
Abstract
Dendritic cells (DCs) play a pivotal role in the pathogenesis of human immunodeficiency virus-1 (HIV-1). Reduced numbers of blood DCs have been observed in individuals with chronic HIV-1 infection. In the present study, we analyzed the expression levels of monocytes, myeloid dendritic cell (mDC) precursors, mDCs, and plasmacytoid dendritic cells (pDCs), in HIV-1-infected patients in China who were infected via different routes of transmission, including heterosexual and homosexual sexual contact, and blood transmission through importation of blood or blood products, to further elucidate their role in HIV. Compared with HIV-negative individuals (n = 40), relative levels of CD11c+CD14− mDCs, CD11c++CD123low mDCs, and CD11c−CD123+ pDCs in total peripheral blood mononuclear cells (PBMCs) were significantly lower in all HIV patients (n = 93), and in those with blood transmission (n = 26) and heterosexual transmission (n = 43), while relative levels of CD11c+CD14− mDCs were significantly lower in HIV patients infected via homosexual transmission (n = 24). The results of correlation analysis demonstrated a significant negative correlation between CD4+ T-cell counts and the relative levels of CD11c++CD123low mDCs in HIV-I patients infected via blood transmission. There was no significant correlation between CD4+ T-cell counts and the expression level of other DC subpopulations in PBMCs from HIV patients. The results of this study suggest that HIV-1 patients with different routes of transmission exhibit altered expression levels of blood DC subpopulations, which contributes to dysregulated immune responses and pathogenesis of HIV-1.
Introduction
Two subsets of blood DCs have been described, according to their differential expression of CD11c and CD123. The first subset is represented by CD3, CD14, CD20, CD19, and CD56 lineage-negative CD11c+ myeloid DCs, which derive from monocytes and are the main producer of interleukin-12 (IL-12) (8,9), which is a major inducer of Th-1 responses. CD11c+ DCs play an important role in antiviral immune responses by acquiring and processing viral antigens into peptides for major histocompatibility complex presentation to T cells in secondary lymphoid organs (3). The second subset of DCs encompasses lineage-negative CD11c−CD123+ plasmacytoid DCs (pDCs), which are the main producers of type I interferon (IFN-I) in response to viral infections (10,11).
The number and phenotype of DC populations have been shown to be affected during HIV infection. Several studies have shown that during HIV infection, both DC subsets are substantially reduced in the blood (12 –20), skin (21), oral mucosa (22), and lymph nodes (23) of HIV-infected patients. In some of these studies, this decrease correlated with plasma viral load, and was partially reversed following highly active antiretroviral therapy (HAART) (12,15,19,20). It has been hypothesized that HIV-induced maturation of both DC subsets may lead to increased DC migration to lymph nodes, partly explaining their absence from blood as infection progresses (24,25).
Most studies of HIV-1 focus on individuals infected through sexual contact or needle sharing. However, in China one portion of the country's initial HIV-1 infections occurred through contaminated blood and plasma products. It is also possible that there is a difference in DC cell numbers based on whether HIV-1 infection occurs directly or indirectly, entering an organism via mucous membranes or blood. HIV-1-infected individuals were infected mainly through sexual contact (heterosexual contacts and homosexual contacts), and blood transmission (injection of blood or blood products contaminated by HIV-1) in China, but the relative percentages of blood DC subpopulations in peripheral blood mononuclear cells (PBMCs) among those infected by different routes of transmission has to date not been investigated. In this study, we investigated the percentages of monocytes, mDC precursors, mDCs, and pDCs from HIV patients and healthy volunteers, to uncover the role in the pathogenesis of HIV patients infected via different routes of transmission.
Materials and Methods
Study population
Ninety-three HIV-1-infected adults were recruited at the Jiangsu Provincial Center for Disease Prevention and Control. The patients were classified into three subgroups according to the mode of HIV-1 transmission: blood transmission (n = 26), heterosexual transmission (n = 43), and homosexual transmission (n = 24). Descriptive characteristics of the study population are shown in Table 1. The date of infection was estimated on the basis of clinical and laboratory results. None of the study subjects had syphilis or hepatitis B or C virus infection. In addition, 40 uninfected subjects were included as healthy controls. Informed consent was obtained from all subjects, and the research conformed to the ethical guidelines of all of the authors' institutions.
