
Abstract
Swine-origin pandemic 2009 A (H1N1) influenza viruses are still infecting humans, and humans are currently being vaccinated with the inactivated vaccine of 2009 A (H1N1) influenza virus. We wanted to determine the efficacy of 2009 A (H1N1) inactivated vaccine in ferrets. Ferrets immunized with one dose (7.5 μg) of 2009 A (H1N1) inactivated vaccine were not protected from infections of either pandemic H1N1 or seasonal H1N1 influenza viruses, while ferrets immunized with two doses of 2009 A (H1N1) inactivated vaccine were protected from infections of pandemic H1N1, but not seasonal H1N1 influenza viruses. IgG subtype of antibody was dominantly detected in tissues of immunized ferrets. Our study suggests that pandemic H1N1 vaccine may not elicit the antibody cross-reactive to the seasonal H1N1 influenza virus.
Introduction
Genetic analysis of 2009 A (H1N1) influenza viruses showed that they were novel influenza viruses that had not been previously reported in swine and humans, but were derived from swine influenza viruses (4,23,26). The M and NA genes belong to the Eurasian swine lineage of influenza viruses that were derived from the wholly avian influenza viruses. The NS, NP, and HA genes are derived from the classical swine influenza viruses that entered swine populations in 1918. The PB2 and PA genes are of avian origin and are phylogenetically close to the triple reassortant swine influenza viruses that entered swine around 1998. The PB1 genes are found in triple reassortant swine influenza viruses and were seeded in influenza viruses around 1969.
The clinical symptoms of patients infected with 2009 A (H1N1) influenza viruses include fever, cough, dyspnea (respiratory distress), increased serum lactate dehydrogenase levels, vomiting, diarrhea, and bilateral patchy pneumonia (1,24). Animal studies using mice and ferrets showed that 2009 A (H1N1) influenza viruses are more pathogenic than seasonal H1N1 influenza viruses (12,21,22). In contrast to seasonal H1N1 influenza viruses, the 2009 A (H1N1) influenza viruses can infect the lungs of animals.
Domestic ferrets (Mustela putorius furo) are considered the most relevant small animal model for human influenza infections since the infection course and clinical signs in ferrets closely resemble those in humans. Thus ferrets have been used to study influenza pathogenesis and vaccine efficacy (6,15,18,20). In this study we wanted to determine whether ferrets immunized with one or two doses of the inactivated vaccine of 2009 A (H1N1) influenza viruses were protected from infection by 2009 A (H1N1) influenza viruses or seasonal H1N1 influenza viruses, and which subtypes of antibodies were responsible for protecting the immunized ferrets.
Materials and Methods
Viruses
The new swine origin 2009 A (H1N1) influenza viruses, A/California/04/09 and A/California/07/09, were kindly provided by the WHO Collaborating Center for Influenza, Centers for Disease Control and Prevention. One amino acid of HA or NA protein is different between A/California/04/09 and A/California/07/09. The 2009 A (H1N1) viruses and the seasonal H1N1 influenza virus, A/Brisbane/59/07, were grown in 10-day-old hens' eggs before they were used for this study. All work using 2009 A (H1N1) was performed in a BSL-3 facility approved by the Korean government.
Generation of reassorted 2009 A (H1N1) vaccine virus
The reassorted 2009 A (H1N1) vaccine virus (CNUK-RG A/CA/4xPR/8) was rescued in Vero cells as previously described (7,29). Plasmids of PB2, PB1, PA, NP, M, and NS of A/PR/8/34 (H1N1) were kindly provided by Dr. Robert G. Webster at St. Jude Children's Research Hospital, Memphis, TN.
