
Abstract
Chronic hepatitis B is a major health problem, with more than 350 million people infected worldwide. Available therapies have limited efficacy and require long-term continuous and expensive treatments, which often lead to the selection of resistant viral variants and rarely eliminate the virus. Immunotherapies have been investigated as a promising new approach. Several vaccine formulations have been clinically tested in chronic patients, none of which have clearly demonstrated efficacy so far. In this study we evaluated a new vaccination strategy comprising the simultaneous co-administration by the nasal and parenteral routes of a multicomponent vaccine formulation in BALB/C and HBsAg-transgenic mice. The formulation under study contains the surface and nucleocapsid antigens of the HBV, and was co-administered by the nasal route and three parenteral routes. For parenteral administration we also evaluated the immunogenicity of the antigenic mixture with alum or without the adjuvant. The immune response was evaluated by ELISA and IFN-γ ELISPOT assays. Our results indicate that all variants generated a strong antibody response in the sera against both antigens, but differed in their capacity to induce cellular immune responses against the surface antigen. Mice immunized by the nasal and subcutaneous routes without alum generated the highest IFN-γ-secreting CD8+ T-cell response, and results in this transgenic mouse model showed that there is no need to include alum. In conclusion, our results indicate that the immunization routes have to be carefully selected before carrying out clinical trials to optimize the immune response and promote further clinical development.
Introduction
HBV is a unique DNA virus that replicates in hepatocytes; these cells produce and secrete hepatitis B surface antigen (HBsAg), hepatitis B e antigen (HBeAg), and intact virions (HBV-DNA) into the bloodstream (13). The HBsAg and the core antigen (HBcAg) are the main structural antigens of HBV. Both antigens constitute potent immunogens for experimental animals, as well as for patients acutely infected with HBV (23).
Chronic infection with the hepatitis B virus (CHB) is a theoretically appealing and practically feasible target for therapeutic vaccination. However, it is difficult to define a strategy that would be most successful. The results from multiple clinical trials show that more powerful antigen formulations and novel adjuvant strategies are required to overcome the state of unresponsiveness of chronic patients (34,37). Numerous studies in HBV-transgenic mice indicated that HBcAg is the antigen of choice for increasing the number of epitopes and the variety of the immune response, even when it is combined with other antigens such as HBsAg (4,36). These studies also confirm the prominent role of HBcAg as a pro-Th1 antigen (28).
Our group has been working on the development of a therapeutic vaccine candidate based on a combination of HBsAg and HBcAg antigens designed for nasal administration. The rationale of this approach is to activate both parenteral and mucosal immunity, in order to increase the probability of success of the vaccine formulation. This is possible by the mucosal route or by a mucosal/parenteral combination, but not by the parenteral route alone. The present vaccine candidate was developed and clinically tested in a phase I trial with encouraging results (1,2,21). However, the immunogenicity of the formulation could be optimized in humans by adding a parenteral immunization branch that would subvert the strong tolerogenic environment to HBV antigens in chronic patients. We decided to explore combinations of routes in order to develop broader and more potent immune responses (15,16). Reports show that in chronic infections such as HIV, the immunization strategy of combining routes enhanced the resulting immune response (9,22,33). The simultaneous nasal-parenteral administration, at the same dose, of a vaccine formulation against HIV-1 was previously reported by our group (16). We showed that the nasal/subcutaneous co-administration generated a better Th-1 immunodeviation, and an overall improved immune response against HIV antigen over that of a previous study, in which mice were immunized intranasally and subcutaneously independently (priming-boosting) with the same multi-antigenic formulation (17).
In the present study we evaluated a new therapeutic vaccine candidate for chronic hepatitis B therapy in BALB/C and HBsAg-transgenic mice. The evaluation included the simultaneous co-administration, by the nasal and parenteral routes, of a multicomponent vaccine formulation. The formulation, called Nasvac, contains the surface and nucleocapsid antigens of HBV, and was co-administered by the nasal route and three parenteral routes. For the parenteral administrations we also assessed the behavior of the formulation with or without the inclusion of alum, which was used as an adjuvant.
Materials and Methods
Antigens
The recombinant HBsAg subtype adw2 was produced at over 95% purity at the production facilities of the Center for Genetic Engineering and Biotechnology (CIGB, Havana, Cuba), as a component of the commercial HBV prophylactic vaccine Heberbiovac-HB. The HBsAg for this vaccine is expressed and purified from the Pichia pastoris yeast. The entire recombinant rHBcAg particle of 183 amino acids was expressed in Escherichia coli and obtained with a purity of >95%. The formulation containing both antigens was obtained by a simple mixture in phosphate-buffered saline (PBS) (0.1 mol/L NaCl, 2 mmol/L KCl, 10 mmol/L Na2HPO4, and 1 mmol/L KH2PO4 [pH 7.2]). For the groups immunized with alum we used the same formulation adjuvanted with 0.5 mg/mL of alum.
