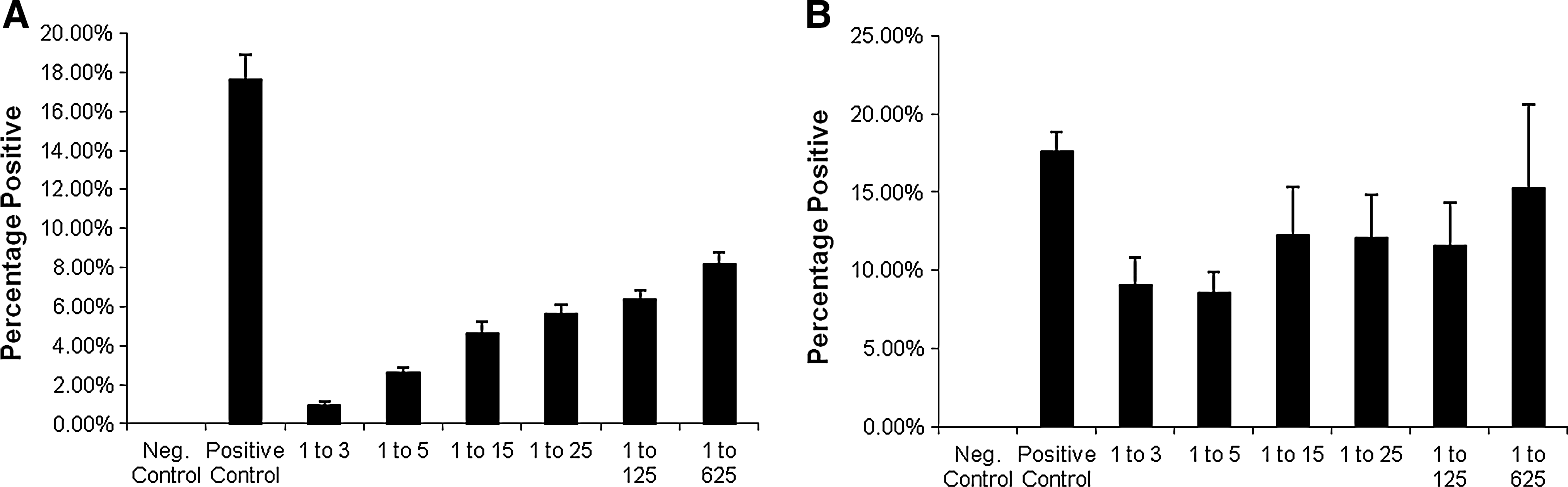
Neutralization of HCV-JFH1 by patient sera. HCV-positive or HCV-negative patient sera neutralization assay cell counts from immunofluorescence were averaged for each dilution. (
HCV, hepatitis C virus; AST, aspartate aminotransferase; ALT, alanine aminotransferase; NA, not applicable.
The HCV-positive patients were comprised of four genotypes. Genotype 1a/1b totaled 40 patients, genotype 2a/2b totaled 9 patients, genotype 3a/3b totaled 5 patients, and genotype 4a/4b totaled 6 patients. There were 90 patients, with average alanine aminotransferase and aspartate aminotransferase levels of 65.9 ± 6.03 IU/L and 72.4 ± 7.32 IU/L, respectively. The average viral load was 5.11 ± 0.15 log IU/mL. The HCV status of each serum sample was determined by database after all results had been completed.
Genotype cross-neutralizing activity of anti-HCV antibodies in patient sera
These results demonstrate that this simple methodology shows the neutralizing activity present in HCV-positive patient sera against genotype 2a. Since the JFH1 virus is of that genotype, it would be expected that genotype 2 patient-neutralizing antibodies would respond better in our system. To better understand how dependent this neutralization is on genotype, we observed the EC50 distribution in all genotypes tested, and calculated the average log of the median effective concentration required to induce 50% effect (EC50) of neutralization in the patients with known genotype and compared them. Fig. 3A through D shows the EC50 distribution among all of the genotypes. While there was a small bias towards genotype 2, there was no significant difference in the average log EC50 of the patients of the other genotypes (Fig. 3E). These results not only validate our assay, but also provide the basis for the notion of a cross-genotype protective antibody. Based on these results, neutralizing antibody titer varies considerably in the population, regardless of viral genotype.