Data are means ± SDs.
HIV, human immunodeficiency virus; M, mean; R, range; SD, standard deviation.
CD4+ T-cell counts of HIV-1-infected individuals
CD4+ T-cell counts were determined as reported elsewhere (26). Briefly, anticoagulated whole blood collected from HIV-1-infected individuals was tested within 6 h for CD4+ T-cell counts. The 50-μL whole blood sample was stained with mixed antibodies (MultiTEST CD3 FITC/CD8 PE/CD45 PerCP/CD4 APC Reagent), and incubated for 15 min in the dark. Then, 450 μL of 1 × lysing solution was added to the solution for 15 min. The labeled cells were analyzed by BD FACSCalibur flow cytometry (BD Biosciences, San Jose, CA), and 105 cells were acquired.
Immunophenotyping analysis of DC cells and monocytes
PBMCs were isolated after whole-blood Ficoll gradient centrifugation. One million PBMCs per sample were used for cell-surface staining. Non-specific sites were blocked by incubation for 10 min on ice with fluorescence-activated cell sorter buffer (1 × PBS, 2% heat-inactivated FBS, and 0.1% sodium azide) containing 12% heat-inactivated FBS and 10 μg of total mouse IgG (Sigma-Aldrich, St. Louis, MO). The cells were labeled with mouse anti-human monoclonal antibodies as previously described: fluorescein isothiocyanate-conjugated anti-CD3, anti-CD19, and anti-CD56 (lineage markers); phycoerythrin (PE)-cyanine (Cy) 5.5-conjugated anti-HLA-DR; allophycocyanin (APC)-conjugated anti-CD11c; and PE-conjugated anti-CD123, anti-CD14, and anti-CD16 (eBioscience, San Diego, CA). Dead cells were identified by trypan blue exclusion. Data acquisition of 100,000 events per sample was performed on a BD FACSCalibur device (BD Bioscience), and the analysis was done using CellQuest™ software (BD Bioscience).
Statistical analyses
Results are expressed as mean ± standard error. The statistical significance of the differences between routes of transmission was assessed using the Mann-Whitney U test. The Wilcoxon signed-rank test was used for pair-wise comparisons of different routes of transmission. Analysis were performed using GraphPad Prism for Windows software (version 5.0; GraphPad Software, San Diego, CA). Spearman's correlation was used to determine significant associations between CD4+ T-cell counts and DC subpopulations. Correlations are reported as the correlation coefficient (r) with associated p value. Analyses were performed using SPSS software (version 17.0; SPSS Inc., Chicago, IL). Statistical significance was set at p < 0.05.
Results
CD4+ T-cell counts
There were no significant differences in the CD4+ T-cell counts among HIV-1 patients of differing routes of transmission (Fig. 1). CD4+ T-cell counts demonstrated a significant negative correlation with the relative level of CD11c++CD123low mDCs in PBMCs from HIV-1 patients infected via blood transmission (r = −0.463, p = 0.017; Fig. 2). However, there were no significant correlations between CD4+ T-cell counts and other DC subpopulations among all patients tested (data not shown).