Immunization schedules
BALB/C female mice 8–12 weeks of age were purchased from CENPALAB (Havana, Cuba). Twelve animals per group were inoculated. Group 1 was inoculated with HBcAg + HBsAg IM and IN, group 2 was inoculated SC and IN, and group 3 was inoculated ID and IN. These three groups received the formulation adjuvanted with 0.5 mg/mL of aluminum hydroxide (Superfos Biosector A/S, Vedbaek, Denmark, referred to here as alum) by each parenteral route. Group 4 was immunized with HBcAg + HBsAg IM and IN, group 5 was immunized SC and IN, and group 6 was immunized ID and IN, all without the adjuvant. A seventh group was immunized with 0.5 mg/mL of aluminum hydroxide in PBS (placebo group) IM and IN. In all cases the animals were immunized at the same time by both routes at each dose. The antigens were used at 5 μg per mouse. The immunogens were administered systemically in a final volume of 100 μL. For the nasal route, the proteins were dissolved in sterile PBS. The animals were anesthetized by the IP administration of 30 μL ketamine (10 mg/mL), placed in a supine position, and the immunogens were dispensed slowly in 50 μL (25 μL/nostril) using a pipette tip. In all cases the antigen mixtures were prepared the day before and stored at 4°C until inoculation. The administrations were carried out on days 0 and 14, and the sera were collected 10 d after each dose through the retro-orbital plexus.
For the second set of experiments, a lineage of HBsAg-transgenic mice with a BALB/C genetic background (CIGB, Cuba) (8,10) was used. Mice 8–16 wk old of both sexes were used. Five mice per group were immunized as follows: group 1, HBcAg + HBsAg SC and IN (for this group we subcutaneously administered the formulation adjuvanted by 1 mg/mL of alum); group 2, HBcAg + HBsAg SC and IN; and group 3, not treated. Antigens were used at 5 μg per mouse. The doses were given on days 0, 14, 28, 42, 56, 70, and 84, and blood extractions were carried out on days 38, 66, and 94. As for the BALB/C immunization schedule, the transgenic mice were immunized at the same time by both routes at each dose.
All experiments were conducted following institutional guidelines.
Biological fluids
The animals were bled by retro-orbital puncture and the blood samples were centrifuged at 7800 g for 10 min (centrifuge 5415C; Eppendorf, Hamburg, Germany). Sera were collected and stored at −20°C until evaluation. Vaginal washes were obtained by a skilled technician to avoid trauma and blood contamination. They were obtained by reflushing 100 μL of sterile PBS solution with a micropipette. The volumes collected were centrifuged as above, and the supernatant was stored at −20°C.
Enzyme-linked immunoassay (ELISA) for determining total IgG and IgG subclasses
The specific IgGs against each antigen in the formulation were evaluated by ELISA. Briefly, high-binding plates were coated with 100 μL of the specific antigen at 5 μg/mL in the coating buffer (11 mM Na2CO3 and 35 mM NaHCO3 [pH 9.6]), and incubated overnight at 4°C. The plates were blocked with 2% (w/v) skim milk in PBS (0.1 M NaCl, 2 mM KCl, 10 mM Na2HPO4, and 1 mM KH2PO4 [pH 7.2]) for 1 h at 37°C. Subsequently the plates were incubated with the serum samples diluted with 1% (w/v) skim milk and 1% (v/v) Tween-20 in PBS for 2 h at 37°C. The anti-mouse IgG peroxidase conjugate (Sigma-Aldrich, St. Louis, MO) was incubated for 1 h at 37°C. Subsequently the plates were incubated with the substrate solution (52 mM Na2HPO4, 25 mM citrate, 1 mg/mL OPD, and 0.1% [v/v] H2O2) for 15 min at room temperature. Washes were done with 0.05% (v/v) Tween-20 in PBS solution three to five times between each step. The reaction was stopped with 3 M H2SO4 solution. Finally, the plates were read at 492 nm in a microtiter plate reader (Sensident Scan; Merck Darmstadt, Germany).
The IgG subclasses were evaluated using the kit ISO-2 Mouse Monoclonal Antibody Isotyping Reagents according to the manufacturer's recommendations (Sigma-Aldrich).
The IgA response in vaginal washes was evaluated after the second dose by a similar ELISA assay, using a 1:10 dilution of the serum samples and an anti-mouse IgA peroxidase conjugate 1:5000 (Sigma-Aldrich).
Samples positive for antibody titers were detected using cut-off values of twice the optical density (OD) of negative controls (preimmune serum). Each sample was analyzed using an Excel program that interpolated the OD values on a standard curve consisting of a pool of hyperimmune sera of known titers. This standard curve was included in each individual plate. Finally, the results of total IgG and subclasses were represented as the logarithm of the geometric mean of the titer (GMT) for each treatment group (with a confidence interval of 95%). For the non-seroconverting sera, an arbitrary titer of 1:50 was assigned for statistical processing.
Enzyme-linked immunospot (ELISPOT) assay for determining the interferon-γ (IFN-γ) response
A IFN-γ-based ELISPOT assay was used to detect the sensitization of CD8 T cells by MHC-I-presented synthetic peptides, resulting in the secretion of IFN-γ.
Preparation of target and effector cells
Ten days after the last immunization, spleens were aseptically removed and individual cell suspensions were prepared. Erythrocytes were lysed after 5 min of incubation with 1 mL per spleen of 0.83% (w/v) NH4Cl. The cells were extensively washed with medium, resuspended in RPMI 1640 (Gibco, Carlsbad, CA), and supplemented with 10% (v/v) fetal calf serum (FCS) (Gibco, USA), 2 mM glutamine, 2 mM pyruvate, 50 mM 2-mercaptoethanol, and antibiotics (complete medium), and counted.
