
Abstract
The immunogenicity and efficacy of β-propiolactone (BPL) inactivated whole virion SARS-CoV (WI-SARS) vaccine was evaluated in BALB/c mice and golden Syrian hamsters. The vaccine preparation was tested with or without adjuvants. Adjuvant Systems AS01B and AS03A were selected and tested for their capacity to elicit high humoral and cellular immune responses to WI-SARS vaccine. We evaluated the effect of vaccine dose and each adjuvant on immunogenicity and efficacy in mice, and the effect of vaccine dose with or without the AS01B adjuvant on the immunogenicity and efficacy in hamsters. Efficacy was evaluated by challenge with wild-type virus at early and late time points (4 and 18 wk post-vaccination). A single dose of vaccine with or without adjuvant was poorly immunogenic in mice; a second dose resulted in a significant boost in antibody levels, even in the absence of adjuvant. The use of adjuvants resulted in higher antibody titers, with the AS01B-adjuvanted vaccine being slightly more immunogenic than the AS03A-adjuvanted vaccine. Two doses of WI-SARS with and without Adjuvant Systems were highly efficacious in mice. In hamsters, two doses of WI-SARS with and without AS01B were immunogenic, and two doses of 2 μg of WI-SARS with and without the adjuvant provided complete protection from early challenge. Although antibody titers had declined in all groups of vaccinated hamsters 18 wk after the second dose, the vaccinated hamsters were still partially protected from wild-type virus challenge. Vaccine with adjuvant provided better protection than non-adjuvanted WI-SARS vaccine at this later time point. Enhanced disease was not observed in the lungs or liver of hamsters following SARS-CoV challenge, regardless of the level of serum neutralizing antibodies.
Introduction
The mouse is a reasonable model for screening candidate SARS vaccines because SARS-CoV replicates to high titers in the respiratory tract of mice following intranasal inoculation (14). Furthermore, mice develop a neutralizing antibody response that confers protection against subsequent challenge (14). However, young mice do not develop clinical illness, pneumonitis is transient, and the virus is cleared by about day 5 post-challenge (14). Therefore, the efficacy of a SARS vaccine can only be judged by quantitative virology in this model.
Hamsters are an excellent model for SARS-CoV infection and vaccine efficacy since they support high levels of viral replication and histopathological changes in respiratory tissues, and demonstrate clinical signs (reduced activity) following intranasal inoculation (6,11,15). Efficient replication of SARS-CoV in the respiratory tract following intranasal inoculation of golden Syrian hamsters leads to high virus titers in the lungs from days 1 through 7 post-infection (p.i.), with peak titers occurring at day 3 p.i. Pronounced pneumonitis accompanies SARS-CoV infection on days 3–5 p.i., and consolidation of up to one-third of the lungs occurs at days 5–7 p.i. Viral replication and pulmonary pathology coincide with greatly reduced physical activity of the hamsters (16). Thus the efficacy of a SARS vaccine can be judged by quantitative virology and histopathological changes in the lungs of hamsters.
In this study, we evaluated the immunogenicity and efficacy of a
Materials and Methods
Viruses and cells
SARS-CoV (Urbani strain 200300592) was obtained from the Centers for Disease Control and Prevention, Atlanta, GA. Virus stock was produced in Vero cells (CCL81, obtained from ATCC) in serum-free conditions using OptiPRO medium (Invitrogen, Paisley, Scotland). For vaccine preparation, Vero cells were grown in serum-free conditions and infected with a dilution of 1:100 (v:v) of the SARS-CoV virus stock in 120 mL of OptiPRO medium per tray of cells (Nunc, Roskilde, Denmark). The cells were incubated at 37°C for 72 h. At 3 days p.i., when the cytopathic effect (CPE) was visible, the virus was harvested by collecting cell supernatants. The infectious titer of the virus was determined using a 50% cell culture infectious dose (CCID50) method.
Purification and inactivation of SARS-CoV
Vaccine purification was then performed. After low-speed centrifugation to clarify the cell harvest, SARS-CoV was chemically inactivated with BPL (1:1000 v:v) and incubated for 24 h at 4°C. This was followed by a second incubation for 24 h at room temperature to hydrolyze residual BPL. Virus inactivation was documented by titration of samples before and after inactivation, as well as after completion of purification and concentration of the vaccine. Following BPL inactivation, a polyethylene glycol-sodium chloride (PEG-NaCl) mixture was added to precipitate the inactivated virus. After centrifugation the virus pellets were resuspended in NaCl-Tris-EDTA (NTE) buffer (pH 7.5) before loading onto a 5–45% sucrose gradient. The virus preparation was ultracentrifuged and selected sucrose gradient fractions were collected for further purification. SARS-CoV was recovered in 28% sucrose (w:v) in 150 mM NaCl, 50 mM Tris, and 10 mM EDTA (pH 7.5).
Additional BPL (1:10000 v:v) was finally added to the pool as a bacteriostatic agent. The total protein content as determined by the Lowry method was 467 μg/mL. The relative concentration of S protein was determined by densitometry scanning of Coomassie-stained or silver-stained SDS-PAGE gels. The vaccine was formulated in 100-μL single-dose vials of WI-SARS containing the equivalent of either 0.5, 1.0, or 1.5 μg of S protein. The final formulations were prepared shortly before vaccination by addition of 50 μL PBS or 50 μL adjuvant (GlaxoSmithKline Biologicals, Rixensart, Belgium) to 50 μL of WI-SARS preparation, containing approximately 10% SARS S protein. The Adjuvant System AS01B contained 5 μg 3-O-desacyl-4-monophosphoryl lipid A (MPL), and 5 μg highly purified saponin, Quillaja saponaria Molina (QS21) in 50 μL liposomes. The Adjuvant System AS03A was an oil-in-water-based emulsion. The oil phase contained DL-α-tocopherol and squalene, and the aqueous phase contained the non-ionic detergent polysorbate 80 (Tween 80).
Inactivation of virus infectivity following BPL treatment was confirmed by assaying the infectivity of a sample in Vero cells. CPE was not observed after two passages. In addition, quantitative PCR (Q-PCR) performed at several time points during passage confirmed the absence of amplification of virus genomes (17).
The concentration of viral S protein was determined to be 46 μg/mL, or 10% of the total protein concentration. The concentration of Vero host cell proteins (HCP) in the final material was determined by ELISA and Western blot analysis using serial dilutions of Vero HCP-containing lysates, and was found to be 21% of the total protein concentration of the sample.
