
Abstract
Influenza virus nucleoprotein (NP) binds to the viral genome RNA and forms the internal ribonucleoprotein complex of the virus particle. Avian and human influenza virus NP have characteristic differences at several amino acid positions. It is not known whether any of these differences can be recognized by antibodies. In the present study five monoclonal antibodies (MAbs) were produced against NP of A/Duck/Novosibirsk/56/05 (H5N1) influenza virus. Two MAbs discerned human and avian influenza strains on ELISA testing. The NP expressed in a prokaryotic system was used for the analysis of site-specific mutants carrying amino acid substitutions in the relevant positions. Amino acid residues in positions 100 and 101 were shown to be recognized by the MAbs. The residue in position 100 is host-specific, and its recognition by the MAb 2E6 may be useful for the differentiation of human and avian viruses. The data are discussed in view of the effects of amino acid substitutions in influenza virus NP affecting both host range and antibody-binding specificity.
Introduction
Influenza virus proteins, including NP, are variable, and some of the variations in NP are host-specific (9). Fifteen amino acid substitutions in NP were suggested as genomic signatures of human versus avian influenza A viruses (6), and in addition, four positions were identified as host-specific in the studies of the amino acid changes associated with transmission of avian influenza viruses to humans (20). In four positions, the introduction of a human-like amino acid residue in the genetic background of an avian virus resulted in a non-viable or attenuated progeny (12). However, the majority of host-specific positions are not absolute host signatures. Most of the avian-specific residues are also encountered in the viruses isolated from humans, although at a low frequency (6). Among such residues, those in positions 305 and 372 were identified in our studies as active in MAb-binding (26). This indicated that at least some of the amino acid residues in the host-specific positions are recognizable not only by host proteins, but also by antibodies.
In the present research we used anti-NP MAbs against A/Duck/Novosibirsk/56/05 (H5N1) avian influenza virus to identify the amino acid positions recognized by MAbs, and differing in mammalian and avian influenza A viruses. The amino acid changes were introduced into the NP molecule by site-specific mutagenesis in a prokaryotic system. The goal of the studies was to extend the knowledge of the location of areas in the 3D structure of influenza virus NP recognized by antibodies, and to find out whether the amino acid changes in the NP associated with the virus host range contribute to the reaction with MAbs.
Materials and Methods
Viruses
Influenza viruses were obtained from the virus collection of the D.I. Ivanovsky Institute of Virology, Moscow, Russia. The viruses were propagated in 10-day-old embryonated chicken eggs. The virus-containing allantoic fluid was stored at −80°C. For enzyme-linked immunosorbent assay (ELISA), virus was concentrated and purified by centrifuging the allantoic fluid at low speed, layering it on 20% (w/v) sucrose, and pelleting the virus by centrifugation at 23,000 rpm in an SW.27 rotor (100,000 g max) at 4°C for 90 min. The pellet was resuspended in Dounce homogenizer, clarified by low-speed centrifugation, aliquotted and frozen at −80°C.
Monoclonal antibodies
The preparation of the MAbs used in this study was described in our earlier publications (14,19). The MAbs 1G7, 1G8, 2E6, 4H4, and 2A5 were identified as anti-NP MAbs on the basis of Western blotting and competitive ELISA.