RNAs were extracted from the allantoic fluids containing A/California/04/09 (H1N1) using the RNeasy kit (Qiagen, Valencia, CA). cDNA was made with uni12 primer (5′-AGCAAAAGCAGG-3′) and the ImProm-II™ Reverse Transcription System (Promega Corp., Madison, WI), according to the manufacturer's instructions. HA or NA genes of A/California/04/09 (H1N1) were amplified by PCR using the Expand High-Fidelity PCR System (Roche Diagnostics, Mannheim, Germany) according to the manufacturer's instructions. The HA gene was amplified with the following primers: Bm-HA-1 (5′-TATTCGTCTCAGGGAGCAAAAGCAGGGG-3′) and Bm-NS-890R (3′-ATATCGTCTCGTATTAGTAGAAACAAGGGTGTTTT-5′). The NA gene was amplified using the following primers: Ba-NA-1 (5′-TATTGGTCTCAGGGAGCAAAAGCAGGAGT-3′) and Ba-NA-1413R (5′-ATATGGTCTCGTATTAGTAGAAACAAGGAGTTTTTT-3′). The PCR products were cloned into the vector pHW2000 as previously described (11).
The reassorted 2009 A (H1N1) vaccine viruses were rescued from cloned plasmids in the BSL-3 facility. Vero cells grown to 80% confluence in a flask (75 cm2) were trypsinized and resuspended in 8 mL of Opti-MEM I (Invitrogen, Carlsbad, CA). Then 20 mL of fresh Opti-MEM I was added to 2 mL of resuspended cells, and 3 mL of diluted cells (about 1.5 × 106 cells per well) were added to each well of six-well tissue culture plates (Greiner Bio-one, Kremsmünster, Czech Republic). The cells were incubated in humidified conditions (5% CO2, 37°C) overnight. The next day, 1 μg of each plasmid and 18 μL of TransIT LT-1 transfection reagent (Panvera, Madison, WI) were added to Opti-MEM I to a final volume of 200 μL, and the mixture was incubated at room temperature for 45 min before 2 mL of Opti-MEM I was added. The mixture was added to the cells in each well of a six-well plate after the medium was removed. The transfected cells were incubated for 48 h, then 300 μL of supernatant treated with L-(tosylamido-2-phenyl) ethyl chloromethyl ketone (TPCK)-treated trypsin (1 μg/mL) were inoculated into the allantoic cavity of 10-day-old pathogen-free hens' eggs (Charles River SPAFAS, North Franklin, CT). The presence of rescued viruses in the allantoic fluids in the inoculated eggs was confirmed by genetic analysis and hemagglutination-inhibition (HI) assays using swine H1-specific antibodies.
Preparation of 2009 A (H1N1) inactivated vaccine antigens
The 2009 A (H1N1) vaccine virus was propagated in the allantoic cavities in 10-day-old hens' eggs at 35°C for 60 h. The allantoic fluid was harvested and clarified by centrifugation at 3000 rpm for 15 min. The supernatant was concentrated to 1/10 of the original volume using Amicon concentrator apparatus. The concentrated viruses were purified using 20% sucrose cushion at 26,000 rpm (4°C) for 50 min. The pellet was resuspended in PBS (pH 7.4), sonicated for 3 min, and centrifuged through a continuous gradient of 20–75% sucrose at 26,000 rpm (4°C) for 2 h. Bands containing vaccine antigens were collected using a syringe, diluted in PBS (pH 7.4), and pelleted by centrifugation at 26,000 rpm (4°C) for 1.5 h. The pellets were resuspended in PBS (pH 7.4), sonicated, and treated with formalin (0.02% per volume) at 4°C overnight. The pellets treated with formalin were centrifuged at 26,000 rpm (4°C) for 2 h.
The hemagglutinin protein content in the prepared 2009 A (H1N1) vaccine was determined by standard single radial immunodiffusion technique as previously described (30), using 2009 A (H1N1) standard sera and antigens obtained from the National Institute for Biological Standards and Control (Hertfordshire, United Kingdom).