Evaluation in BALB/c mice
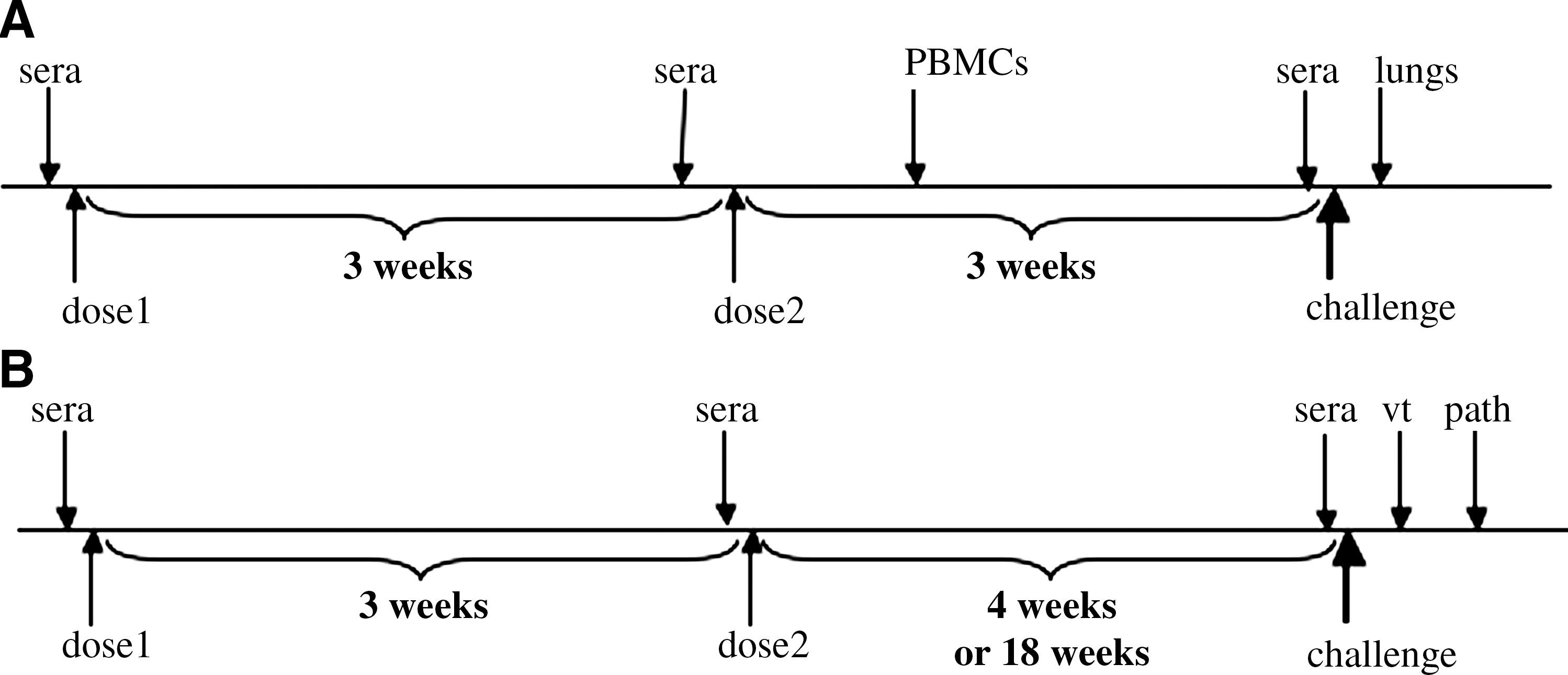
Groups of 8 mice were immunized intramuscularly (IM) on days 0 and 21 with WI-SARS at doses of 0.5 μg, 1.0 μg, and 1.5 μg SARS S-protein in the absence of adjuvant, or with the addition of either AS01B or AS03A adjuvant. A group of mice infected intranasally (IN) with 105 TCID50 of SARS-CoV in a volume of 50 μL served as a positive control. SARS-CoV is not lethal for young BALB/c mice; the virus replicates in the respiratory tract of mice, but is cleared by day 5 p.i. Groups of mice immunized IM with PBS or a BPL-inactivated influenza A virus (Flu-BPL1.5 μg total hemagglutinin protein) prepared in Vero cells, and administered with or without adjuvant AS01B or AS03A were included as negative controls. Sera were collected for determination of SARS-CoV-specific neutralizing antibody titers prior to the first immunization (pre-bleed), the second immunization, and the challenge. The schedule for murine immunizations, challenge, and collection of samples is shown in Fig. 1A.
(
Three weeks after the second dose of vaccine, the mice were lightly anesthetized and challenged IN with 105 TCID50 SARS-CoV. Two days later the mice were euthanized and the lungs were collected and processed for determination of virus titer.
Evaluation in hamsters
Groups of 14 4- to 8-week-old female golden Syrian hamsters were immunized IM on days 0 and 21 with WI-SARS using doses of 0.5 μg or 2.0 μg SARS S-protein (∼5 or 20 μg of total protein) in the absence of adjuvant, or with the addition of AS01B adjuvant. Adjuvant AS03A was not included in the hamster studies because the mouse experiments revealed higher antibody responses with AS01B. Additional control groups of hamsters were infected IN with SARS-CoV (103 TCID50/hamster), or immunized IM with AS01B-adjuvanted Flu-BPL (∼25 μg total protein per dose). Sera were collected for determination of SARS-CoV-specific neutralizing antibody titers prior to the first immunization (naïve hamsters, pre-bleed), prior to the second immunization, and prior to challenge. In order to measure the efficacy of the vaccine against early and late challenge, each group of 14 hamsters was subdivided into two groups (n = 7 each), one for early challenge and the other for late challenge (4 or 18 wk following the second immunization, respectively). For challenge infection, the hamsters were lightly anesthetized and IN inoculated with 105 TCID50 SARS-CoV in 100 μL. Two days after challenge, when peak virus titers were anticipated in the lungs of SARS-CoV-infected hamsters, 4 hamsters in each group were euthanized and the lungs were collected and processed for quantitative virology. Another group of 3 hamsters was sacrificed for histopathological examination on day 5 following challenge, when pulmonary consolidation occurs in SARS-CoV-infected hamsters (15,18). Selected lung sections were stained for SARS-CoV antigen by immunohistochemistry (14). The schedule for hamster immunization, challenge and collection of samples is shown in Fig. 1B.
Microneutralization assay
Twofold dilutions of heat-inactivated serum were tested in a microneutralization assay using 100 TCID50 of SARS-CoV, as previously described (14). The presence of viral cytopathic effect was read on days 3 and 4.