Serologic methods
Indirect ELISA was performed as described by Philpott et al. (17). Briefly, the antigen was bound to the plates at 4°C in 0.1 M carbonate buffer (pH 9.6). MAb in phosphate-buffered saline (PBS; Sigma-Aldrich, St. Louis, MO) supplemented with 0.05% Tween-20 and 0.5% bovine serum albumin (BSA) were added, and the plates were incubated for 1 h at 20°C, followed by three washes in the same solution without BSA. The plates were treated with goat anti-mouse immunoglobulin conjugated with horseradish peroxidase (HRPO; Sigma-Aldrich), and developed using TMB substrate (Sigma-Aldrich). Optical density (OD) was measured at a wavelength of 450 nm. The binding values were calculated using the equation: % binding = 100 × (Bxv/Bpv)/(Bxw/Bpw), where Bxv is the binding of a MAb to the test sample, Bpv is the binding of pooled MAbs to the test sample, Bxw is the binding of a MAb to the wild-type virus A/Duck/Novosibirsk/56/05 (H5N1) or expressed wild-type NP, and Bpw is the binding of pooled MAbs to the wild-type NP (17). For competitive ELISA the MAbs were purified by affinity chromatography on a Protein A-Sepharose CL-4B column (Pharmacia, Bridgewater, NJ), according to the manufacturer's instructions. The protein concentration was measured using a Bio-Rad Protein Assay kit (Bio-Rad, Hercules, CA), and the purified MAbs were conjugated with HRPO by the periodate method (15). The HRPO-conjugated MAbs in dilutions were incubated in wells coated with detergent-treated (17) virus, together with purified nonlabeled antibodies in a concentration of 100 μg/mL. The results were expressed as the degree of inhibition of the binding of the conjugated MAb in the presence of nonlabeled MAb. For Western blotting, samples of concentrated and purified virus were heated at 100°C for 2 min in a reducing sample buffer, and subjected to sodium dodecyl sulfate-12.5% polyacrylamide gel electrophoresis. The proteins were transferred to pure nitrocellulose membranes (Bio-Rad). The membranes were treated with 5% non-fat dry milk (Bio-Rad) in PBS overnight at 4°C, and incubated with MAbs diluted in 5% non-fat dry milk in PBS-0.05% Tween 20 for 1.5 h at 20°C. HRPO-labeled anti-mouse IgG (Sigma-Aldrich) were used for the detection of antibody binding. As a positive control a polyclonal rabbit immune serum against influenza virus core kindly supplied by Dr. M. Khristova (Centers for Disease Control and Prevention, Atlanta, GA) was used, and anti-rabbit HRPO-labeled IgG was used for the detection of the band. After washing, the membranes were developed with 0.05% 3,3′-diaminobenzidine blotting substrate (Sigma-Aldrich) in the presence of 0.01% hydrogen peroxide.
Cloning and expression of NP gene
The plasmid pET32b (Novagen, Gibbstown, NJ) was chosen as a vector for cloning and expressing the NP gene. cDNA copies of the NP gene were transcribed with RT primer Uni from the genomic RNA of A/Duck/Novosibirsk/56/05 (H5N1), A/Puerto Rico/8/34 (H1N1), A/Swine/Wisconsin/67 (H1N1), and A/Brisbane/59/07 (H1N1) viruses, and then amplified with the cloning primer pair NP_NdeI_F: 5′ AATTCATATGGCGTCTCAAGGCACC 3′ and Np(stKpn)R: 5′ ATAGGTACCTCAATTGTCGTACTCCTCTGC 3′. The PCR fragments were cloned into pET32b digested with restriction endonucleases NdeI and KpnI. Site-directed mutagenesis of the plasmids pET32b containing the wild-type NP genes was performed with a QuikChange MultiSite-Directed Mutagenesis Kit (Stratagene, La Jolla, CA), using specific oligonucleotide primers. Sequences of the primers used for site-directed mutagenesis and sequencing are presented in the supplemental data (Supplementary Table 1; Supplementary Data are available online at
+++, competition over 75%; ++, competition over 50% and less than 75%; +, competition over 25% and less than 50%; −, competition less than 25%.
Constructions containing wild-type and mutant NP sequences were expressed overnight in Escherichia coli strain B834 (DE3) co-transformed with pLysS, as recommended by the pET System Manual (Novagen). The T7 promoter was induced at 20°C with 0.5 mM IPTG when the OD 600 of the culture reached 0.6 optical units. Cells from a 200-mL overnight culture were resuspended in 10 mL of phosphate-buffered saline and broken by sonication. The supernatant obtained from centrifuging the cell lysate was used in ELISA testing.
Results
Anti-NP MAbs
Among 16 hybridomas against A/Duck/Novosibirsk/56/05 (H5N1) virus (14), 5 hybridomas were shown to produce MAbs against viral NP protein. Three of them (1G7, 2E6, and 2A5) reacted with NP in Western blotting (Fig. 1), and two others, 1G8 and 4H4, were competitive with 1G7 and 2E6, respectively, in competitive ELISA (Table 1). All anti-NP MAbs were shown in our earlier studies (14) to belong to the IgG1 subclass. The results of Western blotting indicated that the MAb 2A5 reacted with a linear epitope, whereas the MAbs 1G8 and 4H4 recognized conformational epitopes (Fig. 1). The MAbs 1G7 and 2E6 produced a weak band in Western blotting. This result may be interpreted either as a weak affinity reaction with a linear epitope, or as a result of partial refolding of a discontinuous epitope (30). The data from competitive ELISA revealed three operational non-overlapping MAb-binding epitopes (Table 1), one recognized by MAbs 1G7 and 1G8, another by 4H4 and 2E6, and the third by 2A5. The overlapping between the MAbs recognizing the same epitope was either complete (2E6 versus 4H4), or partial.