Immunization of ferrets with 2009 A (H1N1) vaccine antigens and challenge of ferrets with wild-type 2009 A (H1N1) or seasonal H1N1 influenza viruses
Ferrets (n = 4 per group) were purchased from Marshall Farms (North Rose, NY). All ferrets were approximately 7 weeks old and were serologically negative for the human influenza viruses (H1N1, H3N2, and human B) and 2009 influenza A (H1N1) viruses by hemagglutination inhibition (HI) assay. Ferrets were intramuscularly (IM) immunized with one dose or two doses of 7.5 μg of 2009 A (H1N1) vaccine antigens. Ferrets immunized receiving two doses of vaccine antigens were boosted 4 wk after the first immunization. Ferrets were intranasally (IN) infected with 1 mL of 106 log 10 egg infectious dose 50 (log10EID50/mL) of A/California/04/09 (H1N1), A/California/07/09 (H1N1), or A/Brisbane/59/07 (H1N1) at 1 wk after the second vaccination. Unvaccinated ferrets infected with 2009 A (H1N1) influenza virus or naïve ferrets were used as controls. Animal experiments were approved by the Animal Experimental Ethics Committee at Chungnam National University.
Measurement of body temperatures
The rectal body temperatures of the ferrets were measured by inserting a thermometer into the rectum until 10 days after infection.
Clinical scores of ferrets
Clinical scores were the sum of activity and respiratory signs. The activity was graded as follows: 0 representing normal activity, 1 representing calm demeanor, and 2 representing depression and loss of interest in play. The respiratory signs were graded as follows: 0 representing the normal state, 1 representing occasional sneezing and serous nose exudates, and 2 representing frequent sneezing, coughing, and mucosal nose exudates.
Determination of viral titers in nasal washes
Ferrets (n = 4 per group) were immunized with one dose of 7.5 μg of 2009 A (H1N1) vaccine antigens, and 5 wk later were IN infected with 1 mL of 106 log10EID50/mL of A/California/04/09 (H1N1), A/California/07/09 (H1N1), or A/Brisbane/59/07 (H1N1). Unvaccinated ferrets infected with 2009 A (H1N1) influenza were used as controls. The nasal washes were performed by instilling 1 mL of PBS into the nostrils of ferrets 3, 5, or 7 days after infection. The viral titers in the nasal washes were determined by log10EID50/mL.
Determination of viral titers in tissues
The nasal turbinates and lung tissues (0.2 g) of ferrets (n = 4) at 4 d after infection were collected in 1 mL of PBS (pH 7.4) supplemented with 2 × antibiotic-antimycotic solution (Sigma-Aldrich, St. Louis, MO), and were homogenized. The tissue supernatants were serially 10-fold diluted in PBS (pH 7.4) before each diluted sample was inoculated into four 10-day-old embryonated eggs. The presence of virus in the inoculated eggs was determined by hemagglutination assays using 0.5% turkey red blood cells. Viral titers were determined as log10 egg infectious dose50/mL (log10EID50/mL). The detection limit of viruses was 1.0 log10EID50/mL.
Hemagglutination inhibition assay
Sera from vaccinated ferrets were collected and treated with receptor-destroying enzyme (RDE) (Denka Seiken, Tokyo, Japan). RDE-treated sera were serially twofold diluted in PBS (pH. 7.4) in V-bottom 96-well plates, and 8 HA units (25 μL) of A/Califonia/04/09 (H1N1) influenza viruses were added to each well before the plates were incubated for 15 min at room temperature. Turkey red blood cells (0.5%, 50 μL) were then added to each well in the 96-well plates prior to incubation at room temperature for 40 min. The HI titer of each serum was expressed as reciprocal dilutions that completely inhibited hemagglutination.