For measuring frequencies of responding CD8 T cells, an optimized epitope presentation was achieved by using p815 H-2d mastocytoma cells (American Type Culture Collection TIB-64™) as stimulator cells exogenously loaded for 1 h with a synthetic peptide at saturating concentrations. We employed a concentration of 10 mM of the peptide S28–39 (IPQSLDSWWTSL) from HBsAg (CIGB) (30). After incubation, p815 cells were further incubated for 15 min with mitomycin C (Sigma-Aldrich). They were extensively washed to remove any trace of mitomycin C, and resuspended in the complete medium for counting. p815 Cells without the peptide were also treated as controls.
In vitro re-stimulation of primed CTL
After the washing steps, the cells were counted and distributed in 25-cm2 flasks (Becton Dickinson, Alcester, England) at 2 × 106 cells per milliliter in 10 mL of the complete medium, and stimulated with 5 μg/mL of peptide S28–39. After a 4-d culture at 37°C and 5% CO2, half of the total medium was substituted by a new medium containing 20 IU/mL of IL-2 (CIGB). On day 7 the cells were collected and counted.
ELISPOT assay
Nitrocellulose-backed 96-well MAHA S45 plates (Millipore, Molsheim, France) were coated with 100 μL of 5 μg/mL murine IFN-γ-specific mAb R4-6A2 (Pharmingen, Becton Dickinson, Oxford, England), and left to stand overnight at 4°C. Then they were washed three times with PBS and blocked using the complete medium at 37°C for 1 h. Two dilutions (2 × 105 and 1 × 105) of freshly isolated or re-stimulated splenocytes (104 and 5 × 104), and 1 × 105 p815-pulsed cells with the peptide S28–39, were incubated for 20 h at 37°C in 5% CO2. Splenocytes incubated with 2.5 μg/mL of concanavalin A (ConA; Sigma-Aldrich) were used as positive controls. Each group was controlled by the same number of wells incubated with unpulsed p815 cells as the negative control and the experimental controls of placebo mice.
After 20 h of incubation the plates were washed three times with PBS and five times with PBS-0.05% (v/v) and Tween 20, then 0.5 μg/mL of anti-IFN-γ-biotin conjugated antibody (XMG1.2, Pharmingen, Becton Dickinson) was added and left to react at room temperature for 2 h. The plates were then washed five times with PBS-0.05% (v/v) Tween 20, and peroxidase-labeled streptavidin (Sigma-Aldrich) was added at a 1:1000 dilution for 1 h. The wells were washed again with PBS-0.05% (v/v) Tween-20, and the spots were developed by adding 3,3′-diaminobenzidine (3,3′,4,4′-tetraaminobiphenyl) tetrahydrochloride (Sigma-Aldrich) in 50 mM Tris-HCl (pH 7.4) with 0.3% (v/v) H2O2. After 15 min the wells were washed with tap water, dried, and the spots were counted under a dissection microscope (Carl Zeiss, Jena, Germany). The ELISPOTs were assayed under re-stimulation with the S28–39 peptide using five individual samples per group.
Statistical analysis
GraphPad Prism version 4.00 statistical software (Graph-Pad Software, San Diego, CA) was used for statistical analyses. All titers were transformed into log10 for a normal distribution. The statistical treatment was carried out using the F test to evaluate variance homogeneity, followed by the t-test in for the comparison of two groups. For comparisons of multiple groups the results were analyzed with GraphPad Prism 4.00, with one-way ANOVA and Newman-Keuls testing for parametric tests, or Kruskal-Wallis and Dunn's tests for non-parametric tests. A p value of <0.05 was considered to be statistically significant.
Results
Antigen-specific humoral immune response
HBsAg-specific IgG response in the sera
The HBsAg-specific total IgG response was evaluated in sera for each dose. Ten days after the first dose we did not observe seroconversion in any of the tested sera; this behavior is typical of HBsAg, which needs T-cell cooperation for the production of a specific response in the sera. The IgG response obtained after the second dose was high for all groups (GMT >104) (Fig. 1).
