
Abstract
Mannose-binding lectin (MBL) is an important constituent of the human innate immune system, and can bind to a wide range of pathogens, including viruses such as influenza A, HIV, herpes simplex 2, and SARS-CoV. MBL deficiency results from single nucleotide polymorphisms (SNPs) in exon 1, and the promoter region of the human MBL2 gene has been found to be associated with susceptibility to a number of infections. However, studies on the interactions between MBL and CMV infection are limited. In this study, we investigated 104 children suffering from HCMV infection, in an effort to find any association between MBL and HCMV infection of children in China. We analyzed the genotypes of 104 HCMV patients and 105 healthy controls, and investigated the distributions of polymorphisms at −550(H/L), −221(Y/X), and +4(P/Q), together with their structural variants. Although there was no significant difference in the distribution of B alleles between HCMV patients and healthy controls, the frequencies of the high-MBL-level related genotype of YA type in HCMV patients were significantly lower than those seen in healthy controls, while low-level related genotypes of XB type were more common in HCMV patients. In addition, CMV-DNA quantification revealed higher viral loads of the XB type in HCMV patients. Thus we can speculate that as an acute response protein and a pattern-recognition molecule of the innate immune system, MBL may play a role in protecting against HCMV infection in children, and MBL gene mutations may be a significant risk factor for the development of infantile HCMV infection.
Introduction
Mannose-binding lectin (MBL) is an important constituent of the human innate immune system, especially for infants who are in the “window of vulnerability” (approximately 6–24 months of age), when maternal immunoglobulin G (IgG) antibody levels have waned, but the infants' own adaptive immune response is still immature (3). It has been demonstrated that MBL can bind to mannose or other carbohydrate components presenting on the surface of a wide range of microorganisms, and activate complement via the lectin pathway to lyse pathogens and increase phagocytosis. Serum levels of MBL vary widely due to three single nucleotide polymorphisms (SNPs) in exon 1 of the human MBL2 gene on chromosome 10. Three SNPs, those at codon 52 (CGT-TGT), codon 54 (GGC-GAC), and codon 57 (GGA-GAA), encode for variant alleles D, B, and C, respectively, which reduce functional MBL levels by impairing the assembly of MBL monomers into functional oligomers, and A indicates the wild allele. Three SNPs at positions −551(H/L), −221(X/Y), and +4(P/Q) in the 5′-flanking region also influence the MBL serum level, with H > L, Y > X, and Q > P. Promoter haplotype HYP is frequently associated with high levels of MBL, and LXP is associated with low levels of the protein. As a result of linkage disequilibrium, only seven haplotypes are observed, namely HYPA, LYPA, LYQA, LXPA, HYPD, LYPB, and LYQC.
Previous studies have found that MBL can bind several viruses, such as influenza A, HIV, herpes simplex 2, and SARS-CoV (4). However, studies of the interactions between MBL and HCMV are limited. In this study, we investigated 104 children suffering from HCMV infection, in an effort to find any association between MBL and HCMV infection of children in China.
Materials and Methods
Patients and controls
In all, 104 children diagnosed with HCMV infection (64 boys and 40 girls, mean age 4.29 mo, median 2 mo, range 1–24 mo) were investigated. All patients were inpatients at Children's Hospital, Zhejiang University School of Medicine, between March and December of 2007. All patients were seropositive for CMV-IgM antibody as tested by ELISA, and HCMV DNA varied from 4 × 103 copies/mL to 7 × 106 copies/mL as detected by quantitative PCR in urine. All patients had clinical symptoms, and single- or multiple-organ damage was observed, including hepatitis, pneumonitis, colitis, thrombocytopenia, and involvement of the central nervous system. We used 105 healthy symptom-free children negative for CMV-IgM and CMV-IgG from the same area as controls. Both patients and controls were Chinese Han children, and written consent was obtained from the parent or guardian. The study protocol was approved by the local ethics committee.
MBL measurement
The concentration of serum MBL was determined in 105 healthy children by sandwich enzyme immunoassay using a human MBL ELISA kit (HK323; Hycult Biotechnology, Uden, The Netherlands). Diluted samples and standards were incubated in microtiter wells coated with mannan at room temperature for 1 h. After washing, the plates were incubated for 1 h with diluted tracer, followed by incubation for 1 h with diluted streptavidin-peroxidase conjugate. Color was developed using TMB substrate solution, and the reaction was stopped by adding stop solution. The plates were placed in a spectrophotometer and the absorbance was measured at 450 nm.
DNA sample
Genomic DNA was extracted from the blood of a total of 209 cases, including both patients and controls, according to a standard phenol-chloroform procedure.