Detection of antibody subtypes in immunized ferrets
The immunized ferrets were euthanized 5 weeks after immunization with high doses of Zoletil (Virbac Laboratories, Carros, France), exsanguinated, and 0.05-g samples of ferret lung (n = 4) were collected and homogenized in 0.5 mL of PBS (pH 7.4). Sera from the immunized ferrets were treated with RDE. The subtypes of antibodies in tissues or sera of immunized ferrets were determined by enzyme-linked immunosorbent assay (ELISA). The wells of 96-well ELISA plates (Greiner Bio-one) were coated with 100 μL (0.05 g) of purified inactivated A/California/04/09 (H1N1) virus diluted in 1 mL of carbonate-bicarbonate buffer (pH 9.6). To each well, 100 μL of supernatants from the homogenized tissues in PBS (pH 7.4) containing 5% horse serum and 0.05% Tween 20 (PBS-Tween 20) were added. The plates were incubated at room temperature for 1 h and then washed three times with PBS-Tween 20. To each well, 100 μL (1:1000 dilution in PBS-Tween 20) of horseradish peroxidase (HRP)-conjugated goat anti-ferret IgA (Acris Antibodies, Herford, Germany), IgM (Acris Antibodies), or IgG (Novus Biologicals, Littleton, CO) were added. The plates were incubated at room temperature for 1 h and then washed three times with PBS-Tween 20. To each well, 100 μL of ABTS peroxidase substrate (KPL, Gaithersburg, MD) were added. The plates were incubated at room temperature for 30 min, and ABTS peroxidase stop solution (KPL) was added to stop the reaction. Optical density (OD) was read at 405 nm using an ELISA microplate reader (Tecan Systems, Inc., San Jose, CA).
Statistical analysis
The statistical significance was determined by a two-tailed, paired Student's t-test. p Values less than 0.05 (p < 0.05) were regarded as significant.
Results
Antibody response in vaccinated ferrets
We immunized ferrets with two doses of 7.5 μg of 2009 A (H1N1) inactivated vaccine to determine antibody responses. Sera were collected from ferrets immunized with one dose or two doses of vaccine, and HI titers against 2009 A (H1N1) influenza viruses (A/California/04/09 and A/California/07/09), or seasonal H1N1 influenza virus (A/Brisbane/59/07) were determined (Fig. 1). The mean HI titer of sera from one-dose immunized ferrets was less than 10 against A/California/04/09 (H1N1), A/California/07/09 (H1N1), or A/Brisbane/59/07 (H1N1), while that from two-dose immunized ferrets was 64 against A/California/04/09 (H1N1) (p < 0.001), 66 against A/California/07/09 (H1N1) (p < 0.001), and 10 against A/Brisbane/59/07 (H1N1).
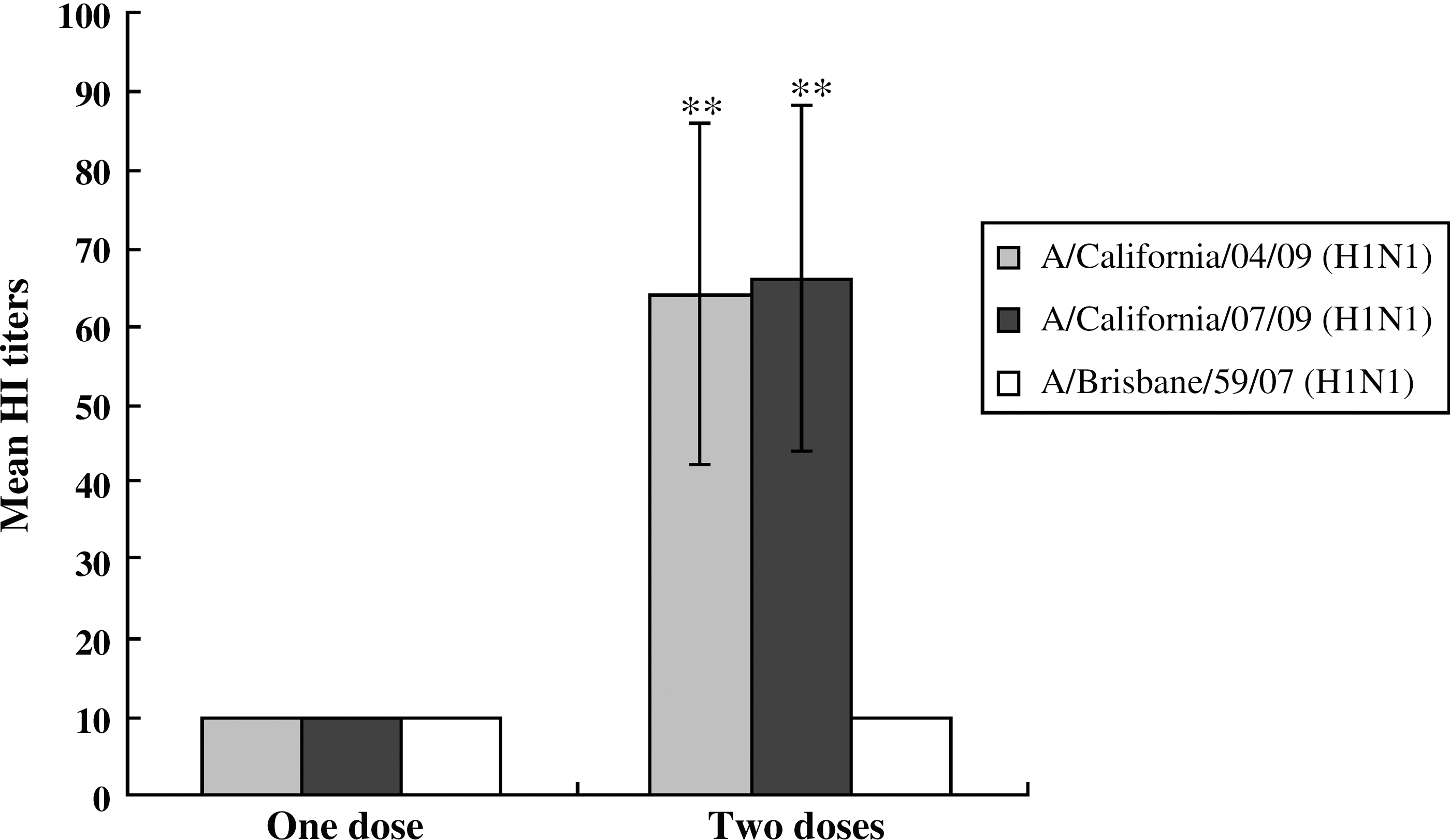
Hemagglutination inhibition titers in sera of immunized ferrets. Sera collected from ferrets (n = 4) immunized with one or two doses of 2009 A (H1N1) inactivated vaccine were treated with RDE. HI titers were determined with A/Califonia/04/09 (H1N1), A/California/07/09 (H1N1), or A/Brisbane/59/07 (H1N1) influenza viruses and 0.5% turkey red blood cells. The data are the mean of 4 ferrets ± standard errors. The statistical analysis was performed between HI titers of one-dose and two-dose immunized ferrets (**p < 0.001).
Protection of immunized ferrets by 2009 A (H1N1) inactivated vaccine
