
Abstract
T helper-1 (Th-1)-type immune responses play an important role in viral clearance during infection with hepatitis B virus (HBV). Unmethylated CpG motifs present in bacterial DNA can activate toll-like receptor 9 (TLR9) signals and act as potent adjuvants to induce Th-1-type immune responses. Here, a mini-plasmid with 812 base pairs in length was constructed and used as a vector to prepare a series of plasmids containing 3–21 copies of D-type CpG motifs. In vitro, these CpG-enriched plasmids strongly stimulated proliferation of human peripheral blood mononuclear cells (PBMCs) and enhanced secretion of interferon-γ (IFN-γ) and interleukin-12 (IL-12). The responses of the PBMCs from healthy individuals to the plasmids were stronger than those obtained from HBV-infected individuals. Contrary to the strong Th-2-biased response induced by surface antigen of hepatitis B virus (HBsAg) plus alum adjuvant, immunization of BALB/c mice with HBsAg plus these plasmids induced a strong Th-1-biased response. The plasmids increased the titers of HBsAg-specific total immunoglobulin G (IgG) and IgG2a. HBsAg-specific IL-2 and IFN-γ production and cytotoxic activity were also enhanced in the presence of the plasmids. The strength of the immune responses positively correlated with the number of CpG motifs in the plasmids. These results indicate that the use of CpG-enriched plasmids as an adjuvant to recombinant HBsAg could provide a promising and cost-effective approach for the development of efficacious therapeutic vaccines against HBV infection.
Introduction
Unmethylated CpG motifs present in bacterial DNA stimulate innate host defense mechanisms by activating toll-like receptor 9 (TLR9) signaling pathways (5 –7). Synthetic oligodeoxynucleotides (ODNs) containing CpG motifs have been shown to act as potent adjuvants that can induce Th-1 immunity by mimicking the effects of bacterial DNA (8 –11). ODNs can improve immune responses to a variety of co-administered antigens, and therefore represent a promising new approach for improving vaccine efficacy (12 –14). Recently, ODN CpG 7909 has been shown to enhance the immunogenicity of various microbial antigens and tumor antigens in humans (15 –18). CpG motifs present in the backbone of DNA vaccines play an important role in the development of immune responses, and increasing the number of the motifs could enhance immune responses to target antigens (19 –22). Compared with synthetic ODNs, plasmids have more stable chemical properties and are economical to prepare in large quantities. Hence, CpG-enriched plasmids may prove to be more useful than protein or peptide vaccine adjuvants.
Th-1-type immune responses play a critical role in virus clearance during acute or chronic HBV infection (23 –25). HBV vaccines have recently taken on a new role as therapeutic vaccines in attempts to cure or to control HBV infection in persistently-infected individuals (26 –29). Aluminum is the adjuvant of choice in commonly used commercial HBV vaccines; however, it is a Th-2-response-stimulating adjuvant, and therefore does not meet current demands for use in a therapeutic vaccine (30,31). Thus it is desirable to identify an adjuvant that promotes Th-1-type responses.
In the present study we constructed a series of plasmids containing multiple copies of CpG ODNs to investigate their effect on the immune function of PBMCs from healthy and HBV-infected adults. We also immunized BALB/c mice with HBsAg in the presence or absence of the plasmid constructs, and measured their Th-1 immune responses to determine the potential of the plasmids as vaccine adjuvants for the treatment of hepatitis B infection.
Materials and Methods
Construction of plasmids for carrying CpG ODN sequences
The plasmid pUC18 was used as a template to amplify a fragment containing the DNA replication origin. The primers used were: Sense: 5′-
Construction of CpG ODN-enriched plasmids
An ODN that contains three copies of CpG motifs has been described previously (21). Single-stranded sense and antisense ODNs comprising the following sequences were used. Sense: 5′-GATC
Chronically HBV-infected subjects and healthy subjects
Eight male HBV-infected subjects, aged 28–34 y, with at least 3 y of HBV infection history were enrolled into the study. All of the subjects were HCV and HIV negative (by ELISA method) and had not previously received any antiviral treatment. HBV DNA levels in the plasma of the subjects ranged from 1.5 × 106 to 4.6 × 108 copies/mL, and ALT levels in the plasma were normal (<40 U/L). All healthy adult subjects (n = 8; age-, sex-, and race-matched) had no evidence of HBV, HCV, or HIV infection (as determined by ELISA tests). Informed consent was obtained from each subject and the experiments were conducted in accordance with the Institutional Review Board guidelines for the protection of human subjects. Heparinized peripheral blood samples were collected on the same day from all of the subjects and used to measure the stimulatory activity of the CpG plasmids.
PBMC isolation and stimulation using CpG plasmids
PBMCs were isolated from heparinized peripheral blood samples using Ficoll-isopaque gradient centrifugation and suspended in RPMI 1640 culture medium supplemented with 10% heat-inactivated fetal bovine serum (FBS) (Invitrogen Corp., Carlsbad, CA), 1.5 mM L-glutamine, 100 U/mL penicillin, and 100 mg/mL streptomycin. PBMCs (1 × 105/well) were incubated in quadruplicate for 3 d at 37°C in 96-well round-bottom plates with or without plasmids at a final concentration of 10 μg/mL. Cell proliferation was assayed using the CellTiter 96 AQueous One Solution. The stimulation index (SI) was determined as described previously (33). PBMCs at a concentration of 2 × 106/mL were cultured in complete RPMI 1640 medium in the presence of various plasmids (final concentration 10 μg/mL) in a 5% CO2-humidified incubator at 37°C. After 48 h, the cytokine levels of the supernatants were measured using commercially available ELISA kits for IFN-γ and IL-12 p40 (Biosource, Nivelles, Belgium).