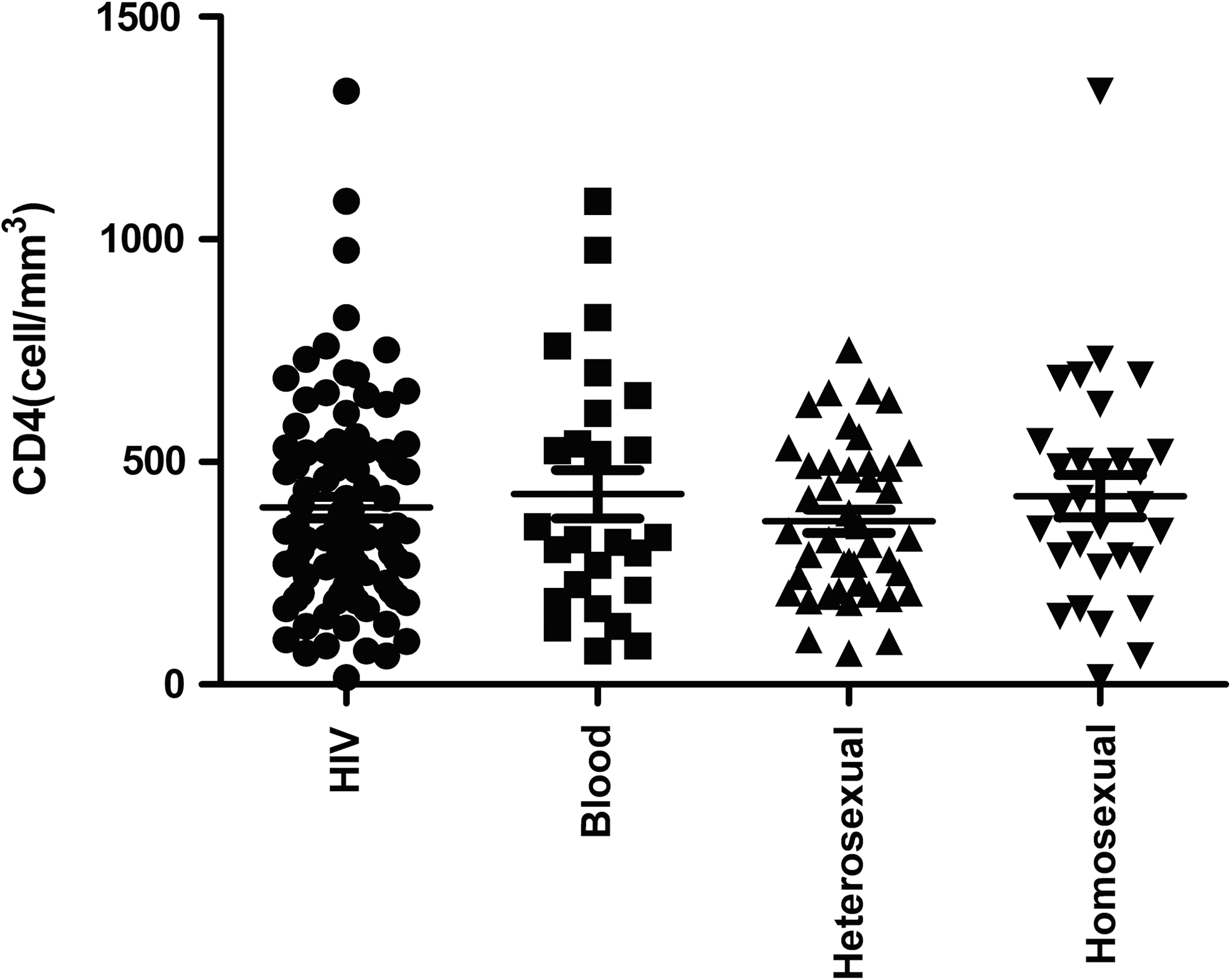
Changes in CD4+ T-cell counts in HIV-1-infected patients with different routes of transmission. Cell populations were evaluated using Wilcoxon's signed-rank and Mann-Whitney U tests for pair-wise comparisons of different routes of transmission, and between study groups, respectively. Statistical significance was set at p < 0.05.

Scatterplot of the correlation between CD4+ T-cell counts and relative percentages of CD11c++CD123low myeloid dendritic cells (mDCs) in peripheral blood mononuclear cells (PBMCs) of blood transmission HIV-infected patients (n = 26).
Relative levels of monocytes in HIV patients infected via different routes of transmission
Compared with HIV-negative individuals, the relative levels of CD11c−CD14+ monocytes in all HIV patients, and those infected via blood transmission, heterosexual transmission, and homosexual transmission, had no significant difference (p = 0.8810, p = 0.8594, p = 0.6834, and p = 0.8292, respectively). Moreover, no significant differences were observed among those infected via different routes of transmission (Fig. 3).

