
Abstract
The pathogenic mechanism of dengue virus infection is related to the host responses within target cells, and therefore we assessed intracellular changes in stress proteins following dengue virus infection. This study provides evidence that Hsp70 helps in viral multiplication by suppressing the type 1 interferon response. Dengue virus infection in human monocytic THP-1 cells led to overexpression of Hsp70, which also acts as a chaperone. The functional role of Hsp70 in dengue virus multiplication was identified by downregulating the Hsp70 gene with its specific siRNA duplexes, which led to a decrease in viral RNA copy numbers in cellular supernatants and intracellular viral load. It also resulted in an increased IFN-α level, which mediates its antiviral effect through double-stranded RNA-induced protein kinase-PKR. Collectively these results suggest that an increased level of Hsp70 expression in dengue-virus-infected THP-1 cells assists in viral replication by escaping the antiviral effect of type 1 interferon.
Introduction
The mechanism involved in the pathogenesis of dengue infection, especially that of DHF/DSS, remains unresolved. The pathogenic effects of infection remain poorly understood. Immunopathological examination suggests the involvement of different tissues during dengue infection, as viral antigens are expressed in liver, lymph node, spleen, and bone marrow (8,9). Blood monocytes and macrophages are the major cell types reported to display dengue antigens in autopsy samples (10,11). Monocytes and macrophages constitute an extremely heterogeneous population of cells, most probably as a result of differentiation during the 36–104 h that they spend in the circulation (12). From the results of previous studies it appears that dengue virus enters target cells after the envelope protein E attaches to an uncharacterized receptor that may display highly-sulfated glycosaminoglycans. Binding moieties on the cell surface membrane may vary between cell types and species origin, as well as for the different serotypes (1 –4). Tsai et al. (13) reported an interaction between dengue virus and human toll-like receptors (TLRs) in vitro. The expression of TLR3 can mediate strong interferon (IFN)-α/β release and inhibit viral replication significantly. Moreover, dengue is strictly a human pathogen, and no appropriate animal model is available, and thus it is important to understand the interaction of dengue virus with its principal target cells, and to ascertain if there is any difference between differentiated and undifferentiated monocytes/macrophages in response to infection, and their potential impact on immunity and pathogenesis.
Dengue virus utilizes host cell proteins for its replication inside the cells, including heat-shock proteins (Hsps), a group of evolutionary-conserved chaperone proteins (14). Hsps have been divided into different families based on their approximate molecular weights. The five main groups are Hsp100, Hsp90, Hsp70, Hsp60, and the small Hsps. The Hsp70 family represents the most highly conserved protein, with functional counterparts in the most primitive form, like bacteria, as well as in more sophisticated forms of higher organisms (14). Hsp70 functions as an ATP-dependent molecular chaperone that assists in the folding of newly synthesized polypeptides, the assembly of multiprotein complexes, the transport of proteins across cellular membranes, and targeting of proteins for lysosomal degradation (15). Hsp70 is expressed only at low or undetectable levels in most unstressed normal cells, but its expression is rapidly induced by a variety of stressors (e.g., viral infection), and therefore it is often called the major stress-inducible Hsp70. Stress-induced accumulation of Hsp70 accelerates cellular recovery by enhancing the ability of stressed cells to cope with increased concentrations of unfolded/denatured proteins (16). It has been reported that Hsp70 and Hsp90 participate in dengue virus entry as a receptor complex, and also in virus multiplication in human monocytes/macrophages (17,18). Our recent study (19) has shown that Hsp60 is overexpressed in promonocytic cell line U937, which suggests the expression of different cell-stage-specific stress proteins. In this study we investigated the characteristics of Hsp70 upregulation in dengue-virus-infected cells, and the potential role that Hsp70 may play in a dengue virus infection model. The results show for the first time that Hsp70 is upregulated in dengue-virus-infected THP-1 cells, coincident with high levels of intracellular viral load. Knockdown of Hsp70 resulted in a dramatic reduction in intracellular dengue virus load and an increased IFN-α level. This suggests the involvement of Hsp70 in the immunopathogenesis of dengue virus infection.
Materials and Methods
Cell line
The monocytic THP-1, BHK-21, and C6/36 were obtained from the National Centre for Cell Science, Pune, India. THP-1 was maintained in RPMI-1640 (Sigma-Aldrich, St. Louis, MO), supplemented with 10% fetal bovine serum (FBS; Sigma-Aldrich), and BHK and C6/36 were propagated in MEM (Sigma-Aldrich), supplemented with 10% FBS at 37°C and 28°C respectively in a 5% CO2 atmosphere.
Virus
Dengue-2 virus New Guinea C strain obtained from the International Centre for Genetic Engineering and Biotechnology, Delhi, India, was expanded in a C6/36 insect cell line. Briefly, the tissue culture supernatant (after centrifugation) obtained from dengue-2-virus-infected C6/36 cells was directly used as the source of virus.
Infection of cells
THP-1 cells were infected with the dengue virus at a multiplicity of infection (MOI) of 2. The virus inoculum was incubated (2 × 105 cells/well) in serum-free medium at 37°C for 1 h to permit viral adsorption. The culture plates were gently agitated for optimal virus-cell contact. Thereafter, the unadsorbed virus was removed by washing the cells three times with plain medium. The dengue-virus-infected and non-infected cells were replenished with fresh complete medium and incubated for another 3 d. At the end of the incubation, cells were harvested and the cell-free supernatant was stored in aliquots at −70°C until assayed for infectious-virus production and other parameters.