Mouse immunization
Recombinant HBsAg (subtype ayw, produced in Saccharomyces cerevisiae) was obtained from Shenzhen Kangtai Biological Products Co. Ltd., Shenzhen, China. Groups of 8-wk-old female BALB/c mice (SIPPR-BK Experimental Animal Co. Ltd., Shanghai, China) were intramuscularly injected with 0.1 μg HBsAg supplemented with 25 μg of the CpG plasmids, or 0.1 μg HBsAg formulated with aluminum hydroxyphosphate sulfate adjuvant (Merck, Darmstadt, Germany). Chicken ovalbumin (OVA; Sigma-Aldrich, Co., St. Louis, MO), used as a control antigen, was injected in combination with 25 μg plasmid pCG3 (containing three copies of CpG motifs). Each group consisted of eight mice in total. All groups received a booster immunization 2 wk after the first was administered.
Serum antibody assay
ELISA plates were coated with 1 μg of recombinant HBsAg per well overnight. The plates were blocked with milk buffer (5% nonfat dry milk and 0.05% Tween 20 in 1× PBS) for 1 h and incubated with diluted mouse sera for 40 min. The plates were extensively washed before incubating with horseradish peroxidase (HRP)-conjugated goat anti-mouse IgG, IgG1, or IgG2a (Southern Biotechnology Inc., Birmingham, AL) diluted 1:2000 in milk buffer for 40 min, after which the plates were washed and the color developed using 3,3′,5,5′-tetramethylbenzidine as the substrate. The absorbance of the samples was measured at 450 and 630 nm. End-point titers were defined as the highest sera dilution that resulted in an absorbance value (OD 450) two times greater than that of mice immunized with OVA plus the pCG3 plasmid. Anti-HBs titers for the groups of animals were expressed as the geometric mean of the titers ± the standard error of the mean (GMT ± SEM) of the values of the individual animals.
Cytokine assay
Three mice per group were humanely killed 1 wk after the booster immunization, and the spleens were removed under aseptic conditions. Single-cell suspensions were prepared by gentle passage of the tissue through sterile screens. Erythrocytes were lysed with Tris-buffered ammonium chloride, and the remaining cells were washed extensively in RPMI 1640 medium containing 2% FBS. Splenocytes were suspended at 5 × 106 cells/mL in RPMI 1640 supplemented with 5 × 10−5 M 2-β-mercaptoethanol, 10% heat-inactivated FBS, and 0.3 mg/mL L-glutamine. The cells were cultured at 200 μL/well in 96-well plates in the presence or absence of HBsAg (20 μg/mL). After 72 h, the cell-free culture supernatant was collected from each well and IL-2, IL-4, and IFN-γ levels were measured using ELISA kits (Biosource) according to the manufacturer's instructions.
Cytotoxicity assay
SP20/S cells derived from SP2/0 cells that constantly express HBsAg were used as stimulator or target cells. SP2/0 cells were used as controls. Effector cell populations were prepared by culturing 3 × 107 splenocytes with 1 × 106 irradiated SP20/S cells. Co-culture of the cells was performed in 10 mL RPMI 1640 medium supplemented with 10 mM HEPES buffer, 5 × 10−5 M 2-ME, 10% FCS, and recombinant mouse IL-2 (5 U/mL). After 5 d, the cultured spleen cells were purified with Ficoll and used as effector cells to determine the level of cytolytic activity (CTL). Serial dilutions of the effector cells were cultured with 2 × 103 SP20/S target cells in 96-well round-bottom plates and incubated for 6 h at 37°C with 5% CO2. HBsAg-specific CTL activity was measured by lactate dehydrogenase (LDH) release using the non-radioactive cytotoxicity assay kit (Cytotox 96; Promega Corp., Madison, WI) as described previously (33). The OD 492 absorbance values of the culture supernatants were measured on an ELISA microplate reader, and cell-specific lysis was calculated according to the following formula: (experimental release – spontaneous release) / (total release – spontaneous release) × 100%. In this study, the term “experimental release” represents the LDH released by target cells in the presence of effector cells. “Total release” represents the LDH released after total lysis of target cells with 5% Triton X-100. Spontaneous release was determined using target cells incubated without effector cells.
Statistical analysis
The data were analyzed for normal distribution, and the statistical significance of the difference between groups was determined by the two-tailed unpaired Student's t-test. In this study, p < 0.05 was considered to be statistically significant.
Results
Plasmid construction
An 812-bp plasmid (pOS) was used as a vector for CpG ODNs. The construct contained a replication origin (567 bp) derived from pUC18, and a tyrosine tRNA suppressor gene (227 bp) for antibiotic selection in E. coli harboring a P3 plasmid (Fig. 1). BamHI and EcoRI restriction sites were used for cloning the synthetic ODNs.