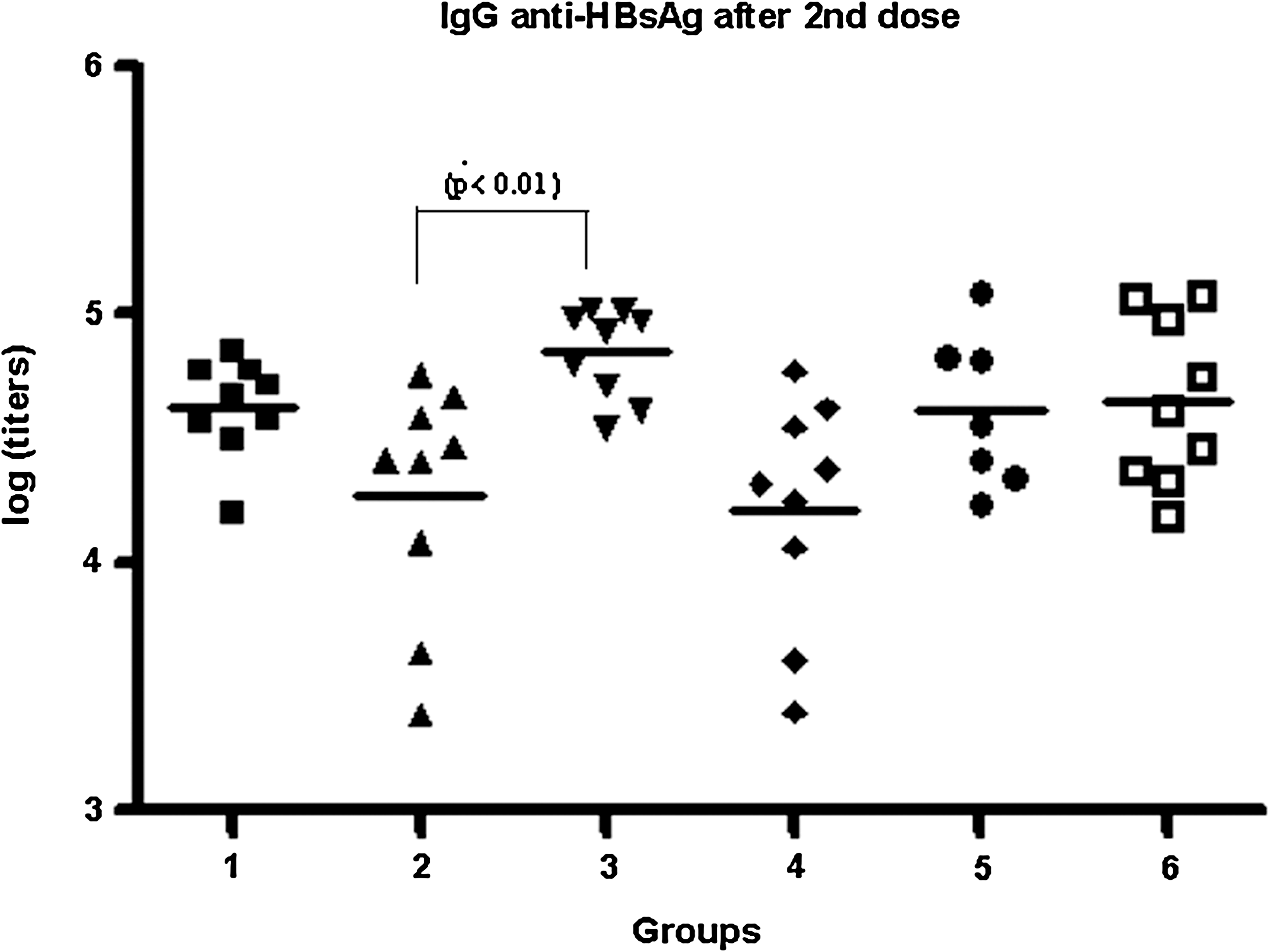
The HBsAg-specific IgG response in sera by ELISA after the second dose. The lines represent the means in each case, and the statistically significant difference observed is indicated. Groups 1, 2, and 3 were immunized with HBcAg + HBsAg IM and IN, SC and IN, and ID and IN, respectively. These three groups received the formulation adjuvanted by 0.5 mg/mL of alum for each parenteral route. Groups 4, 5, and 6 were immunized with HBcAg + HBsAg IM and IN, SC and IN, and ID and IN, respectively, without the adjuvant. The antigens were used at 5 μg per mouse.
In general, we did not observe statistically significant differences in the HBsAg-specific IgG response for the groups immunized with (groups 1, 2, and 3) or without the alum adjuvant (groups 4, 5, and 6) (p > 0.05). After analyzing the influence of the different parenteral routes on the HBs-specific IgG response, we only detected a difference between groups 2 and 3 (p < 0.01). This result indicates that in the presence of alum in the parenterally-administered formulation, the intradermal route yielded better titers than the subcutaneous route. However, in the absence of the adjuvant both routes were similar (p > 0.05) in their capacity to generate HBs-specific IgG responses in the sera.
HBcAg-specific IgG response in the sera
The HBcAg-specific total IgG response was evaluated in the sera after each dose. In contrast with the HBsAg-specific IgG response, 10 d after the first dose we observed seroconversion and high titers (GMT ≥104) in all tested sera. The IgG response obtained after the second dose was similarly high for all groups (GMT ≥105) (data not shown, see online fig. 1).
Response of HBs-specific IgG subclasses in the sera
The specific IgG subclass response was evaluated after the second dose (Fig. 2). In general, the IgG2b titers generated for all groups were similar in magnitude to the IgG1 titers, differing from the IgG2a titers, which were lower.