Cytopathic effect
The cytopathic effect in infected THP-1 cells was studied after infection with dengue virus for 3 d. The cells were washed with phosphate-buffered saline (PBS), fixed with paraformaldehyde on a glass slide, and hydrated by passing through decreasing concentrations of graded alcohol solutions. The slides were stained with hematoxylin for 5–10 min, quickly rinsed in water, and then dipped into 1% HCl (in 70% alcohol), and finally counterstained with 1% eosin for 15 sec to 2 min, followed by dehydration by passing through graded alcohol solutions. Destaining was performed using xylol, and the slides were mounted with slide mounting medium. The cytopathic effect was observed under 100 × magnification.
Plaque assay
BHK-21 cells were seeded in 6-well plates at a density of 1 × 105 cells per well, in 3 mL of MEM supplemented with 10% FBS at 37°C for 24 h. Culture supernatant from dengue-virus-infected cells was serially diluted, and 1 mL of dengue virus sample was added to each well in triplicate. After incubation at 37°C for 2 h, medium containing MEM and 1% agarose (Sigma-Aldrich) with 2% FBS was layered in each well, and the plates were incubated at 37°C for 7 d. Plaques were counted after fixation with 3.7% formaldehyde and removal of the agarose plug, followed by staining with 1% crystal violet solution in 20% methanol. Virus concentrations were determined as plaque-forming units (pfu) per milliliter, and used to calculate the MOI.
Immunoblotting
Dengue-virus-infected and non-infected THP-1 cells were sonicated in the presence of protease inhibitors (1 mM PMSF and 2 mM EDTA; Sigma-Aldrich). Total protein was estimated by the method of Lowry's group (20). Equal amounts of protein (50 μg) prepared in sample buffer were loaded into separate wells of 10% SDS-gel and subjected to electrophoresis, essentially following the method described by Laemmli (21). When the separation of protein was complete, the protein was transferred onto nitrocellulose membranes (Millipore, Billerica, MA), using transfer buffer (48 mM Tris, 39 mM glycine, and 20% v/v methanol), in a semi-dry blotting apparatus (Bio-Rad Laboratories, Inc., Hercules, CA), following the method described by Towbin and associates (22). The nitrocellulose membrane was blocked with blocking buffer (3% powdered skimmed milk and 0.1% Tween-20 in PBS [PBST20, pH 7.4]) for 1 h at room temperature, and then washed three times for 10 min each with PBST20 at room temperature. The washed membranes were further incubated for 1 h with monoclonal antibody (MAb) against dengue virus E protein (Biotrend Chemicals, Destin, FL) at 1:100 dilution, and MAb against Hsp27, Hsp60, Hsp70, and Hsp90 (Santa Cruz Biotechnology, Santa Cruz, CA), at 1: 1000 dilution each, and protein kinase-PKR (Santa Cruz Biotechnology) at 1:100 dilution. Incubation was followed by washing with PBST20; the membranes were again incubated for 1 h with peroxide-conjugated rat anti-mouse IgG (Neomarker, Thermo Fisher Scientific, Fremont, CA) at 1:10,000 dilution. After washing three times with PBST20, the membranes were processed for color development using diaminobenzidine (DAB; Sigma-Aldrich) and H2O2 as an oxidizing agent in PBS. The cells were also exposed to 42°C for expression of Hsp70 as a positive control. Whole-cell lysates were used for immunoblot analysis.
Flow cytometry
Infected THP-1 cells were harvested and washed twice with 0.01 M PBS. The cells were permeabilized with 70% ethanol for 30 min in FACS tubes (BD Biosciences, San Jose, CA). The cells were washed three times and incubated with anti-dengue virus MAb (1:100 dilution as the final concentration) for 60 min. The cells were washed three times and incubated for another 60 min with FITC-labeled anti-mouse IgG secondary antibody. After washing, the cells were suspended in 0.5 mL of PBS, and 10,000 cells were acquired with a flow cytometer (BD FACScalibur), using CellQuest software (19).
Transfections with siRNA
Infected and non-infected THP-1 cells (1 × 106 for flow cytometry, and 5 × 106 for immunoblot analysis) were seeded onto 6-well plates. The cells were transfected with commercially available Hsp70-specific siRNA, using Hiperfectamine in a 1:6 ratio (1 μg siRNA to 6 μL/HiPerFect; Qiagen, Hilden, Germany). The cells were allowed to incubate for 8 h in an incubator with 5% CO2. The cells were washed three times with plain medium and infected with the dengue virus (MOI 2) for 1 h with constant agitation. The cells were washed again three times and further cultured for 72 h in RPMI medium supplemented with 2% FBS in an incubator with 5% CO2.
Real-time PCR
Viral RNA copy number was determined using reverse transcriptase real-time PCR analysis. The cell supernatant was harvested and viral RNA was isolated using a viral RNA isolation kit (Qiagen). This viral RNA stock was used for setting up the real-time PCR reactions using dengue-virus-specific primers and TaqMan probes (Genome Diagnostics, New Delhi, India) using a thermal cycler with florescence measurement capability.