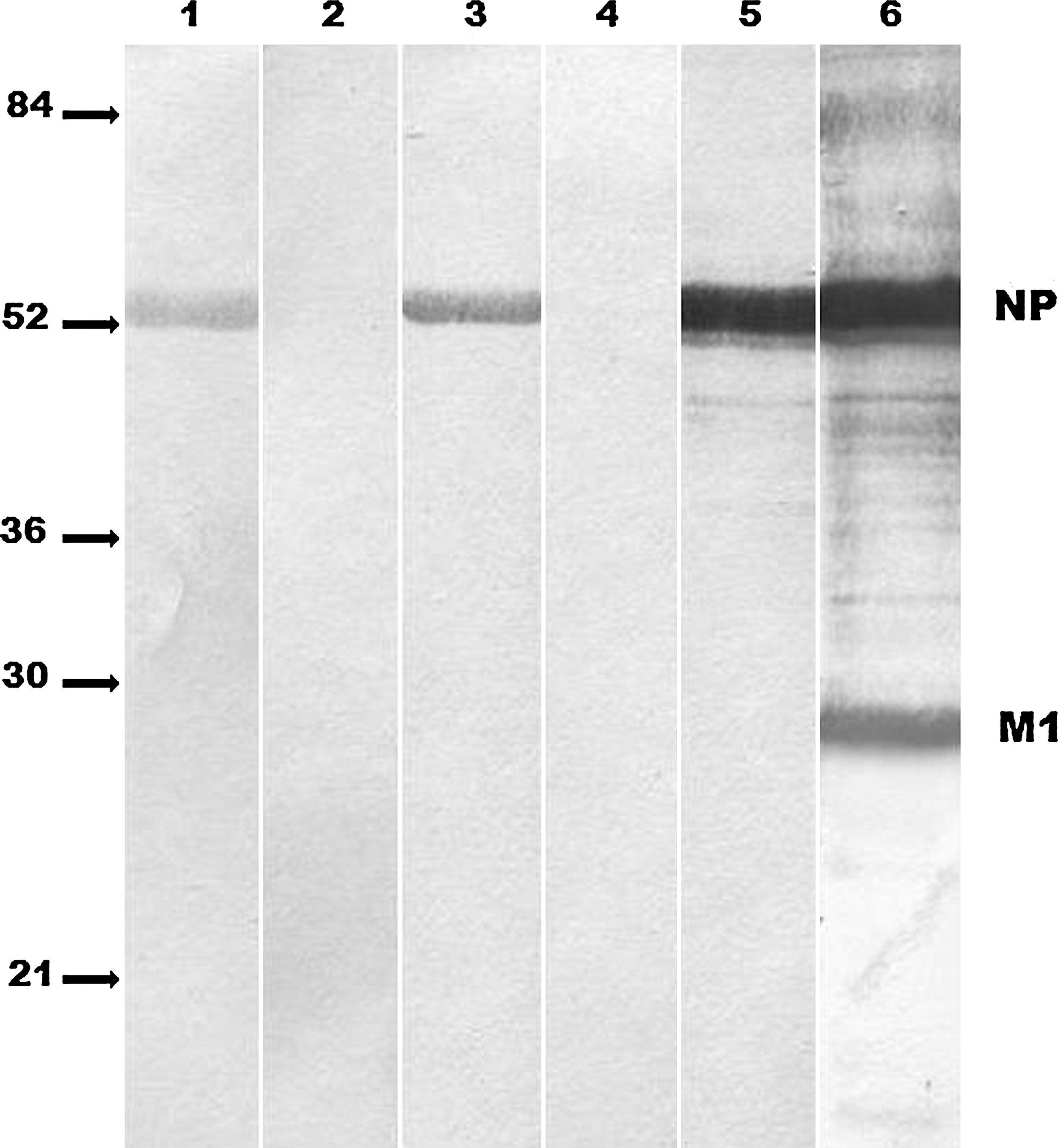
Western blotting of A/Duck/Novosibirsk/56/05 (H5N1) virus with anti-NP MAbs. Lanes 1, 2, 3, 4, 5, 6 show MAbs 1G7, 1G8, 2E6, 4H4, and 2A5, and polyclonal rabbit immune serum against virus core, respectively. Viral proteins are indicated on the right. Positions of molecular mass markers are indicated on the left (NP, nucleoprotein).