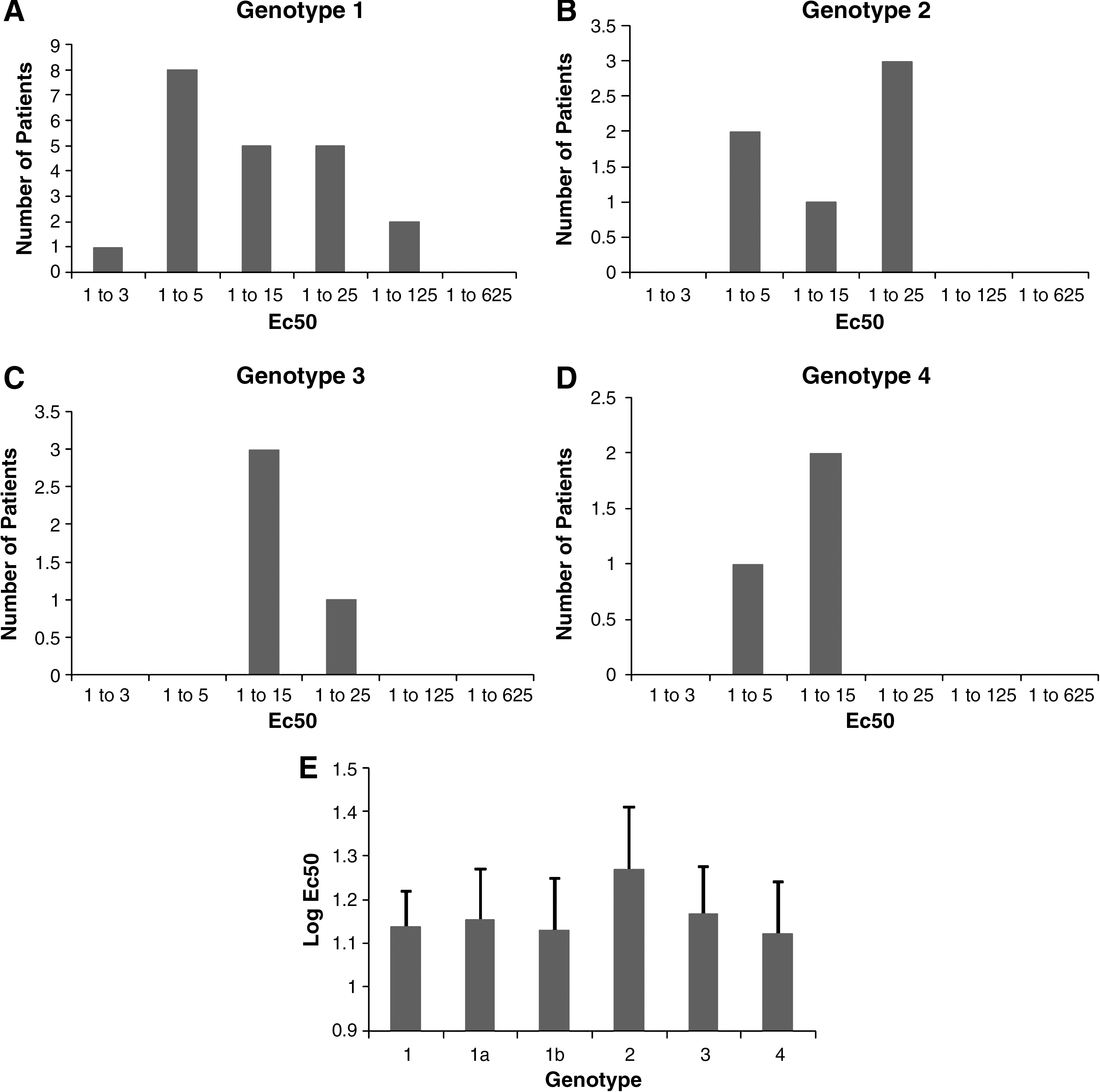
Genotype independence of the neutralizing antibodies. The EC50 dilution distribution was tabulated for each separate genotype available out of 150 patients. (
Neutralizing antibodies are increased in long-term IL-10-treated patient sera
As previously described, the patients were selected for this study based on their failure to respond to IFN treatment, as well as the presence of extensive fibrosis or cirrhosis of the liver (15). Ten patients were assigned per group based on the IL-10 dose received: 4 μg/kg three times per week (TIW), 8 μg/kg TIW, and 8 μg/kg daily (QD). Serum HCV RNA titers were assessed as described previously. We compared the results for the HCV RNA titer at the beginning of treatment with the HCV titer after 12 mo of IL-10 treatment. As previously shown, chronic IL-10 administration led to significant increases in serum HCV RNA levels (15). We were interested in evaluating the relationship of exogenous IL-10 administration, neutralizing antibody production, and HCV viremia.