Elisa
Solid phase enzyme-linked immunosorbent assay was employed using the multiple antibody sandwich principle (19). ELISA for human IFN-α was carried out using a commercial kit (Antigenix America, Huntington Station, NY). Anti-IFN-α antibody-coated plates were incubated for 2 h at 37°C. After adding the standards and the samples, the plates were washed three times with wash buffer, then incubated with anti-IFN-α biotinylated rabbit polyclonal antibody per the manufacturer's instructions, and washed three times using wash buffer. Streptavidin-conjugated horseradish peroxidase was added to each well, followed by peroxide/tetramethyl benzidine substrate solution. Absorbance was measured on an ELISA reader at 450 nm. A standard curve was obtained by plotting a curve between known concentrations of IFN standards (x axis) and absorbance (y axis). The concentrations of the cytokine in experimental samples were determined from the standard curve drawn using SPSS 15.0 software (SPSS, Inc., Chicago, IL).
Statistical analysis
The results are expressed as mean ± SEM and all the statistical comparisons were carried out using Student's t-test for repeated measurements when applicable. The significance level was set at p < 0.05. Data shown are a result of three independent experiments.
Results
Confirmation of infection
Cytopathic effect (CPE)
The cells were infected with virus at 70% confluency for 3 d. Infected THP-1 cells on staining with hematoxylin and eosin showed a significant difference from non-infected cells (Fig. 1). Infected cells were larger in size with oval or round morphology. Crenation of cells was observed with clear features of cellular damage at a dose of MOI 2; however, no such morphological changes were observed in non-infected cells.
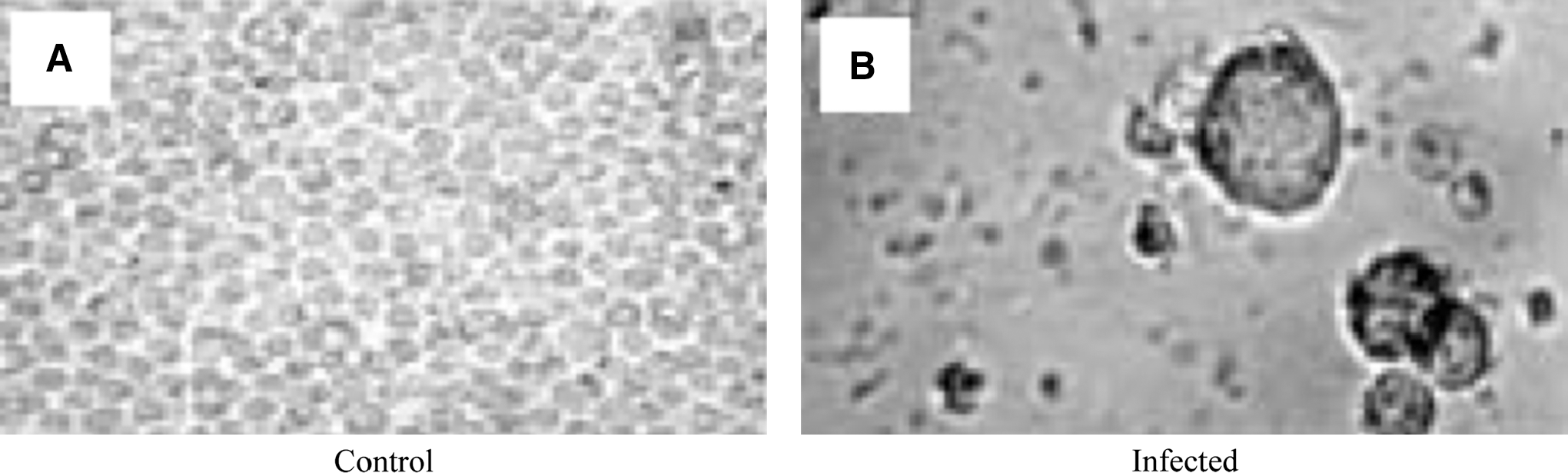
Infection confirmation by cytopathic effect. THP-1 cells were infected with dengue virus, and the cells were stained with hematoxylin and eosin.
Plaque assay
THP-1 cells were infected with dengue virus, and after 3 d of full-blown infection, the supernatant was collected and added to 70% confluent BHK-21 cells for 7 d, resulting in the formation of plaques (Fig. 2). The plaques were counted as plaque-forming units per milliliter (data not shown), confirming viral replication in dengue-virus-infected THP-1 cells.