Ferrets immunized with one or two doses (7.5 μg) of 2009 A (H1N1) inactivated vaccine were IN challenged with 2009 A (H1N1) or seasonal H1N1 influenza viruses. The changes in body temperatures, body weights, and clinical signs such as activity and respiratory signs (sneezing, coughing, and mucosal nose exudates) were observed for 10 d after infection. The changes in body temperatures, body weights, clinical scores, and viral titers in tissues of infected ferrets immunized with one dose of vaccine were similar to those of unimmunized and infected ferrets. The mean body temperature of unimmunized ferrets infected with A/California/04/09 (H1N1) was 39.5°C, and that of one-dose immunized ferrets infected with A/California/04/09 (H1N1) was 39.6°C at 4 d after infection. The mean body temperature of unimmunized ferrets infected with A/California/07/09 (H1N1) was 39.4°C, and that of one-dose immunized ferrets infected with A/California/07/09 (H1N1) was 39.8°C at 4 d after infection (data not shown). The mean body temperature of unimmunized ferrets infected with A/Brisbane/59/07 (H1N1) was 38.8°C, and that of one-dose immunized ferrets infected with A/Brisbane/59/07 (H1N1) was 39.2°C (data not shown). When we measured the change in body weight of the ferrets (data not shown), the immunized or unimmunized ferrets infected with A/California/04/09 (H1N1) or A/California/07/09 (H1N1) lost about 7% of their original weight until 6 d after infection, and then started to regain weight. The immunized or unimmunized ferrets infected with A/Brisbane/59/07 (H1N1) lost about 6% of their original weight until 2 d after infection, and then started to regain weight. The unimmunized and uninfected naïve ferrets gained about 7% of their original body weight over 10 d. The clinical scores of the ferrets were recorded until 10 d after infection (data not shown). The immunized or unimmunized ferrets infected with A/California/04/09 (H1N1) or A/California/07/09 (H1N1) had similar clinical scores, ranging from 14 to 16. The immunized or unimmunized ferrets infected with A/Brisbane/59/07 (H1N1) also had similar clinical scores ranging from 8 to 10. Viral titers were determined in nasal turbinates, lungs, and rectal swabs (data not shown). The immunized or unimmunized ferrets infected with A/California/04/09 (H1N1) or A/California/07/09 (H1N1) contained similar viral titers in nasal swabs, lungs, and rectal swabs, with viral titers from 1.75–6.0 log10EID50/mL. The immunized or unimmunized ferrets infected with A/Brisbane/59/07 (H1N1) contained similar titers in nasal turbinates, with viral titers from 6.0–6.5 log10EID50/mL. No virus was detected in the lungs and rectal swabs in the immunized or unimmunized ferrets infected with A/Brisbane/59/07 (H1N1).
When we determined viral-shedding kinetics in the nasal cavities of ferrets immunized with one-dose vaccine, and the viral titers in the immunized ferrets were similar to those in the unimmunized ferrets (Fig. 2). The range of viral titers from 4.5–5.5 log10EID50/mL were detected in the nasal washes of both immunized and unimmunized ferrets 5 d after infection.