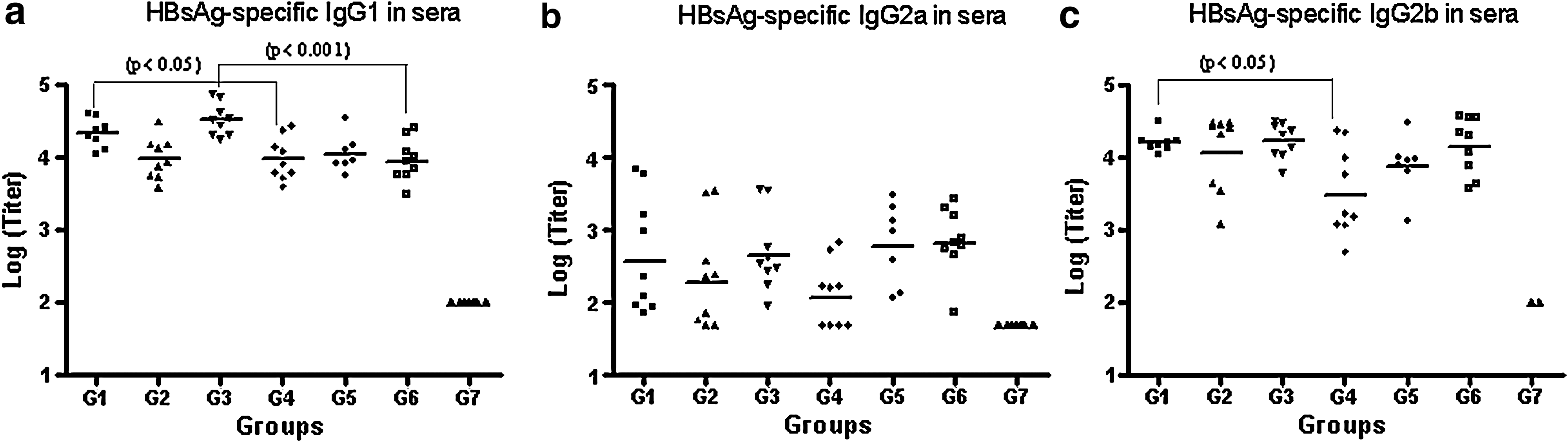
The HBsAg-specific IgG subclass response in the sera by ELISA after the second dose represented as individual subclass titers for IgG1 (

Comparing the groups immunized systemically with or without the alum for IgG1, we only detected statistically significant differences for group 1 and 4 (p < 0.05), and group 3 and 6 (p < 0.001). For IgG2a all the corresponding pairs generated a similar response (p > 0.05), and for IgG2b only groups 1 and 4 differed (p < 0.05).
In analyzing the ratio IgG1/IgG2a, we observed a trend towards generating a Th-2-like antibody pattern for the groups immunized subcutaneously or intradermally with the alum adjuvant compared with the same variant without the alum, and this trend was statistically significant for the intradermal route. This trend was not seen for the groups immunized by the intramuscular route, for which the ratios were similar regardless of the presence of the alum.
On the other hand, the ratios of IgG1/IgG2b were similar for all groups, with the exception of group 4, which showed a marked Th-2-like pattern even in the absence of alum, as mentioned above.
The HBsAg-specific IgA response in vaginal washes
Given that intranasal immunization generated humoral responses in mucosal compartments, the HBs-specific IgA response in vaginal washes was evaluated after the second dose. Fig. 3 shows the results expressed as optical density (OD) at 492 nm for each individual mouse. All groups showed a similar IgA response (p > 0.05), although trends toward generation of higher responses were observed for the groups immunized SC/IN and ID/IN with the parenteral formulation adjuvanted by alum.

The HBsAg-specific IgA response in vaginal lavage fluid by ELISA after the second dose. All of the groups showed a similar IgA response (p > 0.05; OD, optical density). The lines represent the means. Groups 1, 2, and 3 were immunized with HBcAg + HBsAg IM and IN, SC and IN, and ID and IN, respectively. These three groups received the formulation adjuvanted by 0.5 mg/mL of alum for each parenteral route. Groups 4, 5, and 6 were immunized with HBcAg + HBsAg IM and IN, SC and IN, and ID and IN, respectively, without the adjuvant. The antigens were used at 5 μg per mouse. A seventh group was immunized with 0.5 mg/mL of alum in phosphate-buffered saline (PBS, placebo) IM and IN.
The HBsAg-specific IFN-γ secretion response in the spleen
Ten days after the first dose, we evaluated the HBsAg-specific IFN-γ secretion response by spleen cells, predominantly by CD8+ T cells. We carried out a 7-day re-stimulated ELISPOT assay as described above, and the results are shown in Fig. 4. The response generated by the group immunized SC/IN without the alum was statistically significantly higher than the corresponding group with alum (p = 0.007), and higher than that seen in the rest of the groups. The response generated by the groups immunized ID/IN with and without the alum did not differ significantly. Under our experimental conditions, in group 4 only 1/5 animals had a positive response, compared to group 1, in which 5/5 responded positively.