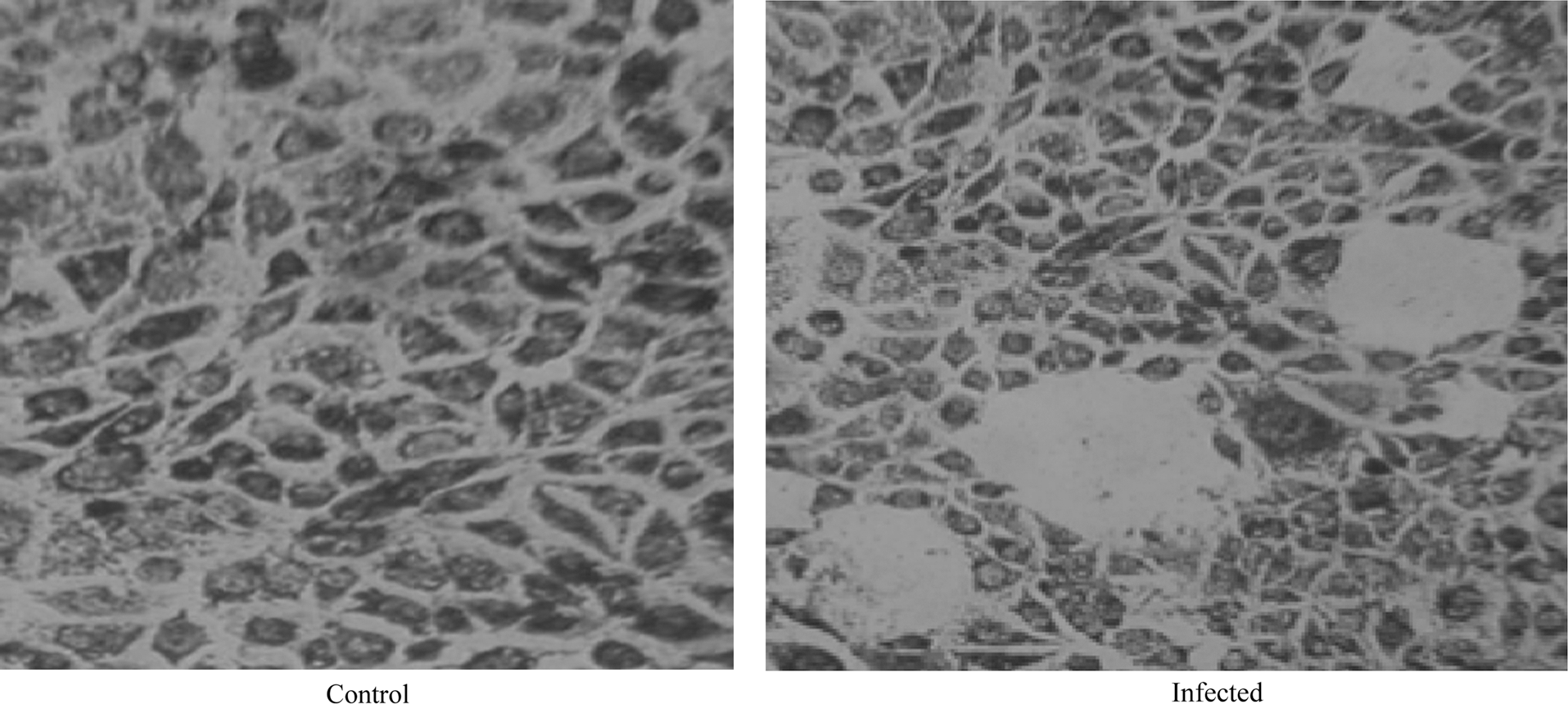
Plaque formation in BHK-21 cells. Shown are control (non-infected) BHK-21 cells (left panel), and infected BHK-21 cells (right panel), inoculated with culture supernatant of dengue-infected THP-1 cells.
Immunoblotting
For determination of intracellular dengue virus protein, immunoblotting was performed using dengue envelope protein antibodies (Fig. 3). Immunoblot analysis revealed the presence of dengue virus envelope protein in THP-1 cells.
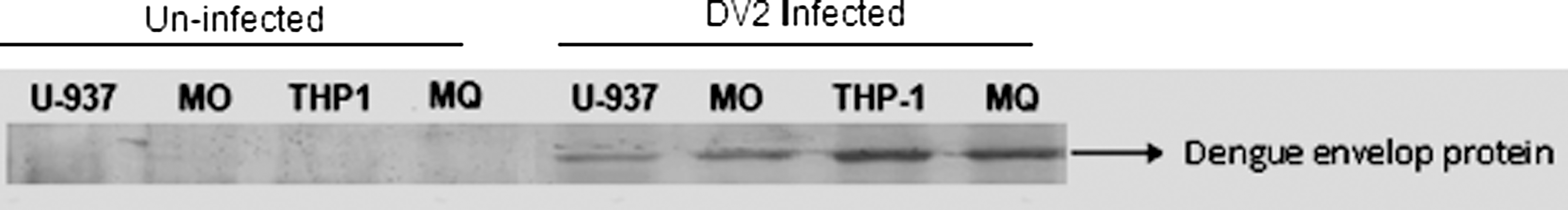
Immunoblot analysis revealing infectivity of dengue virus envelope protein in different cells. The four left lanes show non-infected control cells, and the four right lanes show dengue-virus-infected cells.
Expression of stress proteins during dengue virus infection
The expression of stress proteins during dengue virus infection in THP-1 cells was revealed by immunoblot analysis. The transferred proteins were probed with MAbs against Hsp27, Hsp60, Hsp70, and Hsp90. After 3 d of infection, whole-cell lysates were prepared and subjected to Western blot analysis. The data clearly indicated the predominant expression of Hsp70 compared to non-infected control cells (Fig. 4). There was no binding observed upon probing with Hsp27, Hsp60, and Hsp90 MAbs.