MBL2 genotyping
Six SNPs in the MBL2 gene (−550G/C, −221C/G, and +4C/T in the 5′-flanking region, and codon 52 CGT/TGT, codon 54 GGC/GAC, and codon 57 GGA/GAA in exon 1) were analyzed using a sequencing-based typing method. A DNA fragment including promoter region and exon 1 (953 bp) was obtained by polymerase chain reaction (PCR) amplification using the sense 5′-CCTGCCAGAAAGTAGA GAGG-3′ and anti-sense 5′-CCAGGCAGTTTCCTCTGGA AGG-3′ primers. The cycling conditions were 95° for 5 min, 32 cycles of 95° for 35 sec, 59° for 35 sec, 72° for 45 sec, and finally 72° for 10 min. The PCR products were tested by agarose gel electrophoresis, and subjected to DNA sequencing.
Statistical analysis
The chi-square test was used to test the null hypothesis for the distribution of the allelic and genotype frequencies between patients and controls. The risk of disease associated with MBL2 genotype as a genetic factor was estimated using the calculation of odds ratios with 95% confidence intervals (CI). Comparison of MBL serum levels was done using the Mann-Whitney U test. All tests used were two-tailed, and p values <0.05 were considered significant. Statistical analysis was performed using SPSS V. 11.0 (SPSS, Inc., Chicago, IL).
Results
MBL concentration related to genotype
In this study, we did not find the C and D alleles in exon 1 of the MBL-2 gene, and thus detected only four polymorphic sites (H/L at −550, Y/X at −221, P/Q at +4, and A/B at codon 54). According to previous studies (4,5), as a result of linkage disequilibrium, only five haplotypes were observed, namely HYPA, LYPA, LYQA, LXPA, and LYPB. In this study, 1 or 2 of 4 polymorphic sites were heterozygotic, and by combining them, we could determine the genotype of the MBL2 gene. For example, when two sites (P/Q and A/B) were heterozygotic, the −550 site must be L, the −221 site must be Y, and thus genotype LYQA/LYPB was determined instead of LYPA/LYQB. Thus H/L, Y/X, P, and A combined to form HYPA/LXPA; H/L, Y, P/Q, and A combined to form HYPA/LYQA, and so on. In this study, 14 different genotypes were combined. According to a previous study (5), these genotypes were divided into two groups: the YA-type (homozygously carrying both the Y and A allele), and the XB-type (containing the LXPA or LYPB haplotype, heterozygously). We found that the latter is associated with lower MBL levels (1551.18 ± 1044.74 ng/mL versus 730 ± 425.70 ng/mL; p < 0.001; Fig. 1).
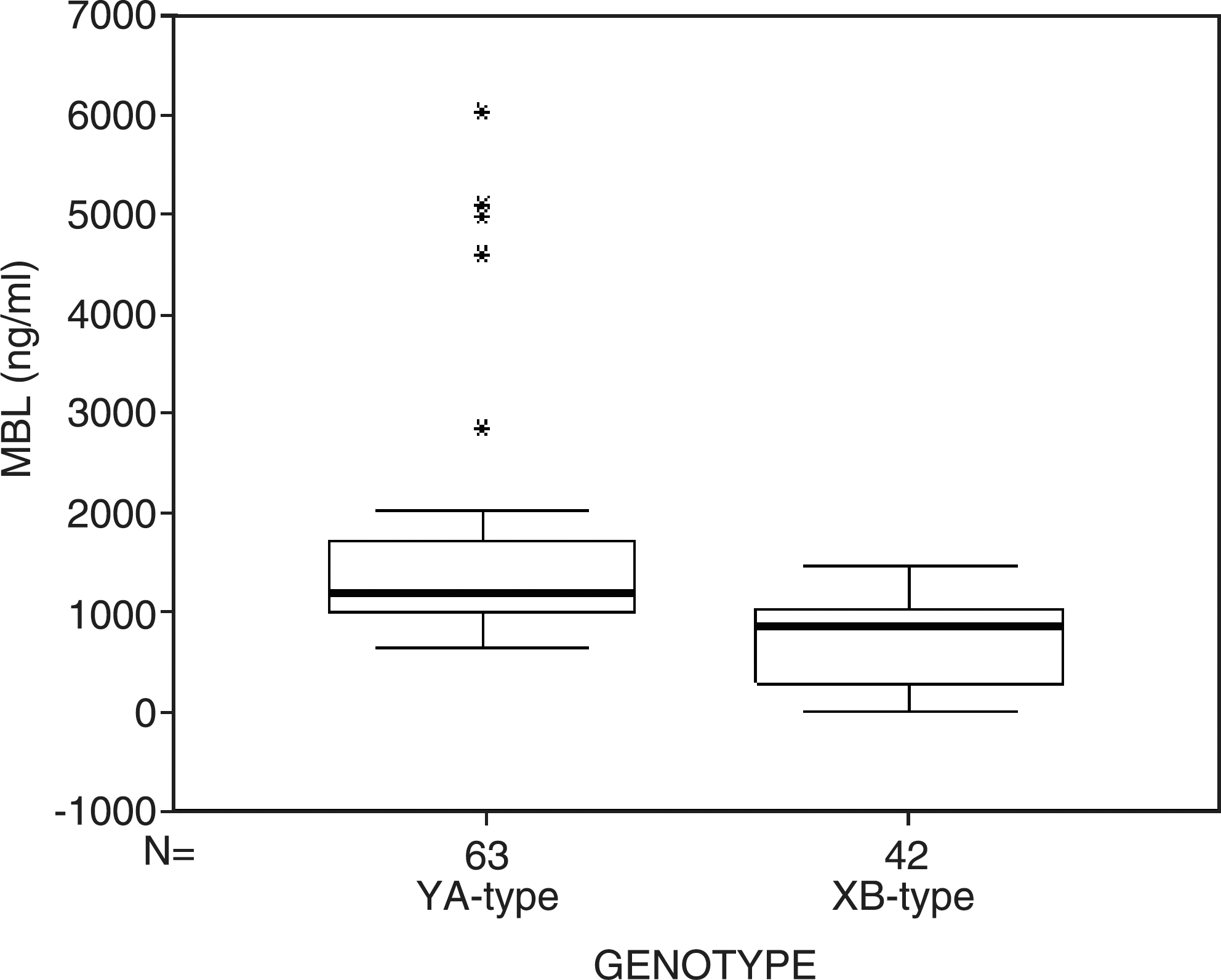