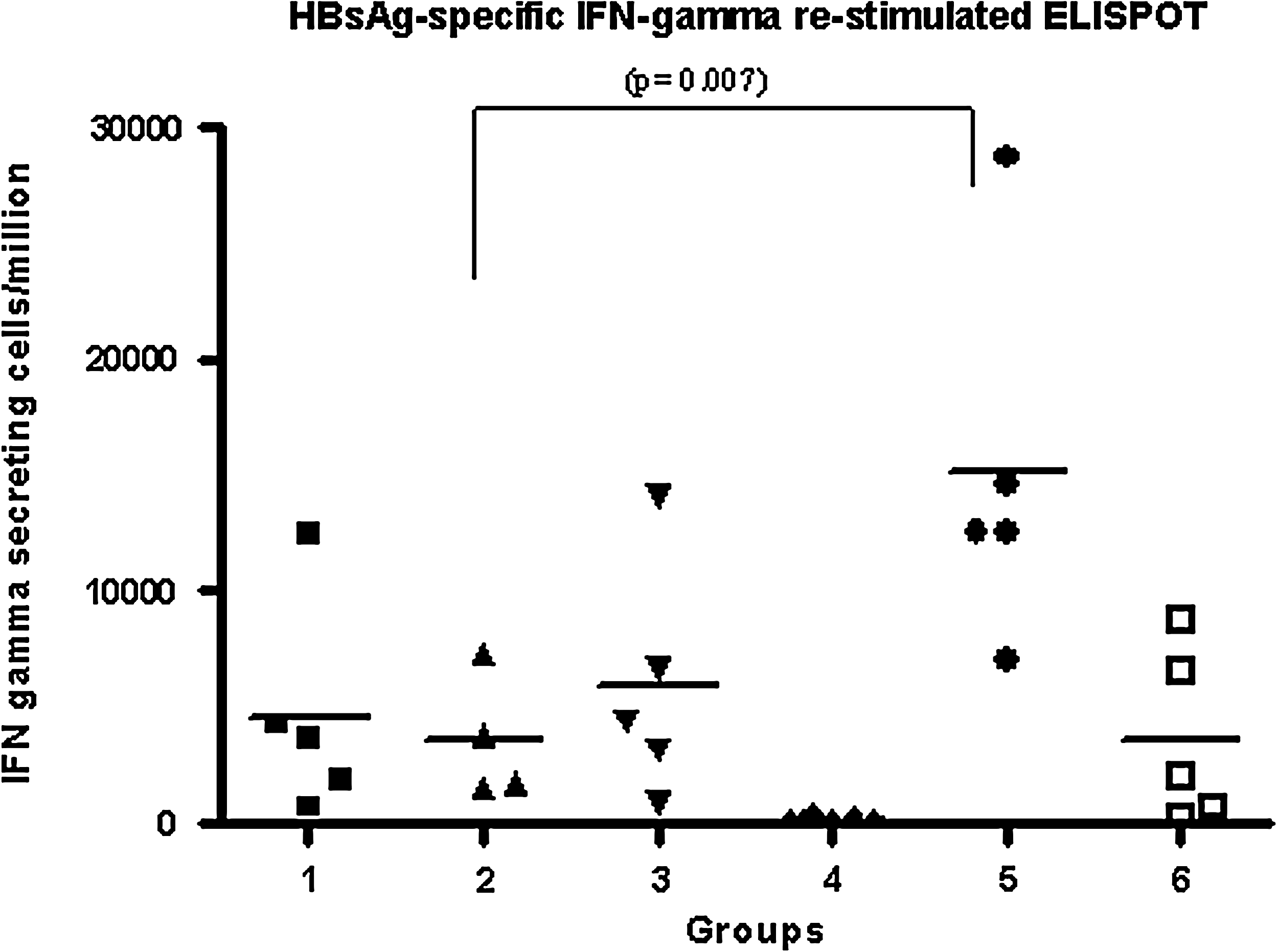
Frequency of IFN-γ-secreting cells as measured by ELISPOT assay after the first dose. The spleen cells were re-stimulated for 7 d with the S28–39 CTL peptide from HBsAg. Groups 1, 2, and 3 were immunized with HBcAg + HBsAg IM and IN, SC and IN, and ID and IN, respectively. These three groups received the formulation adjuvanted by 0.5 mg/mL of alum by each parenteral route. Groups 4, 5, and 6 were immunized with HBcAg + HBsAg IM and IN, SC and IN, and ID and IN, respectively, without the adjuvant. The antigens were used at 5 μg per mouse. In this assay we used cells from each individual animal incubated with concanavalin A (ConA) as the positive control, and cells incubated with RPMI culture medium as the negative control. The lines represent the means and the statistical difference observed is indicated.
HBsAg-specific IgG response in the sera of HBsAg-transgenic mice
In light of the results obtained in BALB/C mice, we decided to evaluate the subcutaneous/intranasal variant in the HBsAg-transgenic mouse model. Five mice per group were immunized, co-administering the formulation intranasally and subcutaneously with or without the alum (groups 1 and 2, respectively). The anti-HBsAg IgG response in the sera was evaluated after the third, fifth, and seventh doses, and the results are shown in Fig. 5. After the third dose we observed no seroconversion in any of the tested sera. After the fifth dose, 3/5 mice from the group immunized with the formulation without the alum generated a detectable IgG response, differing from the group immunized with the formulation without the alum, in which only 1/5 mice seroconverted. The evaluation after the seventh dose showed that both immunized groups generated a similar IgG response. Despite having attained a moderate intensity HBsAg-specific IgG response in both groups, this humoral response was not effective in reducing the amount of HBsAg transgene circulating in the sera (data not shown).