Reaction of anti-NP MAbs in ELISA with avian and mammalian influenza A viruses
The anti-NP MAbs were used in indirect ELISA with avian and human viruses, and with one classical swine virus. Each MAb was titrated in ELISA against the homologous virus, and used in a saturating concentration in further experiments. The results of ELISA with purified preparations of influenza A viruses (Table 2) revealed that all tested viruses reacted with the MAbs 1G7, 1G8, and 2A5, and the avian viruses reacted also with the MAbs 2E6 and 4H4. The binding as calculated by the equation of Philpott et al. (17) (see Materials and Methods section) varied from 52% to 100%. The mammalian viruses failed to react with the MAbs 2E6 and 4H4 (binding less than 5%).
+, binding over 50%; −, binding less than 5%.
ELISA, enzyme-linked immunosorbent assay.
The inspection of the amino acid sequence of NP proteins of the viruses used in ELISA revealed 20 positions among those exposed at the surface of the NP molecule in an x-ray 3D model (16,29), where the amino acid residues were uniform in the NP of all avian viruses tested, whereas amino acid substitutions occurred in at least some mammalian strains (Table 3). Sixteen of them were recognized in earlier studies (6,20) as host-specific. We added positions 101, 351, 421, and 423. The amino acid residues in these positions, like in the ones recognized earlier as host-specific, were uniform in the avian viruses tested, and varied in mammalian viruses.
Reaction of MAbs with site-specific mutants
Data from the comparative analysis of avian and mammalian influenza virus strains in ELISA were used to choose the mutations to be introduced into the plasmid expressing the NP protein of A/Duck/Novosibirsk/56/05 (H5N1) virus. Sixteen positions were chosen, 12 of them recognized as host-specific in earlier studies (6,20). We chose those positions where the amino acid residues were uniform in the viruses reacting with the MAbs 2E6 and 4H4. In the majority of these positions one or several mammalian strains have avian-like residues. We considered that such positions have to be tested, because the absence of the reaction of such strains with the MAbs 2E6 and 4H4 can result from the presence of human-like residues in other positions.
The mutant proteins were expressed and analyzed with ELISA (Table 4). Each antigen was titrated against the mixture of MAbs, and the minimal saturating concentration was used in the reactions with individual MAbs. The results revealed that the amino acid substitution R100V abolished the reaction with the MAbs 2E6 and 4H4. The substitution D101N abolished the reaction with 4H4. The other amino acid changes had no effect on the reaction.
+, binding over 50%; −, binding less than 5%.
We attempted to introduce the amino acid substitutions V100R and N101D into the plasmid-expressed NP of mammalian influenza viruses in order to make them reactive with the MAbs 2E6 and 4H4. The introduction of the V100R substitution into the NP of A/Brisbane/59/07 (H1N1) and A/Swine/Wisconsin/67 (H1N1) viruses led to the acquisition of the ability of their NP to react with 2E6 and 4H4 MAbs. Both viruses have D101, that is, they do not differ from the avian viruses by the amino acid residue in position 101. The NP of A/Puerto Rico/8/34 (H1N1) virus after the introduction of the V100R substitution acquired the ability to react with 2E6, but not with 4H4. The introduction of the second substitution, N101D, made A/Puerto Rico/8/34 (H1N1) virus reactive with both MAbs (Table 4).
We did not introduce amino acid substitutions into the plasmid expressing the NP of A/Duck/Novosibirsk/56/05 (H5N1) virus in positions 293, 305, 372, and 455, although initially we planned to obtain such mutants. These amino acid residues are both host-specific and exposed at the surface of NP molecule. However, in any of these positions the NP of A/Puerto Rico/8/34 (H1N1), A/Brisbane/59/2007 (H1N1), and A/Swine/Wisconsin/67 (H1N1) has a human-like residue. Since these residues do not preclude the reaction of the MAbs 2E6 and 4H4 with NP after the introduction of the V100R substitution in the NP of A/Brisbane/59/07 (H1N1) and A/Swine/Wisconsin/67 (H1N1), or V100R and N101D substitutions in A/Puerto Rico/8/34 (H1N1), it seems obvious that the human-like amino acid residues in positions 293, 305, 372, and 455 do not preclude the reaction of 2E6 and 4H4 MAbs with NP. Therefore we regarded as unnecessary the production of site-specific mutants with human-like residues in these positions.