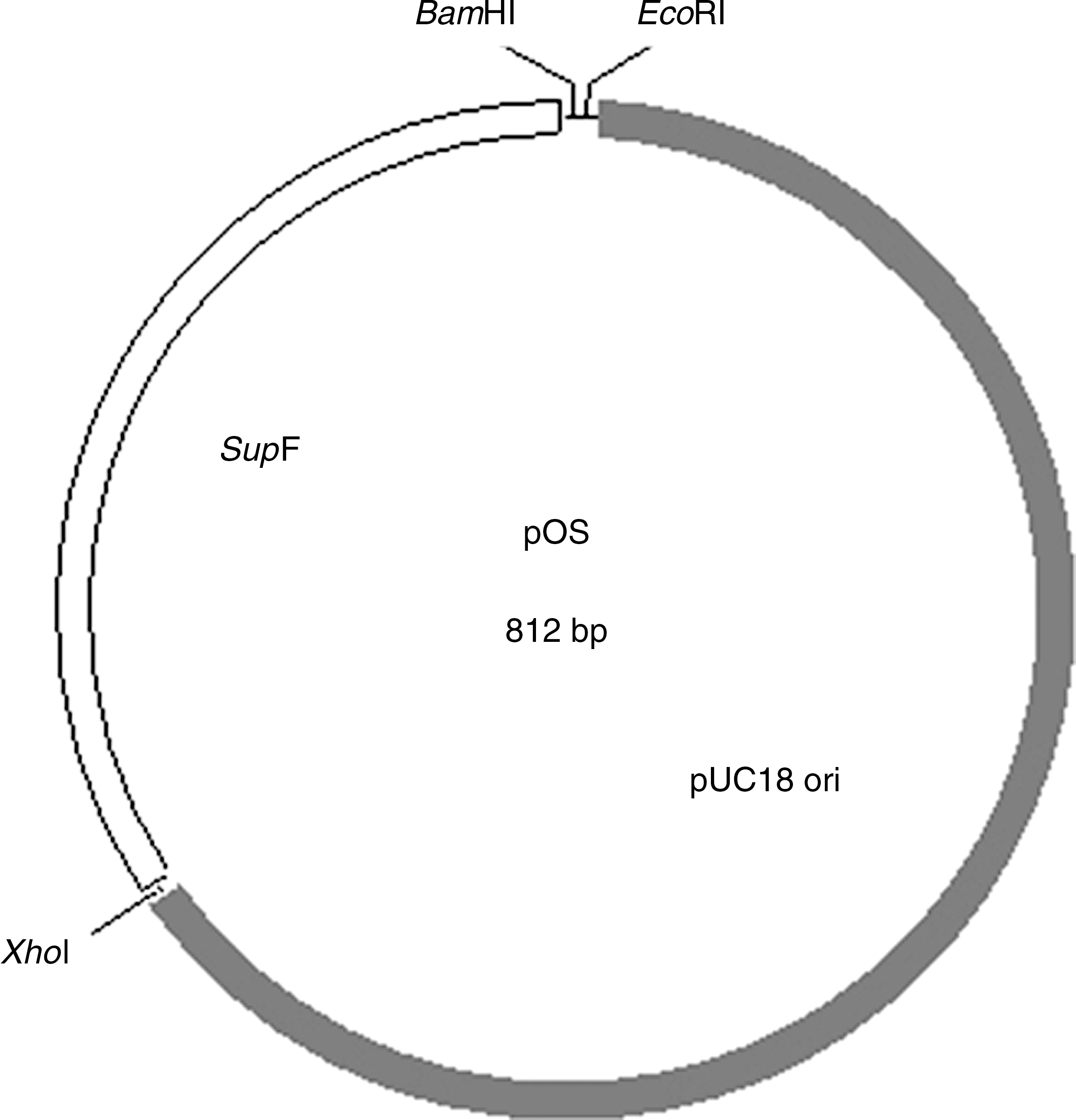
Structure of the pOS plasmid. The plasmid used for carrying CpG ODNs (pOS) was constructed as described in the materials and methods section. It contains two functional elements, a pUC replication origin, a tyrosine tRNA suppressor (supF gene), and three restriction sites.
CpG plasmids enhance proliferation and cytokine secretion of PBMCs from healthy and HBV-infected adults
A series of plasmids containing 3–21 copies of CpG motifs were used to investigate their influence on the cell proliferation and cytokine production of PBMCs obtained from healthy and HBV-infected adults. Another plasmid, pOS-woCG, containing a single copy of the synthetic ODN lacking CpG motifs, was used as a control.
The plasmids pOS and pOS-woCG (containing an ODN but lacking CpG motifs) showed a similar degree of enhancement of proliferation of PBMCs isolated from healthy or HBV-infected adults (compared with cells that had not been treated with plasmid; p < 0.01). Compared with those treated with pOS or pOS-woCG, cells treated with plasmid pCG3 (containing three copies of the CpG motifs) showed much stronger proliferation profiles (p < 0.01; Fig. 2). Fig. 2 also shows that the SI was positively correlated with the number of CpG motifs present in the plasmids. Increasing the number of CpG motifs resulted in an enhancement of PBMC proliferation overall. It is noteworthy that the responses of the PBMCs from healthy individuals to the CpG plasmids were constituently stronger than those from the HBV-infected subjects. We also noted that when the CpG motifs exceed 15 and 18 copies, the proliferation of PBMCs of healthy or HBV-infected subjects reached a maximum level, but increasing the number of CpG motifs did not increase PBMC proliferation further.