T cell responses by intracellular cytokine staining (ICS)
The T-cell responses as measured by ICS were determined using an adaptation of the method described by Suni et al. (19). PBMC samples obtained 7 d post-immunization were stimulated in vitro with 1 μg/mL of WI-SARS for 2 h and cultured overnight in the presence of brefeldin A (1 μg/mL). The cells were then stained by conventional methods (19,20), and analyzed by flow cytometry to measure the frequency of IL-2- and/or IFN-secreting CD4 T cells. CD8 T-cell responses were below the limit of detection in this experiment.
Results
Immunogenicity in mice
A single dose of WI-SARS vaccine was poorly immunogenic; a low titer serum neutralizing antibody response was detected in mice that received either the intermediate- or high-dose WI-SARS vaccine, containing the equivalent of 0.5 or 1.5 μg of S-protein with AS03A (Fig. 2). A second dose of vaccine induced a significant boost in neutralizing antibody titers at all dose levels, with or without adjuvants (Fig. 2). The antibody response in mice that received WI-SARS with AS01B was greater than that seen in mice that received WI-SARS and AS03A, and was significantly greater than that seen in mice that received WI-SARS vaccine alone (the 1.5-μg dose only; p ≤ 0.05; Fig. 2). All of the vaccinated animals had higher serum neutralizing antibody titers than mice that had recovered from infection with 105 TCID50 of SARS-CoV, although this study and previous studies in mice and hamsters have shown that low levels of neutralizing antibodies are sufficient to prevent detectable viral replication following challenge (12,21). A CD8 T-cell response to SARS-CoV was not detected in PBMCs in any group. Although the spleens were not collected in this experiment, SARS-specific CD8 T cells were previously detected in spleens of mice immunized with AS01B- and AS03A-adjuvanted WI-SARS vaccine, with a higher cellular response observed with AS01B than with AS03A adjuvant (data not shown). Mice that had received adjuvanted WI-SARS developed detectable CD4 T-cell responses. The CD4 T-cell response was significantly higher with AS03A-adjuvanted WI-SARS vaccine than with the WI-SARS vaccine without adjuvant for the doses of 0.5 and 1.5 μg of S-protein (p ≤ 0.001; Fig. 3). A trend toward a higher cellular immune response was observed with AS03A than AS01B adjuvant (p > 0.05), and was optimal with a WI-SARS antigen dose of 0.5 μg (Fig. 3).
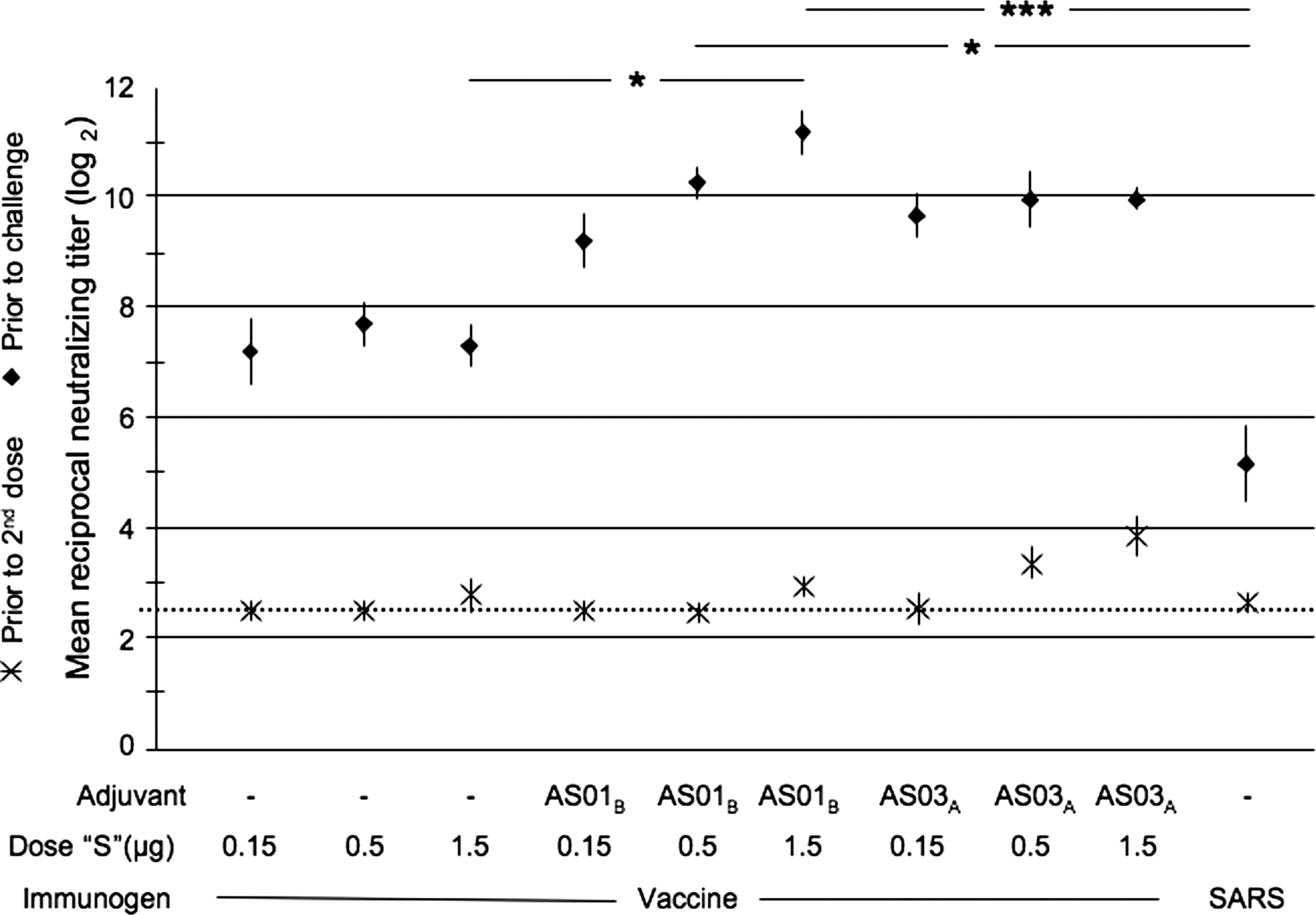
Serum neutralizing antibody responses to WI-SARS vaccine in BALB/c mice. Vaccine dose and adjuvant are indicated along the x-axis; the dose listed is the amount of SARS-CoV S-protein (“S”) administered to each mouse. A control group (SARS) was infected IN with SARS-CoV. Neutralizing antibody titers are expressed as reciprocal mean titer (log2) for groups of 5 mice. Data points show mean titers achieved following one and two doses of vaccine as measured prior to the administration of dose 2 and prior to challenge, respectively. Error bars indicate standard error of the mean. The dotted line indicates the lower limit of detection. Kruskal-Wallis testing with Tukey post-test demonstrated statistically significant differences between the groups indicated, prior to challenge (*p ≤ 0.05; ***p ≤ 0.001).