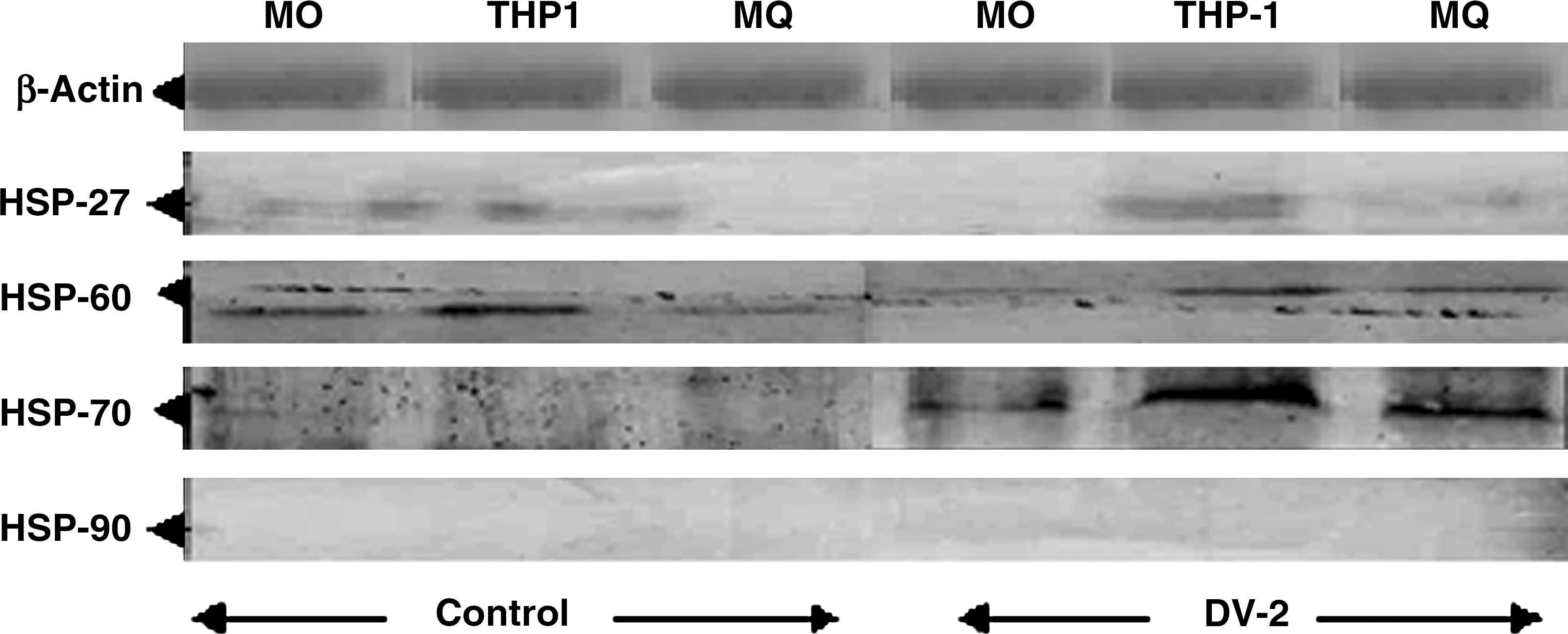
Expression of stress proteins during dengue virus infection in THP-1 cells after 3 d of infection. The rows show probing of blots with different MAbs against Hsps (row 1, β-actin; row 2, Hsp27; row 3, Hsp60; row 4, Hsp70; row 5, Hsp90; left lanes show blots of non-infected cells; right lanes show blots of dengue-virus-infected cells).
Downregulation of Hsp70 in THP-1 cells
To determine the functional role of Hsp70 in virus multiplication in dengue-virus-infected cells, expression of Hsp70 was downregulated by transfecting THP-1 cells prior to infection with specific siRNA duplexes using the gene silencing method. Western blot analysis revealed a marked decrease in Hsp70 expression in silenced and infected THP-1 cells (Fig. 5).

Immunoblot profile of downregulation of Hsp70 in THP-1 cells (C, control; CHS, control THP-1 cells exposed to a temperature of 42°C; CV, dengue-virus-infected cells; Cri, Hsp70-silenced non-infected cells; CRiV, Hsp70-silenced infected cells).
Effect of downregulation of Hsp70 on viral multiplication
Flow cytometric analysis was performed to measure the intracellular viral load in infected cells after silencing Hsp70. A significant decrease in intracellular viral load was observed in Hsp70-silenced dengue-virus-infected THP-1 cells (Fig. 6). Furthermore, real-time PCR analysis also confirmed a marked decrease in viral RNA copy number (Fig. 7).