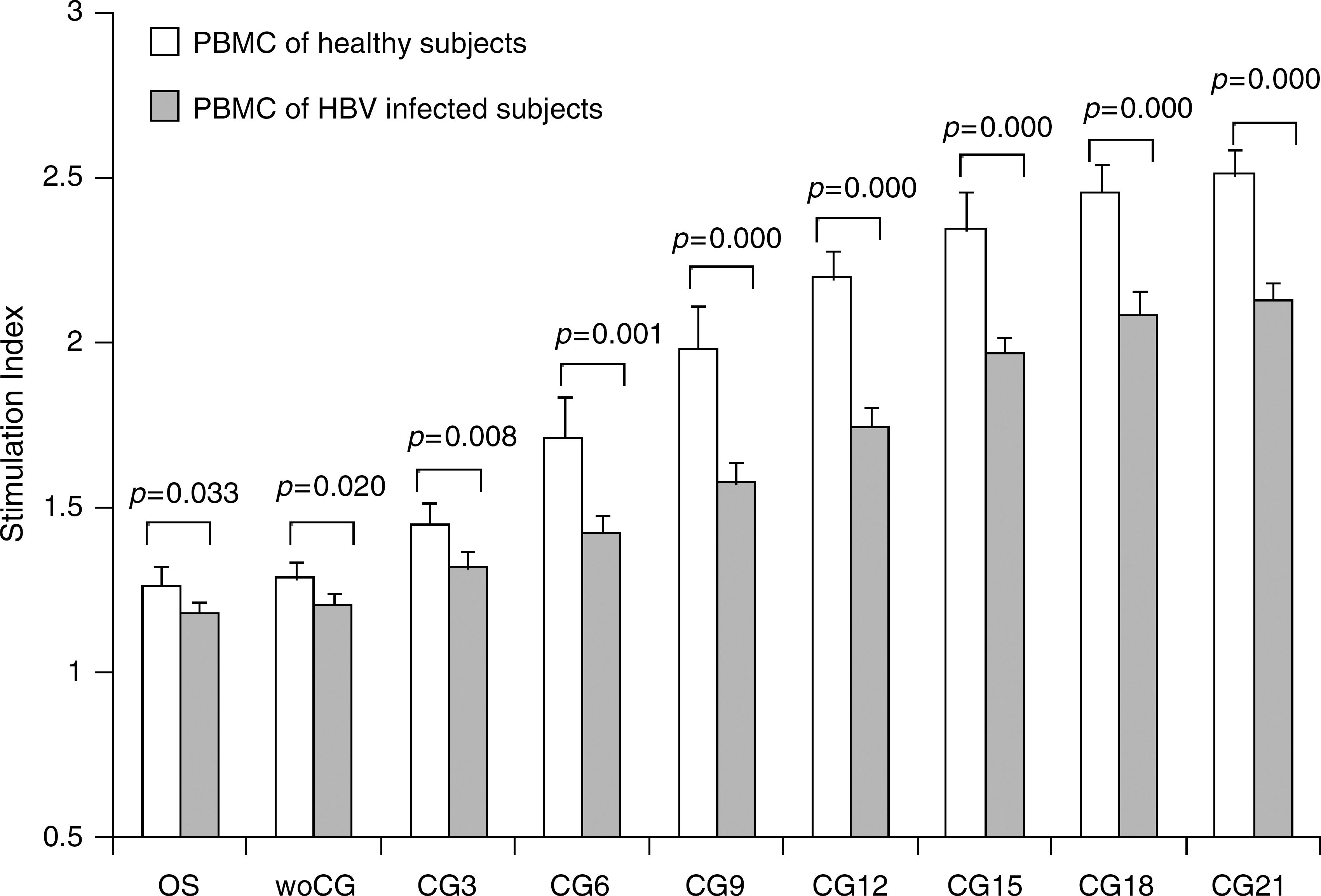
Proliferation of human PBMCs treated with CpG plasmids. PBMCs (1 × 105/well) were cultured in 96-well plates for 3 d with various plasmids at a concentration of 10 μg/mL. Cell viability was assayed using MTS and PES reagents. The data shown are the mean SI of four replicates, and are representative of three independent experiments.
We measured the levels of IFN-γ and IL-12 cytokines from the culture supernatants. Compared with control cultures (that had not been treated with plasmids), cultures treated with pOS or pOS-woCG showed a significant enhancement of IFN-γ and IL-12 secretion (p < 0.001) (Fig. 3A and B). Treatment with the pCG3 plasmid resulted in a significant increase in production of both cytokines (p < 0.001, compared with pOS- or pOS-woCG-treated cells). Measurements of the level of IFN-γ secreted by PBMCs from healthy or HBV-infected subjects showed that when CpG motifs in the plasmid exceeded 12 (healthy) and 18 copies (HBV infected), the levels of enhancement reached a maximum. A similar trend was also observed for IL-12 secretion by PBMCs from healthy or HBV-infected subjects; we observed that enhancement levels reached a maximum when CpG motifs exceeded 12 (healthy) and 18 (HBV infected) copies. Treatment of PBMCs derived from healthy subjects with any one of the plasmids produced higher levels of both cytokines (IFN-γ and IL-12) than those obtained from the HBV-infected subjects.