Serum levels of MBL in relation to MBL2 genotypes in 105 healthy controls (*refers to outlier).
Comparison of MBL2 genotypes between HCMV patients and healthy controls
The distribution of MBL2 SNPs in the promoter and structural regions in 104 HCMV-infected patients and 105 healthy controls are summarized in Tables 1, 2, and 3. All genotype distributions were consistent with the existence of Hardy-Weinberg equilibrium. There was no significant difference in the distribution of the B allele in HCMV patients and healthy controls (18.3% versus 15.2%; p = 0.41). The frequency of the B/B homozygote was low both in patients (1/104, 1%), and controls (3/105, 2.9%). The frequency of the A/B heterozygote was higher, but was not statistically significantly so in patients compared to controls (34.6% versus 24.8%; p = 0.119). Also, there was no significant difference found at the −221 site in the promoter region. However, at the −550 site in the promoter region, the frequency of the H/L heterozygote was significantly higher in patients than controls (56.8% versus 34.3%; p = 0.001; OR 2.51, 95% CI 1.44,1.40), while the frequency of the H/H homozygote was lower in patients than controls (19.2% versus 45.7%; p = 0.000; OR 0.28, 95% CI 0.15,0.55). Haplotype analysis showed that the frequency of wild-type HYPA was also lower in patients than in controls (47.6% versus 62.9%; p = 0.002; OR 0.54, 95% CI 0.36,0.97). The XB-type MBL2 genotypes had a significantly higher frequency in CMV patients than in controls (58.7% versus 40%; p = 0.007; OR 1.47, 95% CI 1.10,1.95).
p < 0.05; odds ratio (OR) and 95% CI for the H/H genotype: OR 0.28, 95% CI 0.15,0.55; for the H/L genotype: OR 2.51, 95% CI 1.44,4.40.
Comparison of HCMV profiles between YA-type and XB-type in HCMV patients
The serum levels of ALT and AST were not significantly different in YA-type and XB-type in HCMV patients. However, CMV-DNA quantification revealed higher viral loads in the XB-type group in HCMV patients (p < 0.05; Table 4).
Values given are median (interquartile range).
Discussion
Mutations in exon 1 of the MBL2 gene have been shown to differ in various populations. In this study, only mutation in codon 54 (B allele) was found, consistent with the results of previous studies, that showed that alleles C (codon 57) and D (codon 52) were present at extremely low frequencies and nearly absent in the Chinese population (6,7). The frequency of the B mutant in this study was 15.2%, which was close to the 20% level found in a previous report (7).