Relative percentages of monocytes. (
Relative levels of mDC precursors in HIV patients infected via different routes of transmission
According to expression levels of the CD11c+CD14+ mDC precursors, no statistically significant differences were observed between HIV-negative individuals and all HIV patients, and those infected via blood transmission, heterosexual transmission, and homosexual transmission (p = 0.3811; p = 0.5681, p = 0.100, and p = 0.5281, respectively). Moreover, no significant differences were observed among those infected via different routes of transmission (Fig. 4).
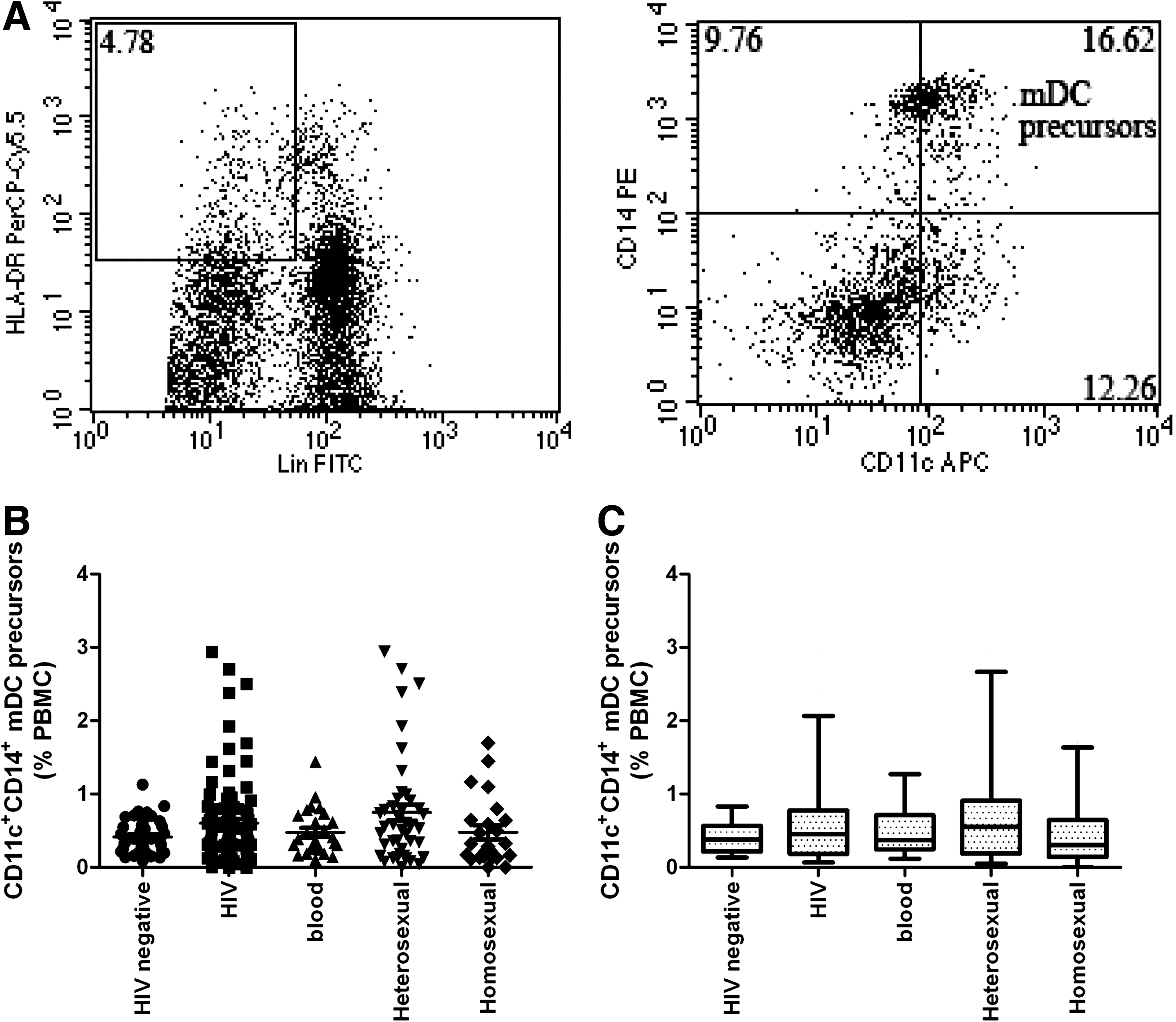
Relative levels of myeloid dendritic cell (mDC) precursor subsets among total peripheral blood mononuclear cells (PBMCs). (
Relative levels of mDCs in HIV patients infected via different routes of transmission
With regard to the variations in the relative frequencies of CD11c+ mDC populations, we identified two different phenotypes: CD11c+CD14− and CD11c++CD123low (Fig. 5A and 5B). We found that the percentage of the CD11c+CD14− subset was significantly lower in HIV-negative individuals than in all HIV patients, and those infected via blood transmission, heterosexual transmission, and homosexual transmission (p < 0.0001, p < 0.05, p < 0.01, and p < 