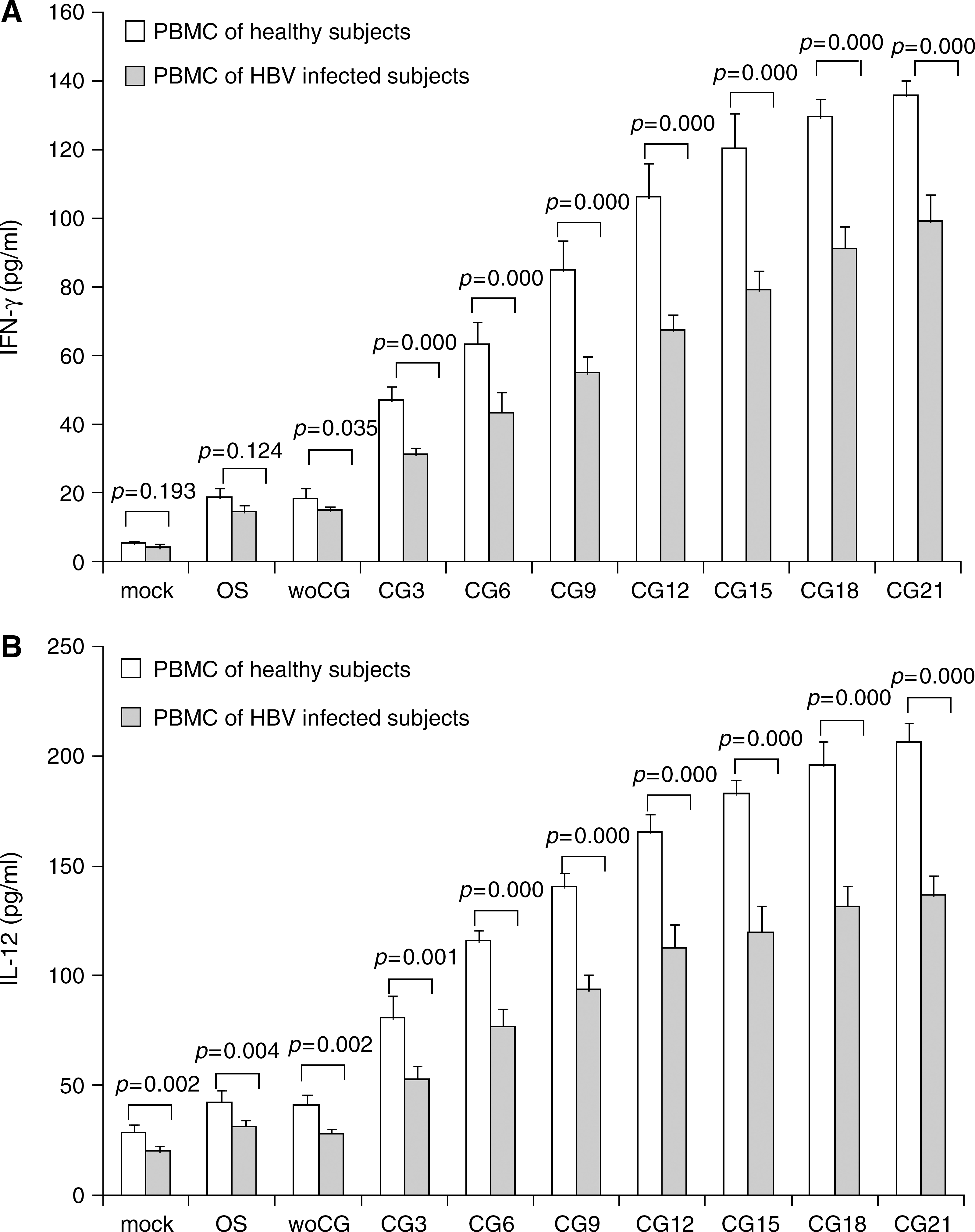
Cytokine secretion of human PBMCs treated with CpG plasmids. PBMCs (5 × 106/mL) were cultured in 24-well plates with various plasmids at concentrations of 10 μg/mL. After 48 h, supernatants were collected and assayed for IFN-γ (
CpG plasmids enhance the production of anti-HBs total IgG and IgG2a in mice
Based on our observations that CpG plasmids activated human PBMCs, we assessed the ability of these plasmids to stimulate immune responses to HBsAg in mice. As shown in Fig. 4A, primary vaccination using 0.1 μg HBsAg alone induced relatively low levels of anti-HBs IgG. Immunization with HBsAg plus alum adjuvant induced much higher levels of anti-HBs IgG (p < 0.001). The antibody titers of mice immunized with HBsAg plus plasmid pOS were higher than those in mice immunized with HBsAg alone (p < 0.05), but lower than those in mice immunized with HBsAg plus alum adjuvant (p < 0.01). When used in combination with the pCG3 plasmid, the titers increased to comparable levels with the alum adjuvant group, and the titers rose further when plasmids with increasing numbers of CpG motifs were used (OS versus CG3, p < 0.01; CG3 versus CG9, p < 0.05). The antibody titers did not increase markedly when plasmid CG15 or CG21 was co-administered (CG9 versus CG21, p > 0.05). After the booster immunization, the antibody titers of all the groups tested increased significantly, while the antibody differences within the groups did not change throughout the observation period.