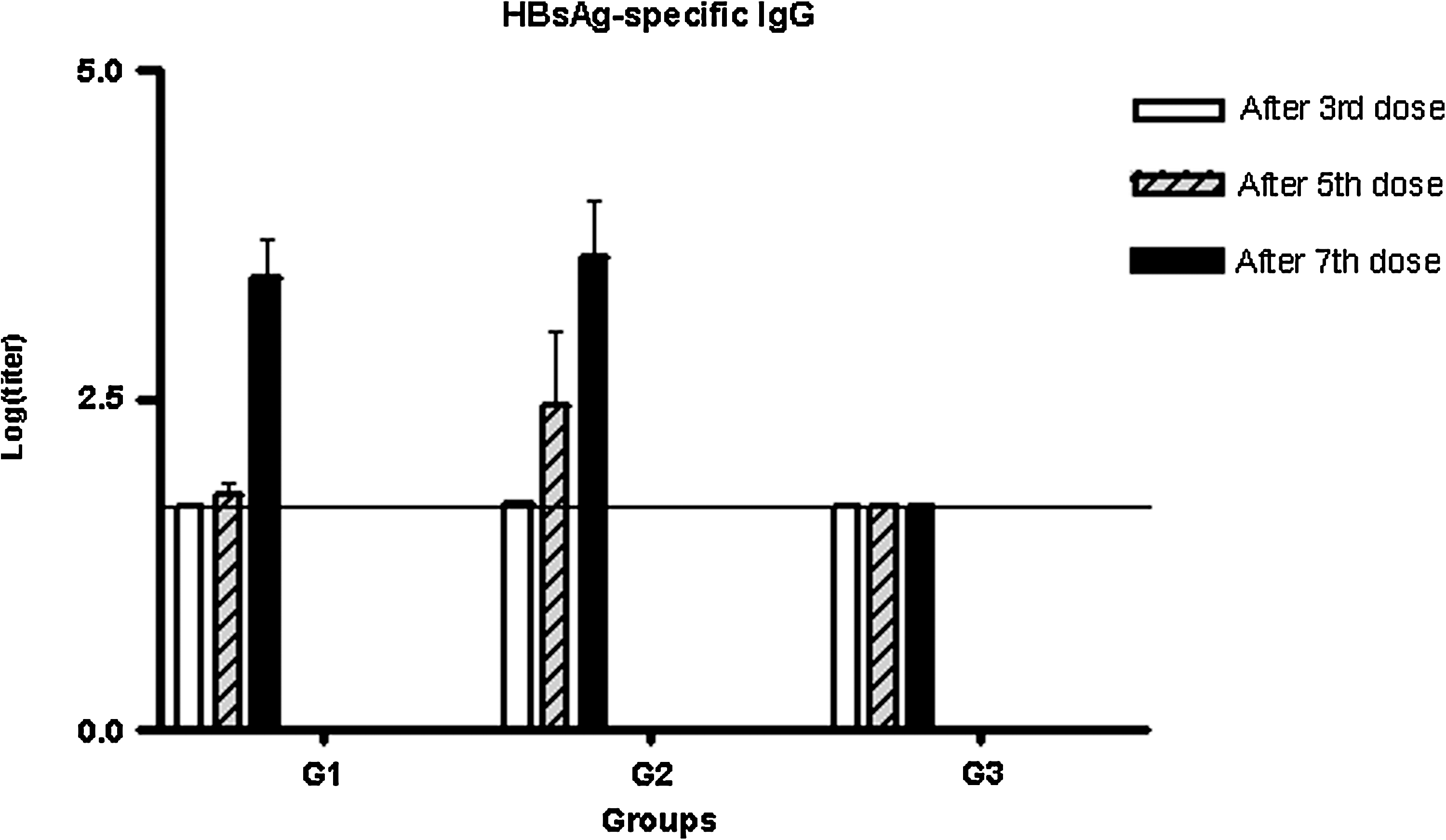
The HBsAg-specific IgG response in sera by ELISA after three, five, and seven doses. Groups of five mice were immunized with HBcAg + HBsAg SC and IN (G1) with the formulation adjuvanted by 1 mg/mL of alum, with HBcAg + HBsAg SC and IN with the formulation without the adjuvant (G2), and were left untreated (G3). The antigens were used at 5 μg per mouse. The error bars represent the standard error of the mean in each case.
Discussion
We evaluated a new therapeutic vaccine candidate for CHB therapy based on HBV surface and core antigens in BALB/C and HBsAg-transgenic mice after the simultaneous co-administration by the nasal and parenteral (IM, SC, and ID) routes. A strong antibody response against both antigens was obtained for all groups, but differences in the induction of the cellular immune response against the surface antigen were found.
The capacity to generate potent polyclonal and multispecific cellular immune responses along with strong antibody responses are important features when selecting a therapeutic vaccine candidate for HBV control during CHB infection (4,36). Furthermore, the seroconversion to anti-HBsAg is regarded as the ideal end-point in CHB therapy (18).
Even though the IgG antibody response for all groups can be considered high, the comparison between groups in terms of IgG levels showed differences. There was a superiority of IN/ID with alum (group 3) over IN/SC without alum (group 2) and IN/IM (group 4); however, group 3 titers were similar to those of the rest of the groups, including the two groups without alum (groups 5 and 6). In the setting of therapeutic vaccination, groups 5 and 6 were more desirable based on the potential need for multiple administrations. Alum has been associated with several adverse reactions (11,27), and even though they are milder compared than oil-based adjuvants, this could be a concern in cases requiring 5, 10, or more administrations, as is current clinical research practice in therapeutic vaccination (14,34).
It has been reported that the HBcAg included in the formulation is a strong Th-1-promoting immunogen, with better immunogenic properties than the co-administered HBsAg (28,29,35). The formulation of HBcAg and HBsAg in PBS (Nasvac) has shown a vigorous response against both antigens, deviating from the pattern of an HBsAg-specific immune response to Th-1 (1,21). In our study strong serum IgG1 and IgG2b HBsAg-specific responses were seen in all study groups, and a less intense IgG2a response was seen. According to the IgG1/IgG2a ratio, the presence of alum in the parenteral formulation favored a Th-2-like pattern of antibodies in the case of ID/IN administration. IM administration also appears to reinforce the Th-2-like pattern, regardless of the presence of alum, compared to the other parenteral groups. In general, the groups administered IN/SC and IN/ID without alum (groups 5 and 6, respectively) showed the most Th-1-like patterns, in agreement with previous results (16). It should be also considered that the humoral response measured in the serum was the result of simultaneous nasal and parenteral administration, and that the formulation administered by the nasal route (HBsAg plus HBcAg in PBS) promoted a Th-1-like IgG pattern of response against HBsAg, as was previously reported (1,21).