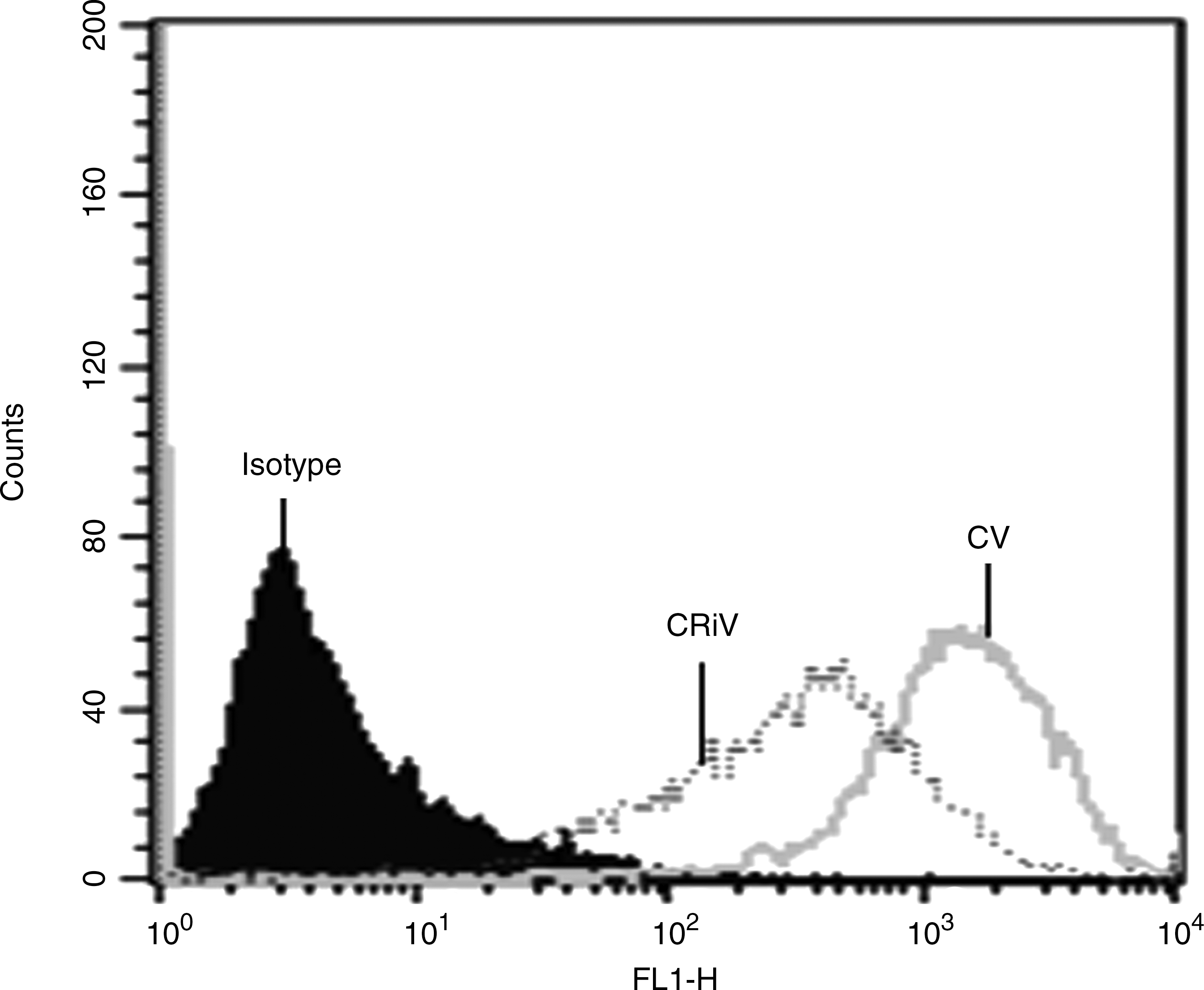
Determination of viral load in Hsp70-silenced infected THP-1 cells (CV, dengue-virus-infected cells; CRiV, Hsp70-silenced infected cells).
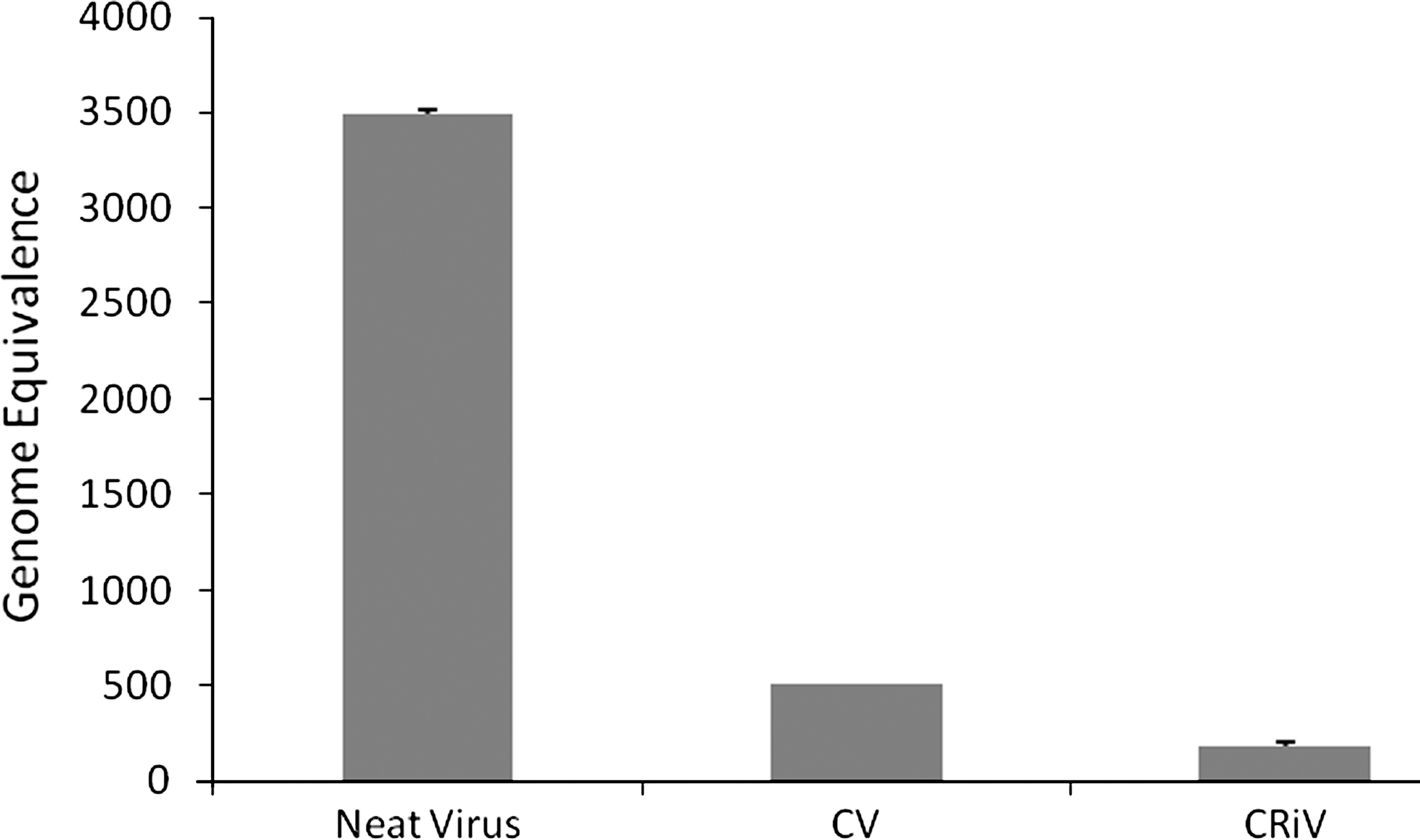
Determination of viral load in Hsp70-silenced dengue-infected THP-1 cells. Viral titers in cell supernatant as assessed by real-time PCR (CV, dengue-virus-infected cells; CRiV, Hsp70-silenced infected cells; neat virus was run as control).