Viral titers in nasal washes in one-dose immunized ferrets. Ferrets (n = 4 per group) were immunized with one dose of 2009 A (H1N1) vaccine, and 5 wk later were IN infected with A/California/04/09 (H1N1), A/California/07/09 (H1N1), or A/Brisbane/59/07 (H1N1) virus. The viral titers were determined in the nasal washes collected 3, 5, or 7 days after infection by log10EID50/mL. The data are the mean of 4 ferrets.
We immunized ferrets with two doses of 2009 A (H1N1) vaccine to determine whether the immunized ferrets could be protected from infection by 2009 A (H1N1) or seasonal H1N1 influenza viruses, since one dose of 2009 A (H1N1) vaccine could not protect ferrets from infection by 2009 A (H1N1) or seasonal H1N1 influenza viruses. In contrast to unimmunized ferrets, ferrets immunized with two doses of 2009 A (H1N1) vaccine and infected with 2009 A (H1N1) influenza viruses did not exhibit elevated body temperatures (p < 0.05) (Fig. 3A). At 4 d after infection, the mean body temperature of immunized ferrets infected with A/California/04/09 (H1N1) and A/California/07/09 (H1N1) was 38.2°C, while that of unimmunized ferrets infected with A/California/04/09 (H1N1) and A/California/07/09 (H1N1) were 39.4°C and 39.3°C, respectively. Ferrets immunized with two doses of 2009 A (H1N1) vaccine and infected with seasonal H1N1 influenza viruses had similar body temperatures (Fig. 3A). The mean body temperature of immunized and unimmunized ferrets infected with A/Brisbane/59/07 (H1N1) was 39.0°C, and 38.9°C, respectively. We also determined the change of body weights in ferrets immunized with two doses of vaccine and infected with 2009 A (H1N1) or seasonal H1N1 influenza viruses (Fig. 3B). Ferrets immunized and infected with 2009 A (H1N1) influenza viruses gained weight (p < 0.05), while unimmunized ferrets infected with 2009 A (H1N1) influenza viruses lost weight compared to their original weights before infection (Fig. 3B). At 4 d after infection, the mean body weights of ferrets immunized and infected with A/California/04/09 (H1N1) and A/California/07/09 (H1N1) were 103.9% and 102.9% of the original body weights before infection, respectively, while those of ferrets unimmunized and infected with A/California/04/09 (H1N1) and A/California/07/09 (H1N1) were 94.5% and 94.1% of the original body weights before infection, respectively. At 2 d after infection, the mean body weights of ferrets immunized and infected with A/Brisbane/59/07 (H1N1) were 93.5% of the original weight before infection, and those of unimmunized ferrets infected with A/Brisbane/59/07 (H1N1) were 92.4% of the original weight before infection (Fig. 3B). The clinical scores of the ferrets were also determined. The ferrets immunized with two doses of 2009 A (H1N1) vaccine and infected with 2009 A (H1N1) influenza viruses did not show any clinical signs (p < 0.001), while the unimmunized ferrets infected with 2009 A (H1N1) influenza viruses had clinical scores of up to 16 (Fig. 3C). The clinical scores of the immunized or unimmunized ferrets infected with seasonal H1N1 influenza viruses were similar (Fig. 3C). At 5 d after infection, the clinical scores of the immunized and unimmunized ferrets infected with A/Brisbane/59/07 (H1N1) were 9 and 8, respectively. When