Current reports in the HCV literature suggest the importance of antibodies in aiding in viral clearance, specifically the role of neutralizing antibodies that prevent entry of the virus into permissive cells (5,27). With the aid of the methodology described, we explored if there was any correlation of neutralizing antibodies in the sera of IL-10 recipients with the increased viremia. The IL-10-treated patient sera were incubated at a 1:1 ratio with infectious JFH-1 virions, as described above, with sera from the beginning of the study, and the 12-mo-treatment sera. RNA was collected and HCV replication was measured by real-time RT-PCR by the ΔΔCt method as compared to an endogenous control, the housekeeping gene GAPDH, and neutralization activity is depicted as the log10 of the inverse of the viral replication. The experimental control was the same amount of virus incubated alone in Huh 7.5 cells. There was a significant dose-dependent decrease in the neutralization of HCV by the patient sera after treatment with IL-10 (Fig. 4A).
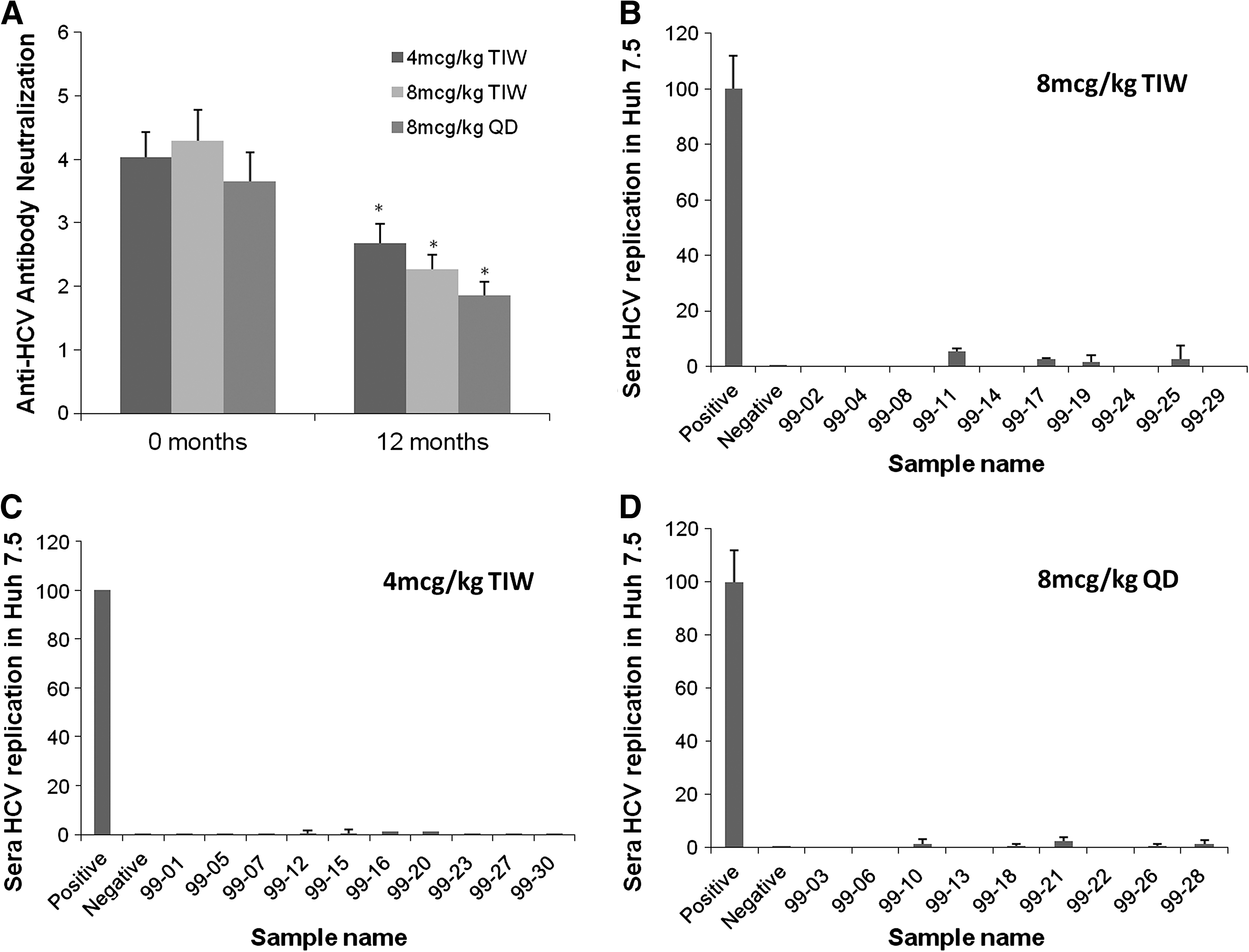
Patient serum neutralizing activity in vitro is decreased after 12 mo of treatment with rIL-10, as observed by increased viral replication: A neutralization assay was performed in which patient sera were incubated at a 1:1 ratio with HCV-JFH1 viral particles (100 μL of sera per 100 μL of 1 × 105 ffu/mL of HCV-JFH1 for 30 min at room temperature). Afterwards, each mixture was added to previously-seeded plates (5000 Huh 7.5 cells per well) and cultured for 24 h. At this point the wells were washed, and fresh media (virus-free) was added to the cultures and incubated for 6 more days. RNA was collected by Trizol isolation and real-time RT-PCR was run for HCV, with the housekeeping gene GAPDH as an internal control. Each point was calculated by the ΔΔCt method and compared against the positive control (virus alone in Huh 7.5 cells). Neutralization is represented as the log10 of the inverse of those results. (
This could indicate that the quality of the anti-HCV antibodies lost some of their neutralizing capacity in the sera of patients after long-term IL-10 treatment, but also, since we used whole serum, the increased replication could also be attributable to the increased HCV viral titers seen in the sera of IL-10-treated patients. Although no HCV strain other than JFH1 has been shown to replicate in culture with Huh 7.5 cells, we confirmed that the increase in viral replication in vitro was not due to other replicating virus in the sera by incubating 12-month sera (highest titer) in Huh 7.5 cells (Fig. 4B, C, and D). Comparisons of the amount of viral replication compared to negative and positive controls were measured by real-time RT-PCR, and showed no viral replication in any of the sera involved in the IL-10 study on Huh 7.5 cells. These results indicate that the replication observed in the neutralizing antibody assay were indeed due to replication of HCV JFH1, and an increase in its replication strongly correlates with a decrease in neutralizing antibodies present in the sera of IL-10-treated patients after 12 months.