Estimation of interferon-α in supernatants of Hsp70-silenced dengue-virus-infected cell culture
The effect of downregulation of Hsp70 in dengue-virus-infected THP-1 cells on the relevant cytokine production was determined by estimating the type I interferons. IFN-α was estimated in the supernatants of Hsp70-downregulated dengue-virus-infected THP-1 cells by commercially available kits. Fig. 8 shows that after downregulation, there was a significant increase in IFN-α production compared to controls. The increase in IFN-α production was inversely proportional to the viral load.
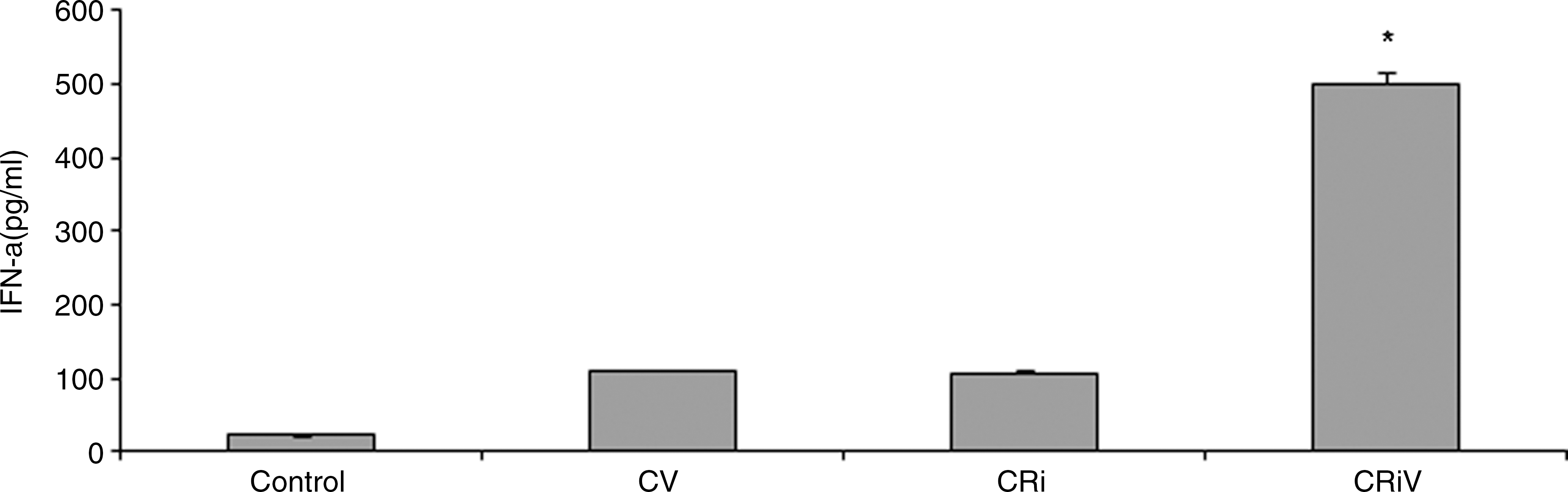
Interferon-α (IFN-α) production in Hsp70-silenced dengue-virus-infected THP-1 cells after 72 h of incubation (Control, non-infected cells; CV, dengue-virus-infected cells; Cri, Hsp70-silenced non-infected cells; CRiV, Hsp70-silenced dengue-virus-infected cells; *p < 0.01 versus CV).
Determination of PKR expression in Hsp70-silenced dengue-virus-infected cells
To determine whether the decrease in viral titer was due to an interferon-mediated antiviral mechanism via intracellular expression of double-stranded RNA-induced protein kinase (PKR) expression, immunoblot analysis was performed (Fig. 9). The results showed marked upregulation in the expression of PKR compared to controls in Hsp70-silenced dengue-virus-infected cells, confirming the interferon-mediated antiviral mechanism.

Immunoblot analysis of PKR expression in Hsp70-silenced dengue-virus-infected THP-1 cells after 72 h of incubation (C, control non-infected cells; CV, dengue-virus-infected cells; Cri, Hsp70-silenced non-infected cells; CRiV, Hsp70-silenced dengue-virus-infected cells).