Cellular immune responses in mice. Shown are percentages of WI-SARS-specific CD4+ T-cell responses (geometric mean titer ± CI 95%) in PBMCs obtained on day 7 post-immunization, and re-stimulated with 1 μg/mL WI-SARS (based on S-protein content). Vaccine dose and adjuvant are indicated along the x-axis. The dose indicates the amount of SARS-CoV S-protein (“S”) as part of the WI-SARS material administered to each mouse. One-way ANOVA with Tukey post-test demonstrated statistically significant differences between the groups indicated (*p ≤ 0.05).
Efficacy in mice
WI-SARS vaccine was highly efficacious in mice at all dosage levels, with or without adjuvants. The challenge virus replicated to titers of 107.5–108 TCID50/g in the lungs of mice that received Flu-BPL with and without adjuvant or PBS. The mean virus titer in the lungs of mice that had recovered from prior infection with SARS-CoV was 102 TCID50/g, corresponding to a 100,000-fold reduction in titer compared to mice that received PBS or influenza vaccine. Replication of the challenge virus was not detected in the lungs of mice that received WI-SARS alone or WI-SARS + AS01B or AS03A (Fig. 4).
Efficacy of WI-SARS vaccine in BALB/c mice. Shown are the mean virus titers in lungs 2 d after challenge with 105 TCID50/mouse SARS-CoV. Vaccine dose and adjuvant are indicated along the x-axis. The dose listed is the amount of SARS-CoV-S-protein (“S”) administered to each mouse. Control groups included mice immunized with a Vero cell-grown BPL-inactivated influenza virus vaccine (Flu-BPL), adjuvants AS01B and AS03A alone, PBS, and primary infection with SARS-CoV. For the schedule of immunization and challenge see Fig. 1. The dashed line indicates the limit of detection (101.5 TCID50/g). Error bars indicate standard error of the mean (n = 5 per group).
Immunogenicity in hamsters
The adjuvant AS03A was not included in the hamster study because higher antibody responses were seen with AS01B adjuvant in the mouse study.
Three weeks following a single dose of vaccine, hamsters that received 2 μg WI-SARS + AS01B had a higher serum neutralizing antibody response than the groups that received WI-SARS alone or 0.5 μg WI-SARS + AS01B adjuvant. However, the neutralizing antibody titer was modest and was lower than the response seen in hamsters that had recovered from prior infection with SARS-CoV (Fig. 5). Administration of a second dose of vaccine induced a significant boost in serum neutralizing antibody titer in all groups. The group that received 2 μg WI-SARS + AS01B had an antibody titer equivalent to that of hamsters that had recovered from prior infection with SARS-CoV (Fig. 5).
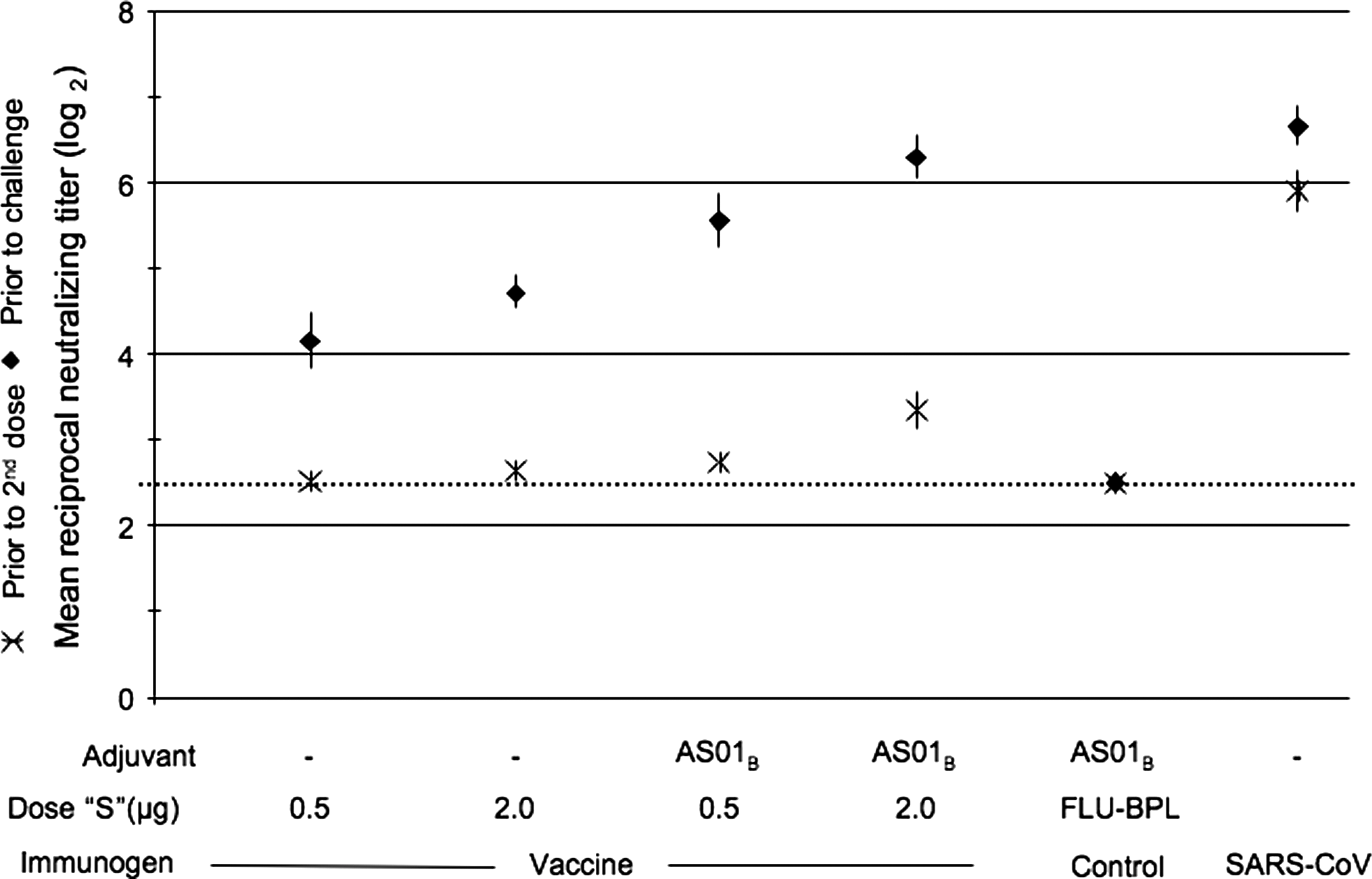
Serum neutralizing antibody responses to WI-SARS vaccine in golden Syrian hamsters. Vaccine dose and adjuvant are indicated along the x-axis; the dose listed is the amount of SARS-CoV S-protein (“S”) administered to each hamster. Control groups include hamsters immunized with a Vero cell-grown BPL-inactivated influenza virus vaccine (Flu-BPL), and primary infection with SARS-CoV. Neutralizing antibody titers are expressed as the reciprocal mean titer (log2) for groups of 3 hamsters. Data points show mean titers achieved following one and two doses of vaccine as measured prior to the administration of dose 2 and prior to challenge, respectively. Error bars indicate standard error of the mean.