Taking into account that the intranasal route of administration could induce a secretory IgA response in mucosa, even in remote areas such as the genital tract (24), we evaluated the HBs-specific IgA response in vaginal lavage fluid. A similar mucosal immune response was generated in all groups, although there was a tendency for it to be higher in the groups receiving the parenteral formulation with the alum adjuvant, especially in the subcutaneous and intradermal groups. The generation of a specific humoral response in mucosal tissues could be effective in the control of viruses that are found in these tissues and their secretions (42). One possible disadvantage of current therapeutic vaccinations may be that they focus on stimulating systemic immunity rather than utilizing mucosal immune system resources, which may be less affected by chronic infections.
The role of potent cellular immune responses in HBV clearance in chronically infected patients has been consistently demonstrated (5,36). The ability to achieve clearance by the transfer of bone marrow from a naturally HBV-immune donor, and the fact that chronically infected patients recovering from infection develop cytotoxic T lymphocyte (CTL) responses similar to those of acute patients, further support this effect (7,18). The concept of cytokine-mediated HBV clearance has been also widely accepted (6,38).
In this study we explored the cellular immune response using an ELISPOT assay that can measure the frequency of IFN-γ-secreting CD8+ T cells, which are specific for the Ld-restricted HBV surface 28-39 CTL epitope. The ELISPOT assay showed differences in the frequencies of IFN-γ-secreting CD8+ T cells between groups. The three groups receiving the formulation with alum adjuvant generated a similar response, which differed from that seen in the three groups immunized with the variant without alum. The most vigorous response was obtained in the group immunized via SC/IN without alum, and this response was significantly higher than that seen in the equivalent IN/SC group with alum adjuvant. A lower IFN-γ-secreting response was elicited in the group immunized IN/IM without alum, corresponding to the marked Th-2-like pattern observed for that group at the IgG subclass level.
Taking into account these results, we selected the SC/IN variant to test the need for the adjuvant in an HBsAg-transgenic mouse model. Our results showed that only after five doses of this formulation were we able to obtain a detectable HBs-specific IgG response in the sera of HBsAg-transgenic mice. Interestingly, the group immunized with the variant without alum was more efficient in generating a specific humoral response, although after the seventh dose both vaccinated groups showed similar IgG responses. These results are encouraging in light of several reports showing that the induction of immune responses by vaccination is difficult in HBsAg-transgenic mice. It has been reported that vaccination techniques that efficiently primed the humoral and CTL responses with Th-1 or Th-2 polarization profiles to HBsAg in H-2d and H-2b mice have been inefficient in HBs-transgenic mice (31,32,39,40).
In this study the HBs-specific humoral immune response generated in HBsAg-transgenic mice was not able to decrease the amount of HBsAg circulating in the sera. It is important to mention at this point that the transgenic mice used in the study have a high HBsAg expression level, with a circulating concentration in the serum ranging from 5 to 20 μg/mL. This feature could affect the quality and quantity of the generated humoral response on the circulating transgenic antigen. Decreasing or suppressing HBsAg antigenemia has proven to be difficult to attain in HBsAg-transgenic mice (31). We have preliminary data demonstrating that the adoptive transfer of spleen cells from BALB/C mice immunized SC/IN without alum (group 5 of the BALB/C groups described above) significantly reduced HBsAg antigenemia in this transgenic model in a transient way.
Other studies in which transgenic mice are administered more doses to evaluate the possibility of generating a cellular immune response are in progress. Future evaluations in transgenic mice expressing the complete HBV genome would also be useful.
To date there is one report (16) documenting the nasal/parenteral co-administration of a similar formulation containing an HIV-1 chimera antigen and the HBV antigens as adjuvants. That article focused on the evaluation of the immune response against the HIV antigen. Immunological data specific to HBV antigens are in accord with the present results.
In general, these results suggest that the proposed vaccination strategy could be promising as a new therapeutic approach for chronic hepatitis B, by the simultaneous stimulation of two different parts of the immune system in order to subvert the tolerogenic status generated by chronic HBV infection. As reported previously, the mucosal immune system has distinctive features, and nasal administration enables antigens to access specialized mechanisms for antigen sampling, including antigen uptake by M cells (25). M cells transport antigens from the luminal surface through a thin layer of cytoplasm to a pocket at the basal surface. M-cell pockets enable the interaction of the antigen with the cells of the immune system in a compartment protected from the modulatory effect of systemic immunity (25). Nasal administration also enables the interaction of the antigens with dendritic cells (DCs) in the tonsils, where professional antigen-presenting cells (APCs) are organized in a surface network of approximately 500 DCs/mm2 (25). This last feature is very important, considering that chronic HBV infection has been associated with functional defects in dendritic cell populations (3,19,41).
We suggest that simultaneous vaccine co-stimulation of mucosal and systemic immunity may circumvent the systemic tolerance established by chronic HBV infection. Because of the difficulties demonstrated in several HBV therapeutic vaccine approaches in clinical trials (34), more aggressive strategies are required.
Footnotes
Acknowledgment
This study was supported by the Center for Genetic Engineering and Biotechnology, Havana, Cuba.
Author Disclosure Statement
All of the authors are employees of the Center for Genetic Engineering and Biotechnology.