0.0001, respectively). Moreover, significant differences were detected between those infected via homosexual transmission and blood transmission and those infected via heterosexual transmission (all p < 0.01). However, no statistically significant difference was observed between those infected via blood transmission and those infected via heterosexual transmission (p = 0.7522; Fig. 5C and D).
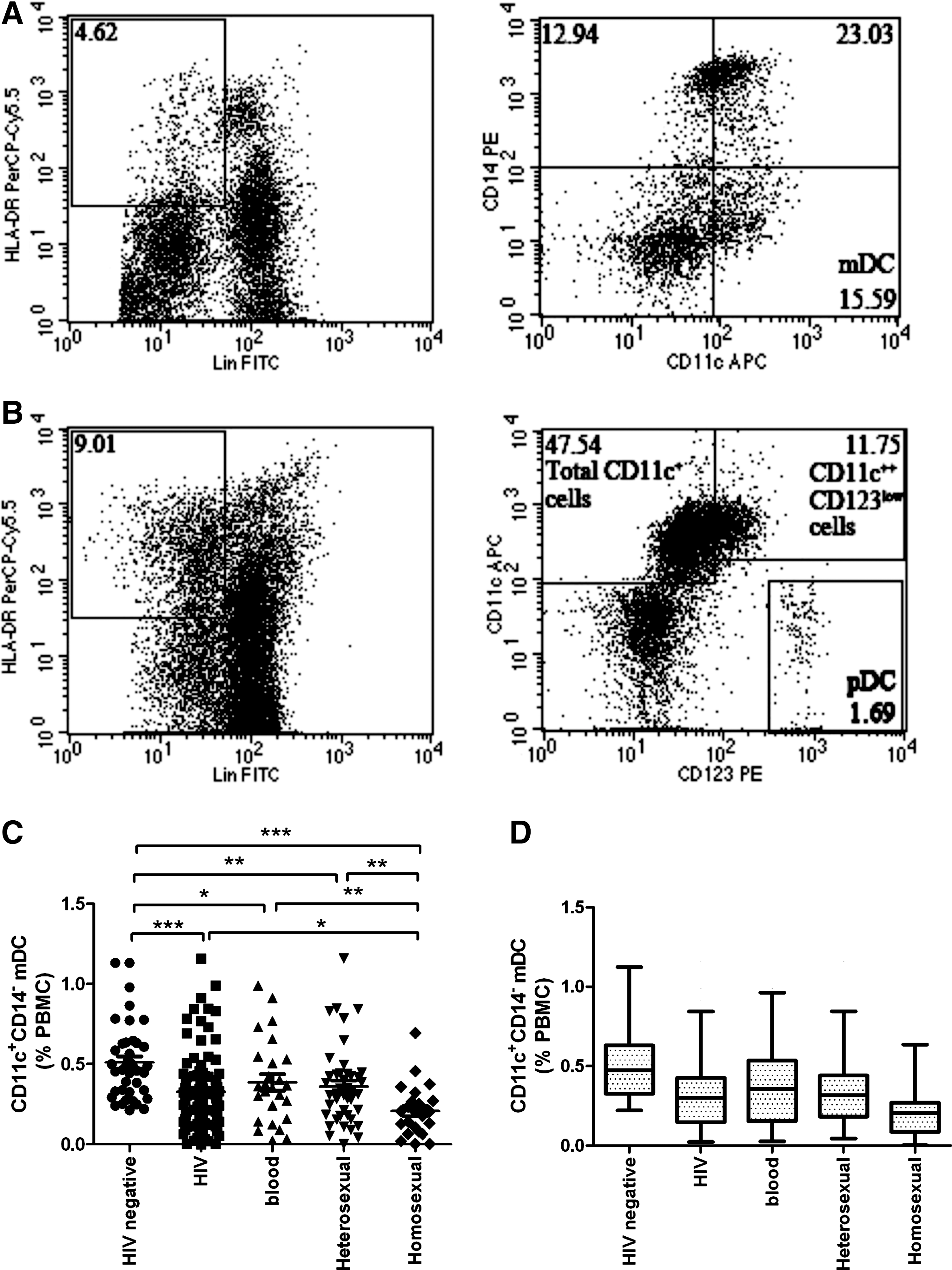
Relative percentages of myeloid dendritic cell (mDC) and plasmacytoid dendritic cell (pDC) subpopulations among total peripheral blood mononuclear cells (PBMCs). (
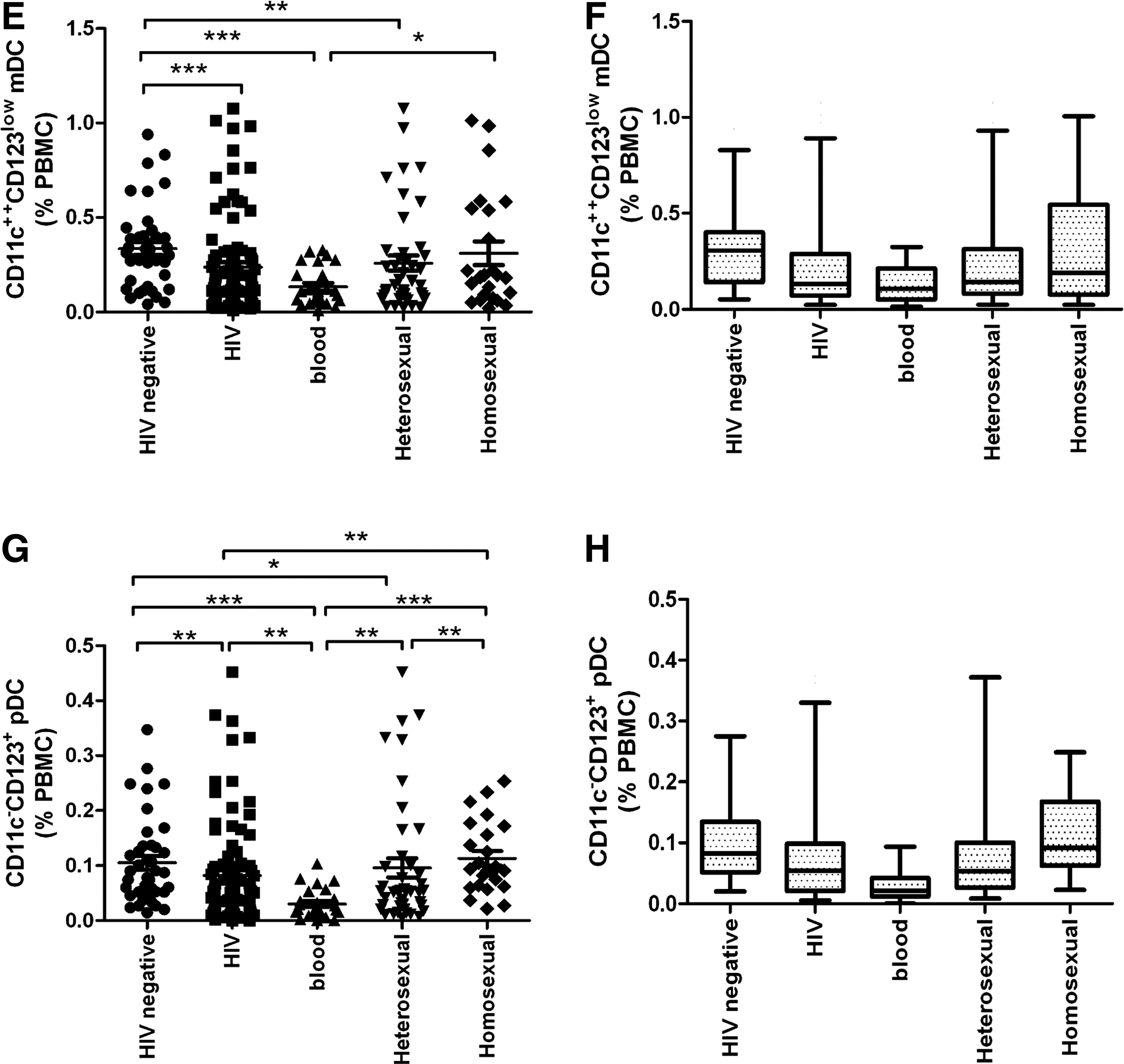
Compared with HIV-negative individuals, all HIV patients, and those infected via blood transmission and heterosexual transmission had significantly lower CD11c++CD123low mDC levels (p < 0.001, p < 0.0001, and p < 0.01, respectively). However, no significant difference was found between HIV-negative individuals and patients infected via homosexual transmission (p = 0.1948). Moreover, a significant difference was detected between patients infected via blood transmission and those infected via homosexual transmission (p < 0.05). However, the relative levels of CD11c++CD123low mDCs in patients infected via blood transmission and those infected via homosexual transmission showed no significant difference compared to those infected via heterosexual transmission (p = 0.0590 and p = 0.6519, respectively; Fig. 5E and F).