At a lower dose of WI-SARS (0.5 μg S-protein without adjuvant), 6 of 14 hamsters seroconverted at 3 wk post-immunization, while in the presence of AS01B adjuvant, 11 out of 14 hamsters had detectable SARS-CoV-specific serum neutralizing antibody responses. With a fourfold higher vaccine dose (WI-SARS 2.0 μg S-protein) without adjuvant, 9 of 14 hamsters seroconverted following a single dose of vaccine. In the presence of adjuvant AS01B, all 14 hamsters seroconverted after a single dose. The variability in antibody responses among age- and gender-matched hamsters likely reflects the fact that they are outbred (15).
Following a second immunization, all hamsters seroconverted to SARS-CoV, except two animals that had received WI-SARS 0.5 μg S-protein in the absence of adjuvant. Although the inclusion of the AS01B enhanced antibody levels, the antibody levels in the hamsters that received two doses of vaccine still did not reach those seen in mice following 2 doses of vaccine.
The sera collected at 3 wk and 16 wk following the second dose of vaccine (prior to early and late challenge) were tested together. A decline in titer was noted in all vaccinated groups at 16 wk post-immunization, but titers were stable in the control group of hamsters that were infected with SARS-CoV (Fig. 6). In the absence of adjuvant, a dose-dependent response was seen with WI-SARS vaccine in hamsters. The antibody titers in the group of hamsters that received a low dose (0.5 μg of S-protein equivalent) of WI-SARS vaccine without adjuvant were just above the limit of detection; mean serum neutralizing antibody titers were higher in hamsters that received 2 μg of S-protein than in hamsters that received 0.5 μg of S-protein.
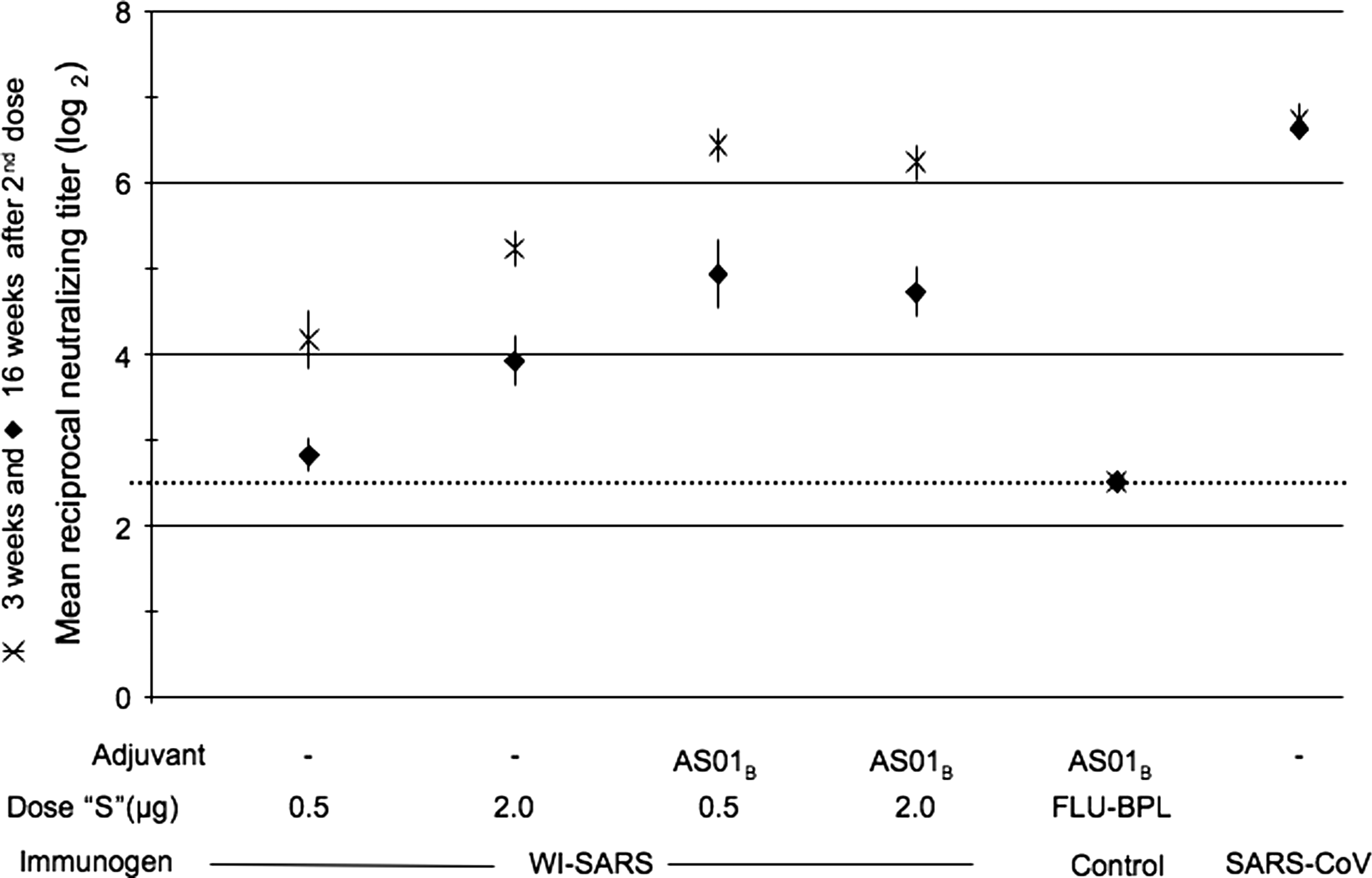
Neutralizing antibody responses to WI-SARS vaccine in golden Syrian hamsters in sera collected 3 and 16 wk following two doses of vaccine. Vaccine dose and adjuvant are indicated along the x-axis; the dose listed is the amount of SARS-CoV S-protein (“S”) administered to each hamster. Control groups included hamsters immunized with a Vero cell-grown BPL-inactivated influenza virus vaccine (Flu-BPL), and primary infection with SARS-CoV. Neutralizing antibody titers are expressed as reciprocal mean titer (log2) for groups of 3 hamsters. Data points show mean titers achieved. Sera were tested in the same assay. Error bars indicate standard error of the mean.