Positions of amino acid residues recognized by MAbs in the 3D structure of NP
The results of site-specific mutagenesis and ELISA with expressed mutant NP allowed us to identify amino acid positions 100 and 101 as recognized by MAbs against the NP of an avian influenza virus. Mapped in the 3D model of NP (29), the site represented by amino acid residues 100 and 101 is located far from the antigenic sites defined by positions 236, 305, 372, and 470 found in our previous studies (26) (Fig. 2). The amino acid positions 100 and 101 are adjacent in the amino acid sequence, and quite expectedly, in 3D structure of NP (Fig. 2). The results of the reaction of site-specific mutants with the MAbs suggest that the MAb 4H4 recognizes both positions, whereas 2E6 recognizes only the residue in position 100. This interpretation is consistent with the data from competitive ELISA (Table 1). The results demonstrate the ability of MAbs to discern subtle changes in the structure of the influenza virus NP molecule.
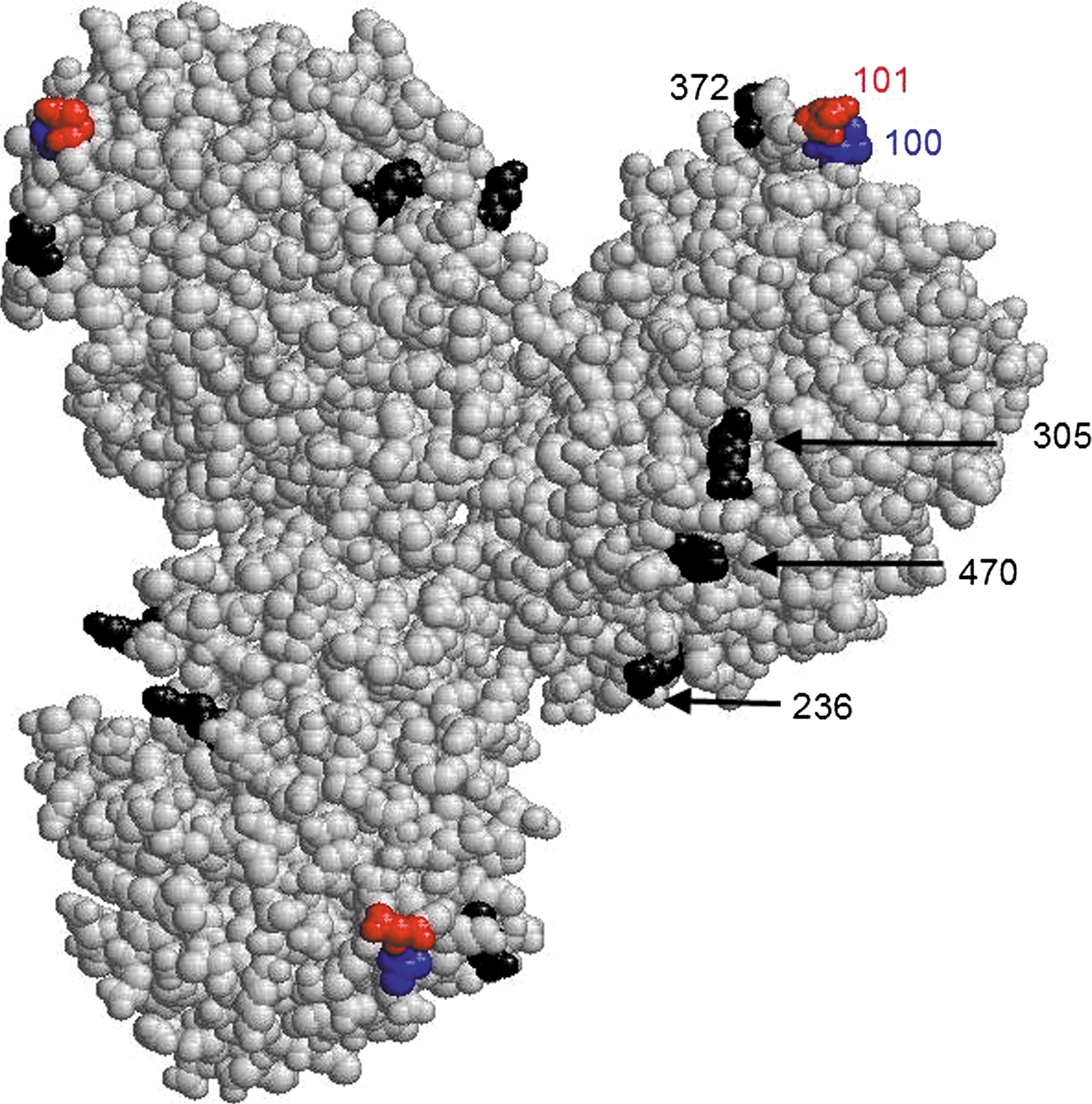
Location of amino acid positions involved in the reaction with monoclonal antibodies (MAb) on the three-dimensional structure of the nucleoprotein (NP) molecule. Images were created with RasMol 2.6, and NP structure was obtained from the Protein Data Bank (PDB accession number 2IQ6). NP is presented as a trimer (29). The amino acid numbers are indicated in one monomer. The amino acid positions defined as MAb-binding in this study are marked in red and blue, and the positions identified as antigenic in our earlier report (26) are marked in black.