Relative levels of pDCs in HIV patients infected via different routes of transmission
Blood pDCs are defined as Lin−HLA-DR+CD11c−CD123+ (Fig. 5B). In our study, the relative levels of pDCs in all HIV patients, and those infected via blood transmission and heterosexual transmission were significantly lower than in HIV-negative individuals (p < 0.01, p < 0.0001, and p < 0.01, respectively), but were not significantly different than those infected via homosexual transmission (p = 0.3315). Patients infected via blood transmission were significantly different than those infected via heterosexual transmission and homosexual transmission (p < 0.01 and p < 0.0001, respectively). Moreover, a significant difference was observed between patients infected via heterosexual versus homosexual transmission (p < 0.01; Fig. 5G and H).
Discussion
Here, for the first time, patients with different routes of transmission were studied for more than 1 y after HIV-1 infection with regard to the percentage of blood DC subpopulations in PBMCs. Also for the first time, differences in the relative levels of DC subpopulations in those infected via different routes of transmission were examined, and in this study we demonstrated a negative correlation between CD4+ T-cell counts and CD11c++CD123low mDC levels in patients infected via blood transmission.
The ex vivo flow cytometry analysis of blood mDCs revealed two different phenotypes of mDCs, namely CD11c+CD14− and CD11c++CD123low. They were present in both HIV-infected and HIV-negative individuals, and may reflect different subpopulations and/or stages of differentiation. mDCs enhance HIV-1 infection via capture of the virus and subsequent transmission to T cells. In this article, we found that the percentage of the CD11c+CD14− subset was lower in HIV-infected individuals than in HIV-negative individuals. In addition to those infected via homosexual transmission, the other HIV-infected groups had lower CD11c++CD123low mDC levels than HIV-negative individuals. Moreover, significant differences were detected among those infected via different routes of transmission, suggesting that the phenotypic disturbances observed in HIV-1 blood mDCs in those with different routes of transmission may have resulted in altered distributions of mDCs in peripheral tissues, contributing to dysregulated immune responses and the acquired immune deficiency syndrome (AIDS).