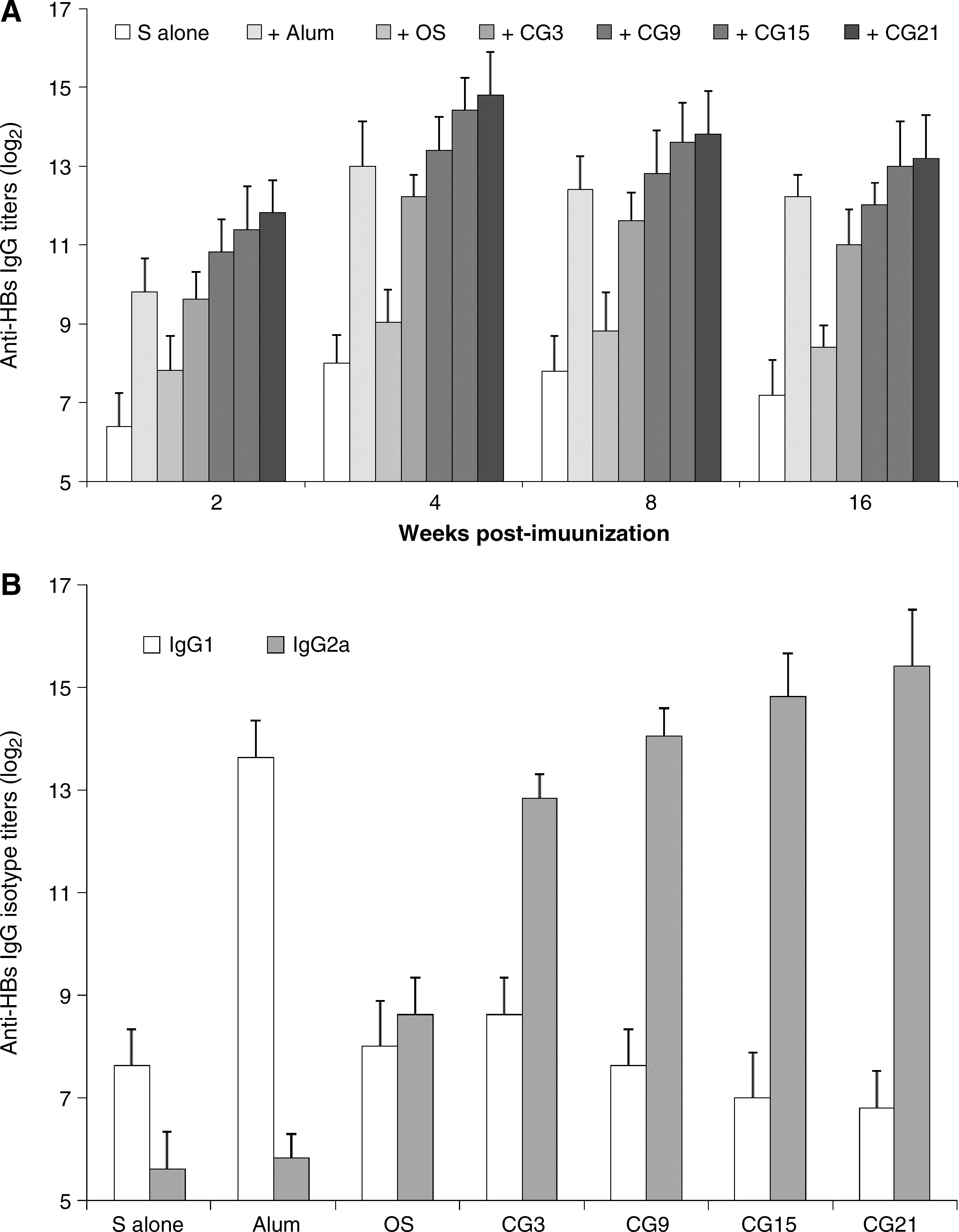
Anti-HBs total IgG, IgG2a, and IgG1, assay of mouse sera. (
IgG1 and IgG2a antibodies are markers of Th-1 and Th-2-type immune responses (34). In this context, we investigated the levels of anti-HBs IgG1 and IgG2a in sera collected at week 4, following the booster immunizations (Fig. 4B). Compared with immunization using HBsAg alone, immunizations using HBsAg in combination with alum adjuvant induced much higher levels of IgG1 (p < 0.001), but comparably low levels of IgG2a (p > 0.05) when used alone or in combination with alum. We found that plasmid pOS strongly enhanced IgG2a antibody titers (HBsAg plus pOS versus HBsAg alone or HBsAg plus alum; p < 0.001), and that plasmid pCG3 and pCG9 further enhanced the IgG2a titer (pOS versus pCG3, p < 0.001; pCG3 versus pCG9, p < 0.01). Increasing the number of CpG motifs in the plasmids further enhanced the IgG2a titers, although to a lesser extent (pCG9 versus pCG15, p > 0.05; pCG15 versus pCG21, p > 0.05; pCG9 versus pCG21, p < 0.05). The IgG1 titers from the groups of mice immunized with HBsAg alone or in combination with various plasmids were similar, but the levels were far below those observed in mice immunized with HBsAg plus alum adjuvant.
CpG plasmids promote Th-1-type cytokine production
Th-1-type immune responses are thought to play a key role in the control of HBV infection (23 –25). In the above experiments, we showed that an upregulation of anti-HBs IgG2a subclass occurred, suggesting the possibility that the CpG plasmids directed a Th-1-biased immune response in the mice. To investigate this further, we measured the cytokine production of mouse spleen cells stimulated with HBsAg in vitro (Fig. 5A). Compared with immunizations using HBsAg alone, combinations of HBsAg with alum adjuvant or with plasmid pOS enhanced production of IL-2 by more than fivefold (alum adjuvant) or eightfold (pOS) (Fig. 5A). When compared with pOS, treatment with plasmid pCG3 doubled IL-2 secretion, while treatment with pCG9 or pCG15 resulted in a further enhancement of IL-2 production. pCG21 and pCG15 promoted similar IL-2 production in the cells.