Discussion
Influenza A viruses are circulating in many species of birds and mammals. Although from time to time a virus crosses an inter-species barrier, on the whole the influenza A viruses are adjusted to propagation in a specific host. Specific amino acid positions in virus proteins have been recognized as “genomic signatures” distinguishing the proteins of human influenza viruses from those of avian influenza viruses (6,7,20). Host-specific amino acid residues differing in the NP of influenza viruses of water birds, gulls, horses, humans, and pigs were recognized two decades ago (9). A correlation between the host range of influenza viruses and the reaction of their NP with MAbs was also described (10). However, to our knowledge, this report is the first in which an amino acid position belonging to host-specific “genomic signatures” of NP has been identified as a part of a MAb-binding site.
The amino acid positions 100 and 101 identified in this study as a part of the same MAb-binding site differ in their relation to host specificity. Position 100 is one of the most strictly defined “genomic signatures.” Basic amino acids in this position are dominant in the NP of avian viruses, and they never occur in human viruses (6). Therefore, the recognition of this amino acid by the MAb 2E6 may be useful for the differentiation of human and avian viruses. On the other hand, D101 is obligatory in avian viruses, and frequent in human and classic swine viruses. In this aspect position 101 resembles positions 236, 305, 372, and 470, which were described in our earlier studies (26) as discernible by the MAbs to A/WSN/33 (H1N1) virus. Amino acid residues in each of these positions are almost uniform in avian viruses and variable in human viruses. Two of them, 305 and 372, were listed as “genomic signatures” (6). However, in fact these two residues are avian-like in the early human H1N1 viruses. Substitutions at positions 236 and 470 do not exhibit such a correlation with the time of strain isolation (26), as well as the change in position 101 identified in this study as affecting the binding of a MAb.
Position 100 has been recognized as host-specific and phenotypically significant not only on the basis of comparative sequence studies, but also on the basis of direct reverse-genetic experiments. The mutant virus carrying the change of avian-like residue R100 to a mammalian-like V100 or I100 in the avian background, could not be rescued, that is, the amino acid change resulted in non-viability of the mutant (12).
The mechanism of the effect of most host-specific amino acid changes in influenza A virus proteins, including the changes in position 100 of NP, is not known. Position 100 is located in the N-terminal part of NP. It has been shown that this part is involved in the interaction of NP with RNA (1), with PB2 (2), and with MxA protein (22). The interaction with MxA occurs through direct protein-protein contact (22). Recently this interaction was shown to be species-specific (32). The data presented in this report suggest that the amino acid change in position 100 is recognized both by antibodies and by host-specific functional cellular proteins. Position 101 is not host-specific, but it may be biologically significant. The substitution D101N was registered in a revertant of a cold-adapted attenuated influenza virus strain (21). Recently, amino acid change D101G was shown to occur in the course of the adaptation of the 2009 pandemic H1N1 virus to mice (11). It seems plausible that position 101, although not listed as a human versus avian genomic signature (6, 20), is important for host range and other phenotypic features, besides its effect on the reaction with MAb.
The approach used in our studies is based on the introduction of site-specific mutations in the positions that are variable in the primary structure. The identification of such positions as MAb-binding by no means implies that they are the only ones involved in the NP-antibody interactions. We identified either one amino acid position per antigenic site (26), or two positions per site (this study). In fact an antibody molecule usually interacts with several amino acid residues. To perform a search for invariant amino acid positions recognized by antibodies, an alternative approach will have to be developed, not involving the strain-specific amino acid differences.
Footnotes
Acknowledgments
The authors are grateful to Dr. Leslie Meenderink of Vanderbilt University for critical reading of this article. This study was supported by grant 10-04-00023 of the Russian Foundation for Basic Research (RFBR).
Author Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.