The numbers of mDCs were significantly reduced among all HIV-1-infected patients (12,13,15,18,20,24,27). Fontaine et al. found that the relative levels of blood mDC and pDC populations decreased significantly in those with untreated primary HIV infection who had differing rates of disease progression, and that in individuals with acute infection the levels remained low during the first 2 y of infection in rapid progressors who responded to ART (28). Our results are consistent with previous findings (28). It has been proposed that DC loss is due to increased recruitment to lymph nodes, although this has not been directly tested. Similarly to HIV-infected humans, we found that mDCs were lost from the blood of SIV-infected rhesus macaques with AIDS (29). Although the mechanisms used by HIV to affect DC populations are not fully understood, the cause of blood DC reductions may be either central or peripheral. In the periphery, DCs may have homed to lymphoid organs and may no longer be present in the blood. An elevated frequency of CD1a+ DCs was found in the tonsils of HIV-positive patients, and these cells may have originated in mucosal Langerhans cells or in blood DC precursors (30). DCs may be destroyed in the periphery by the virus itself, by CD8+ cytotoxic T lymphocytes, by a lack of survival factors, or by other mechanisms. Conversely, the causes of DC reductions may be central, due to a decreased production of DCs by bone marrow CD34+ cells or blood monocyte precursors. On the other hand, the low levels of mDCs in circulating blood suggest that they are actively recruited to lymphoid organs in response to the ongoing attempt of the immune system to fight disease progression. This has been shown in the HIV-transgenic mouse model, in which DCs accumulate in the lymph node subcapsular sinus and spleen (31).
Blood pDCs play an important role in the innate immune responses to different types of viruses, including HIV-1. pDCs inhibit HIV-1 replication in T cells via the secretion of IFN-α (32). During chronic HIV infection, a defective type I IFN in vitro response of DCs to viral stimuli has been demonstrated (33). The relative levels of CD11c−CD123+ pDCs in all HIV patients, and in those infected via blood transmission and heterosexual transmission were lower than in HIV-negative individuals; however, no significant difference was observed with those infected via homosexual transmission. Our results are consistent with prior findings that circulating pDC numbers are reduced in chronic as well as primary HIV infection (34). The relative decrease in blood pDC levels maybe due to cell migration into targeted areas, because HIV-1 has been shown to induce expression of the migration marker CCR7 (35). Blood pDC and IFN levels are decreased during the advanced stages of HIV infection, but can be restored by ART (13). Our results show that pDC levels can be restored during ART in patients infected via homosexual transmission. However, in other types of HIV-infected patients, levels remain low, even after ART is initiated. We suggest that the different pDC subsets seen with different routes of transmission may cause altered phenotypes and modified migratory capacity, suggesting a role in the pathogenesis of AIDS. Blood pDC levels have been shown to be correlated positively with CD4+ T-cell counts, and inversely with viral loads (14,27). In the present study, no significant correlation was seen between CD4+ T-cell counts and pDC levels. This discordant finding may be due to the use of differing study populations.
The CD11c+CD14+ mDC precursor in human peripheral blood has the ability to migrate to sites of inflammation and to differentiate into mature mDCs (36,37). In contrast to mDCs, no significant differences were observed in blood mDC precursors in HIV patients infected via different routes of transmission versus HIV-negative individuals. Our results are consistent with prior findings that the levels of blood mDC precursors are decreased during the acute phase of infection, and that they increase above normal levels after the initiation of ART and during chronic infection (28).
Conclusion
In the present study, we provide the first evidence of altered expression levels of blood DC subpopulations in HIV-1 patients from Jiangsu Province, China, who were infected via different routes of transmission. In light of the present results, the changes in DC subpopulations may be partly due to differing routes of infection, such as via sexual contact or via blood transmission. These changes may play a role in limiting the spread of the virus, and thus allow some HIV-1 patients to slow the progression of the disease. We need to further explore the clinical significance of these differing expression levels of blood DC subsets in HIV-1 patients infected via different routes of transmission.
Acknowledgments
This study was supported by the Natural Science Fund of Jiangsu Province (BK2009435), and the Ministry of Science and Technology (MOST) 973 Project (grant 2006CB504203). We wish to thank the HIV-1 patients for their special contribution to this study.
Author Disclosure Statement
No competing financial interests exist.