Total anti-HCV antibodies are increased in the sera of IL-10-treated patients
It is known that IL-10 modulates B-cell activity. To determine the effect of IL-10 on total HCV antibodies, we performed iIFA assays with the sera as the primary antibody against pJFH1-electroporated Huh 7.5 cells (the source of our virus). Three dilutions of sera were used, and the secondary antibody, anti-human IgG conjugated with Alexa 488, was used to select for IgG antibodies only. Nuclei were counterstained with DAPI (blue). A representative figure of the images taken is shown in Fig. 5B and compared to negative staining (Fig. 5A), and positive-staining cells stained with anti-NS5A antibody (staining of Huh 7.5 cells with no virus was also negative; Fig. 5C). Cells with cytoplasmic staining were counted as positive. The averages of the percentages of positive cells were calculated from three separate images at 100× magnification. In general, chronic IL-10 administration significantly increased the titer of total anti-HCV IgG in the serum of infected patients (Fig. 5D).
HCV antibody titer in patient sera increases after 12 mo of IL-10 treatment. To test for HCV antibody titer in the patient sera we performed three dilutions of sera and used a secondary antibody (anti-human IgG conjugated with Alexa 488 [green]). Nuclei were counterstained with DAPI (blue). (

Discussion
In HCV, throughout the acute phase of the infection, B cells generate antibodies against viral proteins and these include neutralizing antibodies, but the direct contribution of these antibodies to HCV has not been examined in depth due to the lack of a fully replicative virus (2,28,29). Based on studies of other systems (animal models, clinical data, and virus like particles), it has been determined that there seems to be a correlation between the induction of neutralizing antibodies and the HCV viral load (24,30 –33). In this study we first validated a simple methodology for the study of viral neutralization, based on the fully-replicative HCV strain JFH-1, similarly to a recently described technique by Fournier et al. (20). Our methods produced similar results, demonstrating that this technique can be used for the study of viral neutralization. We found that the sera cross-reacted with the other genotypes tested, confirming the broad specificity of these antibodies against HCV (JFH-1 is of genotype 2a) (34 –41). This observation raises another set of questions about why these antibodies are ineffective against HCV, and what role they play in the setting of chronic infection. Based on our results, we extended the use of this methodology to the study of neutralization in the sera of long-term IL-10-treated recipients.