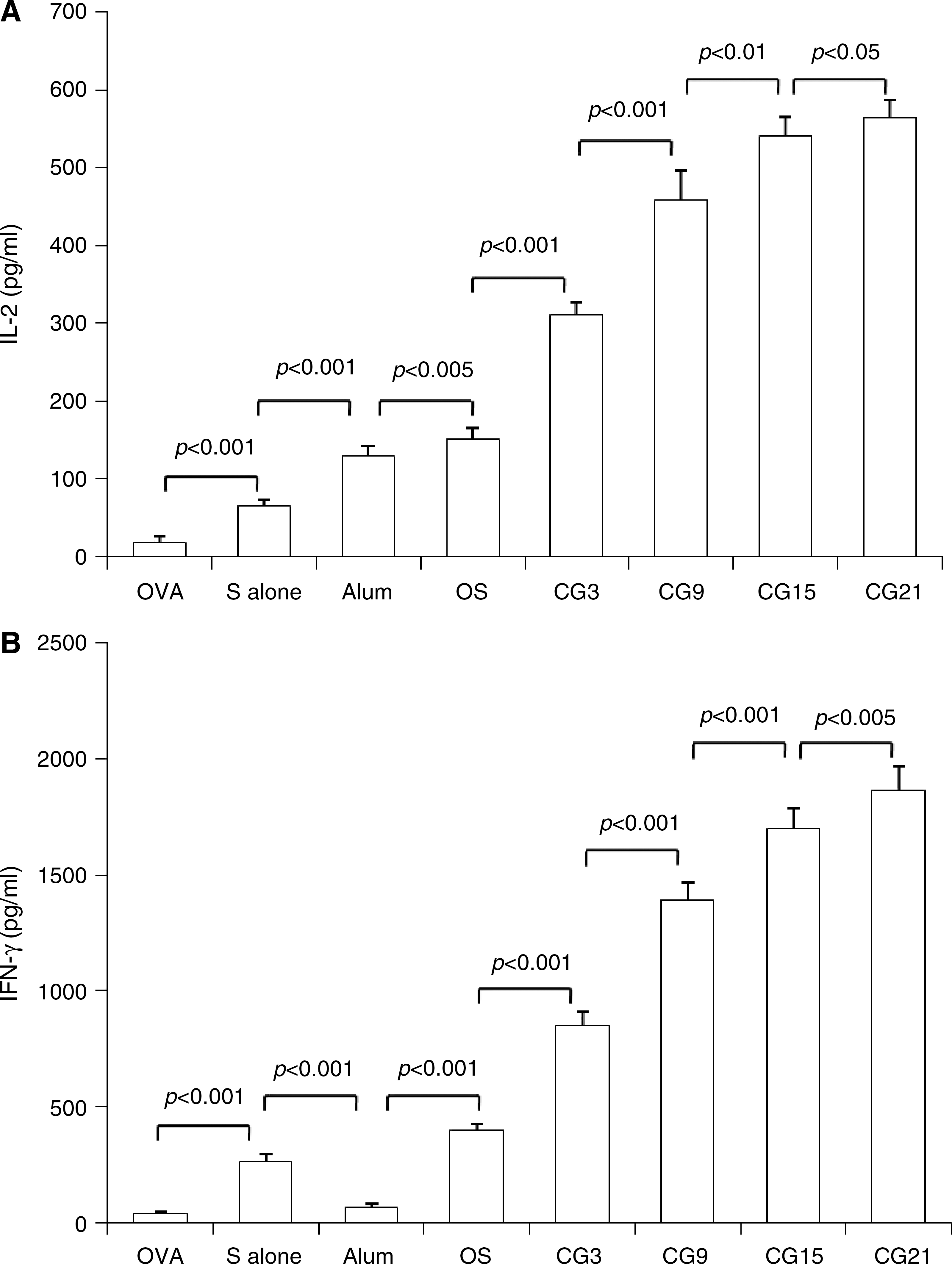
In vitro cytokine production by spleen cells. Splenocytes were isolated from different groups of immunized mice 1 wk after the booster immunization and cultured in presence of HBsAg at a concentration of 20 μg/mL. Splenocytes from the OVA-immunized mice were used as a control. Cell-free supernatants were harvested after 72 h and assayed for IL-2 (
Compared with immunization with HBsAg alone, HBsAg and alum adjuvant decreased IFN-γ production notably (p < 0.001). We found that pOS enhanced IFN-γ production by approximately 1.5-fold, while the pCG3 plasmid enhanced IFN-γ production by more than threefold (Fig. 5B). A strong correlation was found between IFN-γ production and the number of CpG motifs with plasmids pCG9, pCG15, and pCG21 (Fig. 5B). The Th-2-type cytokine IL-4 was not detectable in any of the groups tested. The spleen cells from mice immunized with HBsAg alone or plus various plasmids failed to respond to OVA (data not shown).
CpG plasmids augment HBV-specific cytotoxic killing
We measured HBsAg-specific cell lysis of spleen cells at different effector target ratios (E:T ratio; Fig. 6). Compared with the spleen cells from mice immunized with OVA plus pCG3, spleen cells from mice immunized with HBsAg alone (or with alum adjuvant) showed moderate CTL-specific activity. We found that treatment with pOS resulted in a moderate enhancement of the CTL response (E:T ratio at 100, 50, and 25, p < 0.05; Fig. 6). The presence of three copies of the CpG motif further enhanced CTL activity (pCG3 versus pOS: E:T ratio at 100, 50, and 25, p < 0.05). Substantial increases in CTL activity were observed in plasmids containing 9–15 copies of the CpG motifs (pCG9 versus pCG3: E:T ratio at 100, 50, and 25, p < 0.01; pCG15 versus pCG9: E:T ratio at 25, p < 0.05). The CTL activity of spleen cells from mice immunized with HBsAg plus plasmids containing 15 and 21 CpG copies were broadly similar.