Efficacy in hamsters
Two doses of vaccine, with either WI-SARS alone (containing 2 μg equivalent of S-protein) or WI-SARS + AS01B (0.5 or 2 μg of S-protein equivalent) provided complete protection from pulmonary replication of the virus upon challenge 4 wk after vaccination (early challenge; Fig. 7).
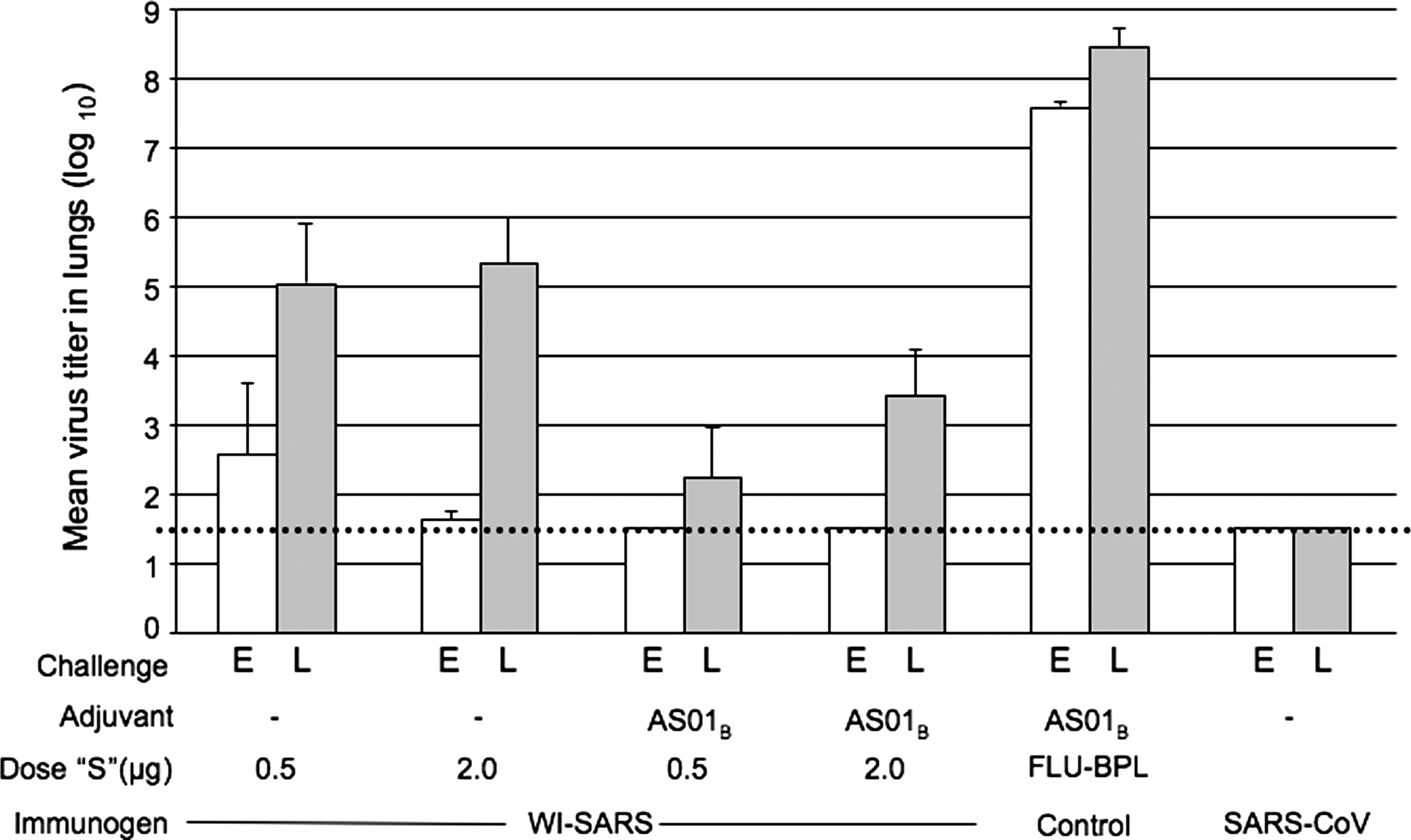
Vaccine efficacy in golden Syrian hamsters at early and late challenge following two doses of WI-SARS vaccine. Vaccine dose and adjuvant are indicated along the x-axis; the dose listed is the amount of SARS-CoV S-protein (“S”) administered to each mouse. Control groups included hamsters immunized with a Vero cell-grown BPL-inactivated influenza virus vaccine (Flu-BPL), and primary infection with SARS-CoV. Shown are mean virus titers in lungs 2 d after challenge with 103 TCID50/mouse SARS-CoV (n = 4 per group). For the schedule of immunization and challenge see Fig. 1B. E indicates early challenge, 4 wk following the second immunization; L indicates late challenge, 18 wk after the second immunization. The dashed line indicates the limit of detection (101.5 TCID50/g). Error bars indicate standard error of the mean.
When hamsters were challenged 18 wk after the second dose of vaccine, none of the vaccinated hamsters were fully protected from replication of the challenge virus. However, the virus titers in animals that received the adjuvanted vaccine were 100- to 1000-fold lower than those seen in animals that received non-adjuvanted vaccine, and the titers in these animals were 1000-fold lower than those seen in the hamsters in the control group that received Flu-BPL + AS01B vaccine. Hamsters that had recovered from SARS-CoV infection were still fully protected from replication of challenge virus 18 wk after infection.