Discussion
The identification of host cell factors that mediate dengue virus replication is an important, but as yet largely underexplored, area of research that will enable not only greater insight into the molecular mechanisms behind viral replication, but will also allow the identification of potential therapeutic targets (23). Some cellular and viral proteins, such as EF1-α, La, PTB, NS3, NS5, and some uncharacterized proteins (24,25), have been shown to bind to the 3′UTR of dengue virus, West Nile virus, and JE virus. Various studies suggest that Hsps may be involved in viral multiplication (18,26). In the present study Hsp70 was found to be overexpressed in dengue-virus-infected THP-1 cells, though it remains unclear at which stage of macrophage maturation dengue virus infects the cells. One of our earlier findings (19) suggests that Hsp60 is overexpressed in human promonocytic cell line U937. These observations constitute evidence of the induction of different stress proteins at different stages of dengue virus infection. The functional significance of these induced stress proteins remains unknown. In some viral infections they appear to be pro-viral and in others antiviral (27,28). However, it is becoming more likely that increased stress protein expression during viral infection is not simply part of the non-specific induction of host genes; it appears that these proteins may be utilized by the virus for replication and morphogenesis, or by the host cells as an intracellular defense against the invading microorganism (29). The interplay between viruses and stress proteins may affect the level of initial translation of the virus, as well as negative strand viral RNA synthesis, viral protein production, and virion secretion.
In a study by Iordanskiy et al. (28), they reported that Hsp70 induced during HIV infection in human macrophages inhibits nuclear translocation of HIV-1 viral protein R, suggesting a role for Hsp70 as an innate factor. In another study, Hsps were shown to play an important role in antibody-dependent entry of dengue virus into host cells. A study demonstrated that an 84-kDa molecule that was later characterized as a member of the Hsp90 family interacts with Hsp70 on the cell surface during LPS-induced cell signaling (17). Hsp90 and Hsp70 constitute the part of the receptor complex that was required for viral entry in neuroblastoma and monocytic cell lines, as well as in human monocytes and macrophages (17). These proviral and antiviral effects of Hsps during different types of viral infections are in accord with the results reported here, as we observed a proviral effect of Hsp70 after downregulation. The downregulation of Hsp70 by a siRNA-directed knockdown approach may help facilitate the study of gene function. In the present study, an siRNA-mediated gene silencing approach was used to determine the role of Hsp70 in viral multiplication. Overexpression of Hsp70 in dengue-virus-infected cells was downregulated by transfecting Hsp70-specific siRNA duplexes. The effect of Hsp70 downregulation on viral multiplication, intracellular viral load, and productive infection in cell supernatant was quantified, which revealed a reduced intracellular viral load and lower viral RNA copy numbers in Hsp70-downregulated virus-infected cells. A similar observation was made by Diamond et al. (30) in patient sera, as well as in infected cell supernatants in in-vitro viral assays, in contrast to the viral isolation seen in mosquito cells and plaque-formation assays (30). This suggests that Hsp70 is somehow involved in supporting viral multiplication, and thus indicates its proviral nature. This activity of Hsp70 is distinct from the mechanism described previously, which involves the binding of Hsp70 to viral complexes to facilitate antigen presentation and to enhance antiviral immunity (26).
Viral infection activates the transcription of a large number of cellular genes (31,32). Interferon regulatory factor (IRF)-3 is a key transcriptional activator, which is activated by dsRNA or by viral infection, and subsequently plays a role in transcriptional activation of IFN-α and IFN-β promoters, and IFN-α/IFN-β-responsive genes (33,34). It has also been reported using different cell types and viral strains that type I IFNs are involved in protection against dengue virus infection in vitro, in contrast to IFN-γ, which has a more variable effect, depending on the cell type and pathway of infection (30). In the present study, the proviral activity of Hsp70 was further confirmed by measuring antiviral cytokine type I IFN production. The cellular supernatants of Hsp70-downregulated cells after infection showed an increase in IFN-α production. This indicates a likely role of Hsp70 overexpression in inhibiting the IFN-mediated antiviral pathway during dengue virus infection.
PKR is a critical mediator of the anti-proliferative and antiviral effects exerted by IFNs. PKR is a serine/threonine-specific protein kinase that is activated by responses to double-stranded RNA, which involves autophosphorylation and also integrates signals in response to TLR activation, growth factors, and diverse cellular stressors (35,36). PKR activation affects both transcription and translation (37). This IFN-induced protein kinase can restrict viral replication through its ability to phosphorylate the protein synthesis initiation factor eukaryotic initiation factor-2α subunit, which reduces viral protein synthesis (38). Therapeutic strategies targeting PKR offer promising results. In the present study PKR expression was analyzed in Hsp70-downregulated dengue-virus-infected cells. High levels of PKR expression in these cells may be correlated with increases in IFN-α production. This indicates that type I interferon production in response to viral infection, in the absence of Hsp70, is exerting an anti-viral effect via induction of PKR expression, which may be responsible for viral translation.
The results of this study lead to the conclusion that elevated levels of Hsp70 expression in dengue-virus-infected THP-1 cells may aid viral multiplication, and represent a potential therapeutic target for the management of dengue virus infection.
Footnotes
Acknowledgements
K.P.M. and Y.S.P. contributed equally to this article. Y.S.P. thanks Defence Research and Development Organization for financial support in the form of junior and senior research fellowships. The authors thank Miss Ramanjot Kaur for technical assistance with the manuscript.
Author Disclosure Statement
No competing financial interests exist.