we determined the viral titers, viruses were not detected in nasal turbinates, lungs, or rectal swabs of ferrets immunized with two doses of 2009 A (H1N1) vaccine and infected with 2009 A (H1N1) influenza viruses (p < 0.001) (Fig. 3D). Viral titers ranged from 1.75–6.0 log10EID50/mL in nasal turbinates, lungs, and rectal swabs of unimmunized ferrets infected with 2009 A (H1N1) influenza viruses. The viruses were only detected in the turbinates of the immunized or unimmunized ferrets infected with A/Brisbane/59/07 (H1N1), with similar viral titers ranging from 5.5–6.0 (Fig. 3D).

(
To determine which antibody subtypes were induced by vaccination in ferrets, we measured the amounts of antibody in sera and nostril, trachea, lung, heart, liver, spleen, kidney, large intestines, small intestines, and brain tissues of ferrets immunized with one or two doses of vaccine as the OD using plates coated with the inactivated purified A/California/04/09 (H1N1) viruses and HRP-conjugated ferret-specific secondary antibodies. IgG antibodies were predominantly detected in sera and tissues of the immunized ferrets. IgM antibodies were detected in sera (p < 0.001), but not in tissues, of ferrets immunized with one (OD 0.5) or two doses (OD 0.6) of vaccine (Fig. 4A). The OD of IgG antibodies in the lungs of ferrets immunized with one and two doses of vaccine (p < 0.001) were 0.06 and 0.48, respectively (Fig. 4B), and the OD of IgA antibodies were similar in sera and tissues of both immunized and unimmunized ferrets (Fig. 4C). These data suggest that IgG antibodies may be responsible for the protective immunity seen in ferrets immunized with the inactivated 2009 A (H1N1) vaccine.

Determination of antibody subtypes in tissues and sera. Antibody subtypes in tissues and sera from ferrets (n = 4) immunized with one or two doses of 2009 A (H1N1) vaccine were determined with plates coated with the purified inactivated virus of A/California/04/09 (H1N1) and HRP-conjugated goat anti-ferret IgM (
Discussion
Swine-origin pandemic H1N1 influenza viruses are causing some mortality of infected humans around the world. Our study showed that ferrets immunized with two doses of the inactivated vaccine of 2009 A (H1N1) influenza viruses were protected from infections by 2009 A (H1N1) influenza viruses, but not from infections of seasonal H1N1 influenza viruses. IgG subtype antibodies specific for 2009 A (H1N1) influenza viruses were predominantly detected in sera and tissues, including lungs, of immunized ferrets.
Our data showed that two doses of inactivated vaccine of 2009 A (H1N1) influenza viruses were sufficient to protect ferrets from infections by 2009 A (H1N1) influenza viruses. Inactivated influenza virus vaccines are considered a primary tool to prevent infection by influenza viruses (3). The inactivated vaccines of seasonal influenza viruses contain 15 μg of HA of H3N2, H1N1, and B viruses, and are parenterally immunized to induce anti-HA antibodies. In the U.S., manufacturers annually produce over 120 million doses of inactivated influenza vaccine. The immunization rate of the U.S. population is estimated to be 65% of elderly persons and 48% of infants.