HCV viral clearance and disease are mediated by a strong T-cell response because of their cytotoxic nature, as well as other effector functions (42 –44). One of these other functions is to induce the production and maturation of antibodies against the virus by signaling to more B cells. Since the time of our study on the role of IL-10 therapy, understanding of the role of this cytokine in intracellular infections has been garnered, and it has been shown to prevent the eradication of pathogens, establishing a chronic state in genetically-deficient mice (45,46). In the context of an HCV infection, a strong Th-1 response is observed in those who clear an acute infection; whereas a weak Th-1 response with a more pronounced Th-2 response, including IL-10, is seen in those with chronic infection (15,47,48). Our original study shed light on the role that IL-10, a type 2 cytokine, plays in modulating the cellular response to HCV, for which Th-1 cells were shown to be downregulated (15,49). In this study we furthered our understanding of in vivo administration of IL-10 in chronic HCV patients, and observed a strong correlation between the increases in the viral load and a decrease in neutralization capacity. We also observed that this decrease in neutralization activity is associated with a significant increase in the overall titer of anti-HCV antibodies.
The correlation we observed with neutralizing antibodies needs to further be assessed to clarify whether the loss of antibody neutralization was a direct consequence of IL-10. Whether the effect seen on T cells is the cause of the neutralizing antibody decline remains unknown. What is known is that HCV may also infect a small number of B cells, and that binding to CD81 (part of the B-cell receptor complex) disrupts receptor-associated signaling pathways (37,50 –56). Furthermore, it is not understood how this affects the antibody response. Our investigation into IL-10 recipients suggests that the study of B cells in HCV infection should be pursued further because of the disparities between total antibody and the neutralization activity of those antibodies. Similarly, Bartosch et al. observed the presence of higher titers of HCV antibodies, but no neutralizing antibodies, in HCV-infected chimpanzees (57). Indeed, a direct interaction of the virus with B cells may lead to somatic hyper-mutation, variable chain rearrangement, and isotype switching, such as that observed with recombinant E2 protein or HCV infection (52). They also showed that the levels of activation-induced cytidine deaminase (AID, which induces the mutational process in the somatic hyper-mutation process of B cells) mRNA are increased with HCV infection. In hybridoma studies, at least, it indicates that these effects require direct replication of HCV in B cells.
Our data could also indicate only a change in the type of antibodies being produced. For instance, the E2 envelope glycoprotein has multiple epitopes recognized not only by neutralizing antibodies, but also by infectivity-enhancing antibodies, and while both of these are present in the host, the ratio of these antibodies decides the outcome of the infection (39,59). This phenomenon has also been observed in other Flaviviruses such as dengue virus (59). Here, we furthered our initial understanding of the role of IL-10 in inducing an increase in viremia by observing that a change in both quantity and quality of the anti-HCV antibodies may be contributors to the success of HCV observed after year-long administration of this cytokine. Furthermore, the neutralizing antibodies found in vivo are among a mix of anti-HCV antibodies of which the majority have no benefit for the host against the infection (60 –64). Although we have no data to show if this phenomenon could be extrapolated to the acute phase versus the chronic phase of the infection, the decrease in neutralizing activity has been seen to decrease from the initial phase of the infection (52). Defining how HCV interacts with neutralizing antibodies would provide novel insights into the biological role of these antibodies in controlling infection, which is a basis for developing effective vaccines against HCV.
Footnotes
Acknowledgments
We would like to acknowledge the Center for Mammalian Genetics and Dr. Terada's laboratory for allowing us the use of their equipment, and Mr. Yingda Xi for his support with the real-time PCR. Huh 7.5 cells were provided by Dr. Charles Rice (Rockefeller University, New York, NY), and the HCV-JFH1 plasmid was kindly provided by Dr. Takaji Wakita. Ms. Eksioglu is an National Institutes of Health Ruth Kirshchtein pre-doctoral fellowship recipient: 1F31AI071353. This work was funded in part by grants NIH DK002958 and AI061158 to C.L., and NIH-AI-064357 to D.R.N.
Author Disclosure Statement
None of the authors have any conflicting interests or commercial affiliations linked to this work.