MBL2 genotype and serum MBL levels show a relatively strong correlation, and studies often just use one of these to infer deficiency. In studies based on MBL concentrations, the cutoff values defining MBL deficiency differed, with 500 ng/mL (8) sometimes suggested, but in some cases 100 ng/mL (9) or 1000 ng/mL (10) have also been used. And in some studies of the MBL2 genotype, only mutations on exon 1 were considered, overlooking the fact that the promoter genotype LXP was also related to low MBL levels. Previous reports have indicated that the promoter variants are associated with decreased levels of serum MBL. The HY haplotype is associated with high levels of MBL, the LY haplotype correlates with intermediate levels of MBL, and the LX haplotype is linked to low levels of MBL. The B allele resulted in the replacement of critical axial glycines by dicarboxylic acids, which distorts the collagen triple helix of trimeric subunits, and reduces the proportion of higher-order oligomers in the circulation, which are required for activation of the complement system. It was also noted that the B-allelic form of MBL failed to bind MBL-associated serine protease (MASP). When these observations are taken together the MBL, which is transcribed from the XB-type MBL gene (containing the LXPA or LYPB haplotype, heterozygously), is most likely a defective molecule in terms of quantity and quality (5).
In this study, we investigated the distribution of genetic polymorphisms at sites −550(H/L), −221(Y/X), and +4(P/Q) in the promoter region, together with their structural variants. There was no significant difference in the distribution of the B allele between HCMV patients and healthy controls. We could found that at the −550 site in the promoter region, the frequency of the H/L heterozygote was significantly higher in patients than controls (56.8% versus 34.3%; p = 0.001; OR 2.51, 95% CI 1.44,1.40), while the H/H homozygote was lower in patients than controls (19.2% versus 45.7%; p = 0.000; OR 0.28, 95% CI 0.15,0.55). But polymorphisms of the H/L allelic site did not influence the levels of MBL as significantly as Y/X, because LYA always related to high MBL levels. However, when we take account of the entire genotype, we found that the frequency of the high-MBL-level related genotype YA-type in HCMV patients was significantly lower than in healthy controls, while the low-level related genotype XB-type was more common in HCMV patients (58.7% versus 40%; p = 0.007; OR 1.47, 95% CI 1.10,1.95). In addition, CMV-DNA quantification revealed higher viral loads in the XB-type group in HCMV patients (p < 0.05). Thus we could speculate that as an acute response protein and a pattern-recognition molecule of the innate immune system, MBL may play a role in protecting against HCMV infection in children. Low-protein-level-related gene mutations of MBL may be a significant risk factor for the development of infantile HCMV infection.
MBL, like C1q, is a pattern-recognition molecule of the innate immune system that has the ability to recognize and bind various pathogens, including viruses such as influenza A, HIV, herpes simplex 2, and SARS-CoV. Once it is bound, MBL is able to activate the complement system to lyse pathogens directly or to increase phagocytosis. This antiviral effect may occur through phagocytosis of the virus following opsonization by C3b, lysis of the virus by the membrane attack complex, or neutralization by coating the virion with complement components that prevent viral binding to its cellular receptors (11).
There has been no investigation that directly demonstrated that MBL can bind HCMV, and it is difficult to predict MBL binding, because some organisms contain mannose, but to which MBL does not bind, and binding changes with growth phase with no change in sugar composition (12). It has been reported that CMV-infected fibroblasts activate complement via the classical complement pathway independent of specific anti-CMV antibody, and incubation of CMV-infected fibroblasts with serum deficient in complement components revealed that C1q, but not MBL, was required for complement activation (13).
In contrast to in vitro studies, two recent clinical studies showed that MBL may play a role in the control of CMV infection. Manuel et al. (14) investigated 16 kidney transplant recipients with high-risk CMV serostatus (donor positive/recipient negative, D+/R−), and found that MBL deficiency may be a significant risk factor for the development of post-prophylaxis HCMV infection in D+/R− kidney recipients. Khotivari and Mikeladze (15) also reported that the concentrations of MBL in the plasma of pregnant women suffering from HCMV infection were highly elevated to 1.278 ± 0.144 mg/mL, compared to the levels in normal women of 0.152 ± 0.025 mg/mL, and this also demonstrates an association between MBL and HCMV infection.
In this study, we found that low-MBL-level-related genotypes were more common in HCMV-infected children than in healthy controls. These data support an association between MBL and HCMV infection. However, further study is needed to define the precise role of MBL in HCMV infection.
Footnotes
Acknowledgment
This work was supported by the Zhejiang Provincial Natural Science Foundation of China (grant no. Y206070) and National Natural Science Foundation of China (No. 30872240).
Author Disclosure Statement
No competing financial interests exist.