Our results showed that ferrets immunized with one or two doses of 2009 A (H1N1) influenza viruses could not be protected from infections by seasonal H1N1 influenza viruses. This suggests that most humans may not possess cross-reactive antibodies against 2009 A (H1N1) influenza viruses, and may need to be vaccinated with at least with two doses of inactivated vaccine for protection from 2009 A (H1N1) influenza viruses. Itoh et al. (12) showed that elderly persons born before 1918, but not persons born in the 1920s to 1950s or since 1977, possess cross-reactive antibodies against 2009 A (H1N1) influenza viruses. This cross-reactivity may be due to the origin of HA of 2009 A (H1N1) influenza viruses, since it was derived from 1918 pandemic virus (H1N1). However, Katz et al. (13) showed that among adults, cross-reactive antibody against 2009 A (H1N1) influenza viruses could be detected in 6–9% of persons aged 18–64 years, and in 33% of persons aged over 60 years. A microneutralization assay also revealed that 4 (4%) of 107 persons born after 1980 contained pre-existing cross-reactive antibody titers of 40 or more against 2009 A (H1N1) influenza viruses, and 39 (34%) of 115 persons born before 1950 had antibody titers of 80 or more against 2009 A (H1N1) influenza viruses (22). A study of pre-existing immunity against swine-origin H1N1 influenza viruses in humans showed that 8 out of 25 of B-cell epitopes present in the seasonal H1N1 influenza viruses were conserved in the swine-origin H1N1 influenza viruses. Among six epitopes present in hemagglutinin (HA) and neuraminidase (NA) of the seasonal H1N1 influenza viruses, one epitope was conserved in the HA and NA of swine-origin H1N1 influenza viruses (8).
Our results showed that immunization with one dose of 2009 A (H1N1) influenza vaccine could not induce HI titers over 40 in the immunized ferrets. Our results are different from those of a study of humans, in which over 70% of humans immunized with one dose of 2009 A (H1N1) influenza vaccine showed HI titers over 40 (2,9,10,19,28). The reason that a one-dose vaccine can induce antibody responses in humans but not in ferrets may be due to the existing cross-reactive memory B cells induced by the seasonal H1N1 influenza viruses. Greenberg et al. (9) reported that about 30% of humans showed HI titers over 40 to 2009 A (H1N1) influenza viruses when sera were collected prior to vaccination. This result suggests that a considerable portion of humans may have memory B cells cross-reactive to 2009 A (H1N1) influenza viruses. Meanwhile, a recent study on the efficacy of commercially available pandemic vaccine in ferrets by Kobinger et al. (16) showed that a single dose of matched inactivated vaccine partially protected ferrets against 2009 A (H1N1) influenza virus.
Our study showed that the protective immunity seen in ferrets immunized with 2009 A (H1N1) vaccine might be mediated by the IgG subtype of antibody. Further study is needed to determine which subclass of IgG antibody is responsible for the protection of immunized ferrets from infection by 2009 A (H1N1) influenza virus. At present, no reagent to differentiate the subclass of ferret IgG antibody is available. Gordon et al. (5) interestingly showed that infected humans with IgG2 deficiency for 2009 A (H1N1) influenza virus suffered from severe clinical signs.
In conclusion, our study using a ferret model suggests that the vaccine of swine-origin 2009 A (H1N1) influenza viruses may protect humans from 2009 A (H1N1) influenza viruses, but not from the seasonal H1N1 influenza viruses.
Footnotes
Acknowledgments
This study was supported in part by a grant (A084411) from the Korea Healthcare Technology R&D Project, Ministry for Health, Welfare & Family Affairs, Republic of Korea. A staff member of HARRISCO, an English editing company, edited this manuscript.
Author Disclosure Statement
No competing financial interests exist.