Histological evidence of protective efficacy and absence of histological evidence of complications in hamsters after challenge
Histopathological findings did not vary within groups among tissues collected after early and late challenge; representative photomicrographs from late challenge animals are shown in Fig. 8. In addition to the reduction in viral titers detected in the lungs, histopathological findings in the lungs of immunized hamsters indicate that WI-SARS vaccine with or without adjuvant confers protection from homologous SARS-CoV challenge. The three Flu-BPL-vaccinated control animals lacked detectable levels of SARS-CoV neutralizing antibodies, as did one hamster immunized with WI-SARS 0.5 μg S-protein in the absence of adjuvant. These animals had severe lung lesions, including perivascular lymphocytic cuffs, bronchiolar lesions, vasculitis with subendothelial infiltrates, and interstitial alveolar lesions including adenomatous lesions (peribronchiolar alveolar epithelial proliferative lesions; Fig. 8A and B). Hamsters that had detectable levels of serum neutralizing antibodies to SARS-CoV at the time of challenge were protected from severe lung lesions (Fig. 8C–F). Hamsters that were vaccinated with S-protein alone or with adjuvant had few pathologic lesions (Fig. 8C and D), and hamsters that were previously infected with SARS-CoV had only mild focal lung lesions of alveolar bronchiolization (Fig. 8E), without viral antigen on IHC (Fig. 8F). Histopathology in the liver was limited to occasional small foci of lymphoid inflammation, which was found in all groups (n = 3 per group), except in the previously SARS-CoV-infected and recovered group. No apparent exacerbation of disease was observed in any group, nor was a direct correlation between SARS-CoV neutralizing antibodies and the presence of liver foci apparent (data not shown).
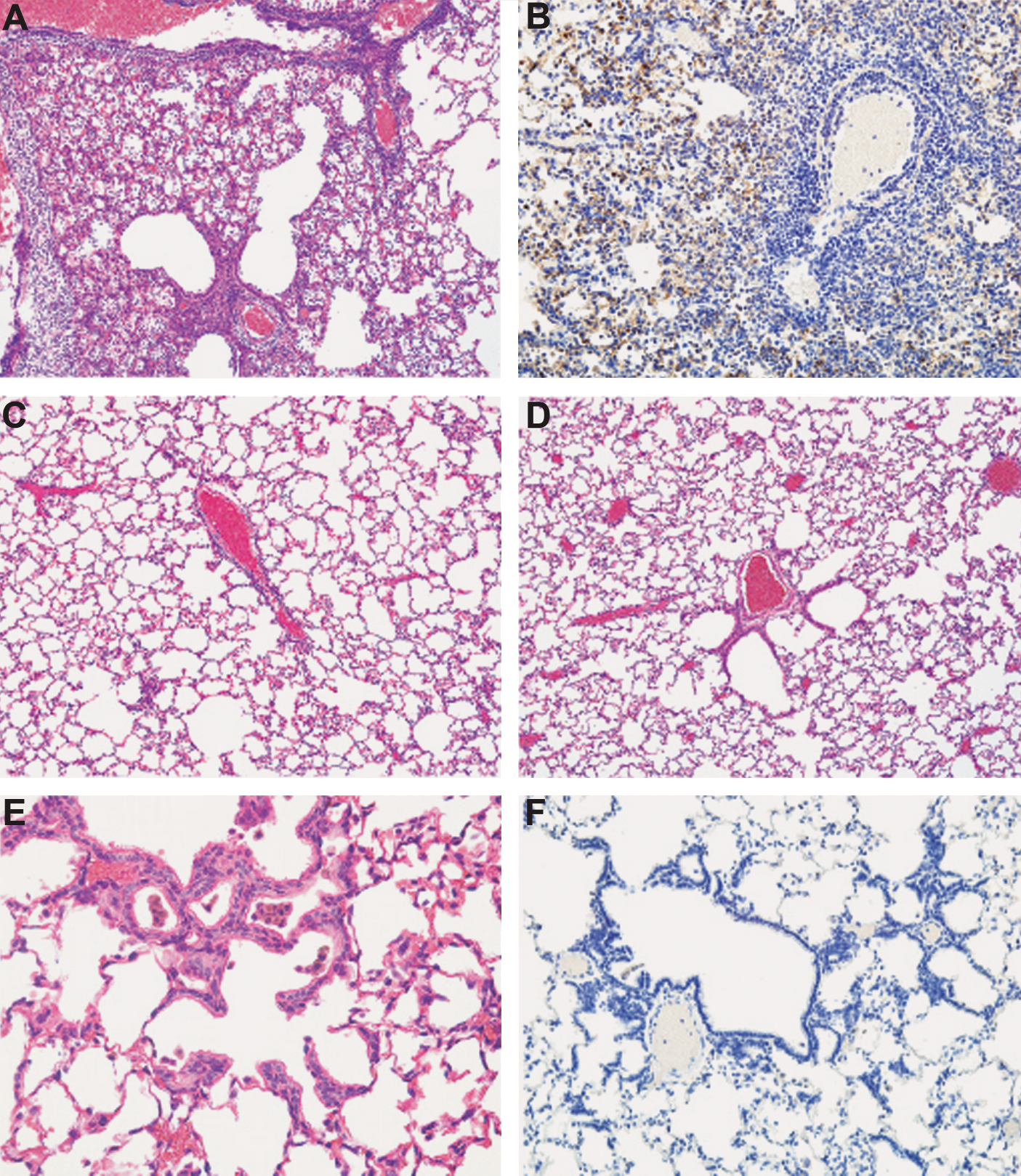
Histopathological findings in the lungs of hamsters sacrificed on day 5 following late challenge. (
Discussion
Vaccines based on whole inactivated viruses have some advantages over other vaccine approaches. In a comparison of vaccines against influenza A virus in two different mouse strains, a whole inactivated virus vaccine produced higher neutralizing and hemagglutination inhibition antibody titers than either virosomal or subunit vaccines (22). The presentation and stability of the antigenic targets for viral neutralization in a whole inactivated vaccine mimic those of the wild-type virus better than subunit or virosomal vaccines, and this may account for the greater immunogenicity of whole-virion vaccines. Furthermore, whole inactivated vaccines can present complex antigenic targets for neutralization that subunit vaccines are not able to reproduce, such as conformational interactions between VP1 and VP3 of hepatitis A virus (23). Although antibodies raised to SARS-CoV S-protein are sufficient for virus neutralization, whole inactivated vaccines present the S-proteins in trimeric conformation, mimicking the wild-type virus. Additionally, whole inactivated vaccines are also more cost effective and more easily generated than recombinant vectored vaccines.
The inactivated vaccine used in this study was prepared by propagation of the SARS-CoV Urbani strain in Vero cells and subsequent inactivation with BPL, using methods that can be easily scaled up. BPL was selected as the inactivating agent because it can break the structure of nucleic acids through reaction with the purine base, and has been widely used for the inactivation of DNA and RNA viruses. See et al. (13) found that a BPL-inactivated SARS-CoV vaccine adjuvanted with alum generated higher titers of neutralizing antibodies and was more protective in mice than a recombinant adenovirus vaccine expressing S and N proteins. Multiple virus inactivation approaches, such as UV irradiation, formalin, or BPL treatment, have been used to prepare whole virus-inactivated vaccines (6). Interestingly, SARS-CoV virions that were inactivated by UV irradiation and additional formalin treatment skewed the cellular immune response toward the Th-2 type, possibly because carbonyl groups were generated by formalin, as was recently described by Moghaddam et al. (24) and Tsunetsugu-Yokota et al. (25). Vero cells are a certified cell line currently used for the production of registered poliovirus and rotavirus vaccines. High infectious titers and virion yields were obtained during preparation of the SARS-CoV vaccine. These features present an attractive basis for further industrial development.
Several studies have demonstrated the feasibility of different vaccine approaches against SARS (6,11). The results of many of these studies indicate that neutralizing antibody responses to the S-protein are an important correlate of immunity against SARS in preclinical models. The magnitude of the antibody response can be significantly increased with adjuvants (12). Recent studies employing a new generation of adjuvants developed by GlaxoSmithKline Biologicals have clearly demonstrated the superiority of these Adjuvant Systems over alum or non-adjuvanted vaccines (e.g., HPV 16/18 L1 VLP adjuvanted with AS04) (26). Additionally, recombinant circumsporozoite protein-based vaccine RTS,S adjuvanted with AS01B (27), and seasonal and pandemic split influenza vaccine adjuvanted with AS03 (28), were more robust in terms of dose-sparing, breadth of cross-protection, and the induction of long-term immunity. The present work further establishes the benefit of using the new generation of proprietary GlaxoSmithKline Biologicals Adjuvant Systems for improved efficacy against SARS CoV.
The Adjuvant System AS01B contains two potent immunostimulants: MPL (3-O-desacyl-4-monophosphoryl lipid A), and QS21 (highly-purified saponin extracted from the bark of a South American tree, Quillaja saponaria Molina), and has been extensively evaluated in animal models and in clinical trials (29 –31). Activation of toll-like receptor 4 by MPL acts not only on innate immune responses, but also on the induction of both humoral and cellular immune responses. In addition, QS21 has also been shown to improve humoral and cell-mediated immunity in preclinical and clinical trials (32). Notably, the animals vaccinated with vaccine including the AS01B adjuvant did not develop measurable CD8 T-cell responses in PBMCs. Nevertheless, in a previous experiment CD8 T-cell responses were detected in the spleens of animals immunized with this adjuvanted vaccine. This response could play a role in protection against SARS CoV. Although AS03A can be considered to be a potent adjuvant for the antibody response, this oil-in-water–based adjuvant has been shown to induce local inflammation and attract immunocompetent cells to the injection site (28). Furthermore, the plant-derived liposoluble vitamin E (DL-α-tocopherol) included in this Adjuvant System increases antibody titers by enhancing production of cytokines, leading to greater proliferation of T cells through reduced production of prostaglandin PGE2, a T-cell-suppressive factor (33). These characteristics of the AS03A adjuvant could explain the relatively high cellular immune response observed in mice immunized with this adjuvant. In this context, Leroux-Roels et al. (33) found that CD4 T cells (but not CD8 T cells) are important in the control of disease pathogenesis and SARS replication in primary SARS-CoV infection in senescent mice. In this model, depletion of CD4 T cells resulted in an enhanced immune-mediated interstitial pneumonitis, and delayed clearance of SARS-CoV from the lungs, which was associated with reduced neutralizing antibody and cytokine production, and reduced pulmonary recruitment of lymphocytes.
Based on promising data from studies in mice, the WI-SARS vaccine was evaluated in golden Syrian hamsters, a model in which efficacy can be measured by quantitative virology, as well as pulmonary pathology following challenge (15). The BPL-inactivated WI-SARS vaccine was immunogenic and efficacious in hamsters. In general, these observations are consistent with other studies of SARS vaccines, in which vaccines that elicit a robust serum neutralizing antibody response in animal models provide protection from challenge with virus (7,11), though systemic antibodies may not confer sterilizing immunity in all models, and mucosal immunity may enhance the level of protection (8,34). Our study also demonstrated an additional correlate of reduced histological evidence of lung disease in vaccinated hamsters. Furthermore, even at very low neutralizing antibody titers (log2 2.7) there was evidence of protection, with a 100- to 3000-fold reduction in virus titer and reduced histopathology. As recommended by the WHO (11), we also evaluated the efficacy of the WI-SARS vaccine at both early and late time points to assess the duration of protection in hamsters. In our study, vaccine-induced protection lasted 4 months in hamsters, and protection from replication of challenge virus was more robust over time in the presence of AS01B adjuvant than in its absence (two-way ANOVA p ≤ 0.05 for comparing the no-adjuvant and AS01B-adjuvant vaccine groups). We examined the lung and liver for evidence of enhanced disease in hamsters following challenge, based on the report of hepatic disease following challenge in ferrets immunized with a modified vaccinia virus expressing the SARS S-protein (35). Even in the absence of detectable serum neutralizing antibodies, enhanced disease was not observed following SARS-CoV challenge.
The data presented in this article reveal that the non-adjuvanted WI-SARS vaccine was immunogenic and provided a high level of protection from early challenge in mice and hamsters. This protection correlated with the presence of high serum neutralizing antibody titers in these animals. However, a decline in antibody titers was observed in hamsters after 16 wk. This decline was less pronounced in animals immunized with the adjuvanted vaccine. Better and more robust protection from late challenge was seen in animals immunized with adjuvanted WI-SARS vaccine than in animals immunized with the non-adjuvanted WI-SARS vaccine. These data were confirmed by the absence of histopathological findings in the lungs of these animals. In addition, these data indicate that the adjuvantation of the WI-SARS vaccine is important for antigen sparing. We have demonstrated the advantages of including an adjuvant to achieve highly protective and sustained immune responses, while maintaining an acceptable level of safety for this vaccine. The use of adjuvants to potentiate the immunogenicity of the WI-SARS preparation did not result in any measurable lung pathology or other safety concerns in vaccinated animals. The decline in the efficacy of the vaccine measured at 18 wk (late challenge), compared to early (4 wk) challenge, coincides with the decline of neutralizing antibody responses seen over time. Interestingly, no decline in protective efficacy upon late challenge was seen in hamsters that were initially infected with live SARS-CoV. Since these animals had similar neutralizing antibody titers at the time of challenge relative to vaccinated animals, it is possible that other components of the adaptive immune response are involved in this sustained and robust protection.
Acknowledgments
We thank Jadon Jackson and the staff of the Comparative Medicine Branch, NIAID, for excellent technical support for animal studies performed at the NIH. This research was supported in part by the Intramural Research Program of NIAID, NIH.
Author Disclosure Statement
Benoît Baras, Geneviève Goossens, Isabelle Knott, and Ventzislav Vassilev are employees of GlaxoSmithKline. No competing financial interests exist for the other authors.