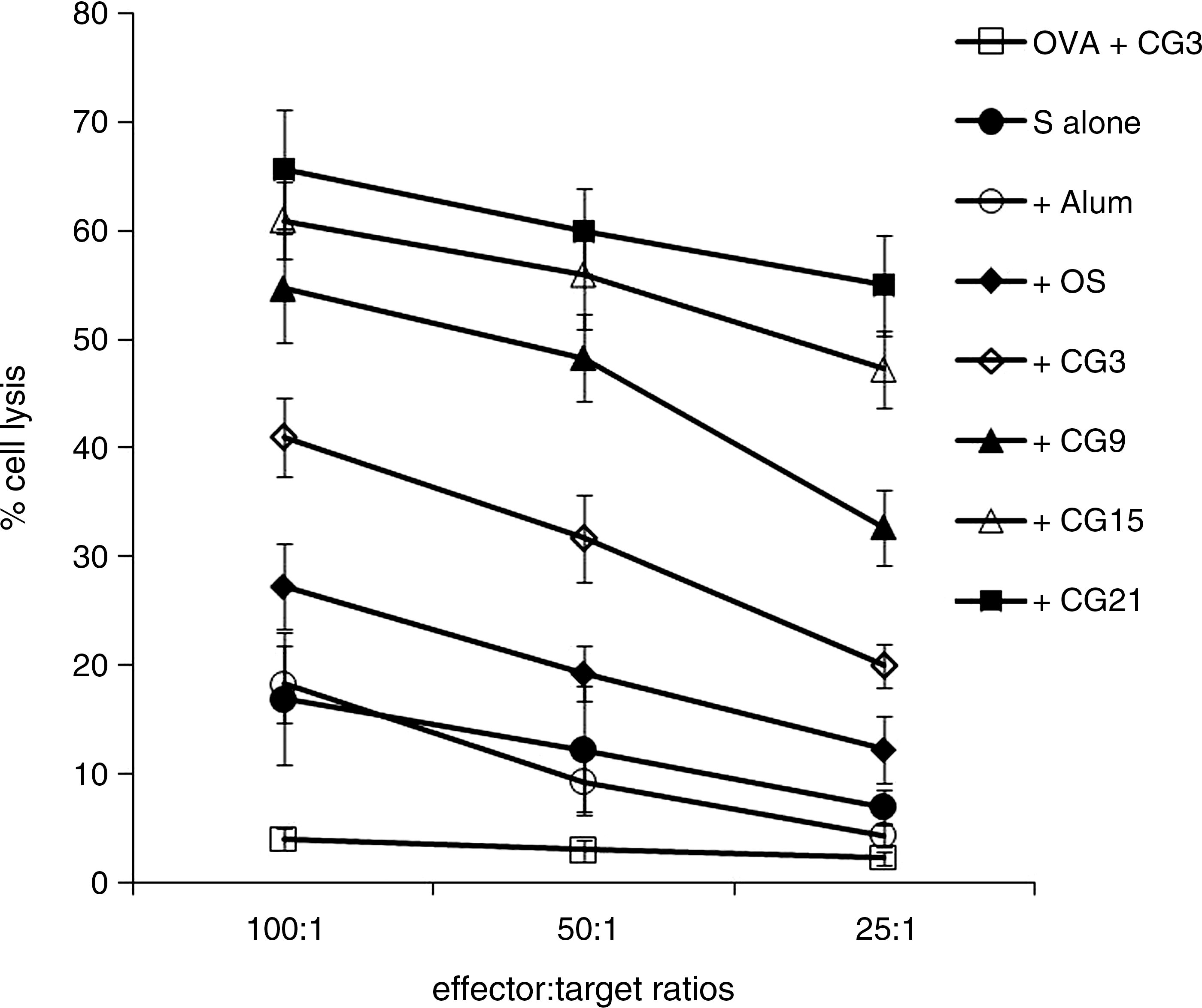
HBsAg-specific CTL response. Mice were immunized as described in the materials and methods section. Cytotoxic killing was measured by an LDH release assay 1 wk after the booster immunization. CTL activity is represented as the mean percentage of measurements performed in triplicate ± SD at different ratios of effector to target.
Discussion
Th-1-biased immunity, comprising cell-mediated immunity and strong neutralizing antibody levels, plays a critical role in controlling intracellular infections such as HBV (23 –25,35,36). In the present study, we constructed plasmids containing multiple copies of CpG motifs, and found that these plasmids strongly activated human PMBCs in vitro and efficiently enhanced the Th-1-type immune response to HBsAg in mice. These results suggest that CpG-enriched plasmids have potential application for a much-needed therapeutic hepatitis B vaccine.
CpG ODNs are recognized by TLR9 molecules expressed by plasmacytoid dendritic cells (pDCs) and B cells, and are essential for proinflammatory cytokine production and other inflammatory responses (8 –11). They also play a role in induction of the Th-1-acquired immune response and in B-cell proliferation (8 –11). Three structurally distinct classes of CpG ODNs have been identified that differentially stimulate primate immune cells (37,38). D type ODNs (also called A type ODNs) contain a central purine/pyrimidine/CpG/purine/pyrimidine motif flanked on both sides by 3–4 self-complementary bases and a 3′ poly-G tail. The CpG hexamer and flanking regions are typically composed of phosphodiester rather than phosphorothioate nucleotides. D-type ODNs trigger pDCs to produce high levels of IFN-α, but have little effect on B cells (39 –41). K-type ODNs (also called B-type ODNs) have a phosphorothioate backbone and contain multiple unmethylated CpG dinucleotides. The K-type ODN molecules stimulate B cells to produce IgM and IL-6, and trigger pDCs to produce TNF-α (40,41). The C-type ODNs combine some of the stimulatory properties of both A and B CpG-ODN classes, because they are capable of activating B cells to secrete IL-6, and pDCs to produce IFN-α (42,43). The main targets for D-CpG ODNs are mononuclear dendritic cells, which are superior to the K-type ODNs in inducing cellular immune responses (39). It has been shown that DNA vaccine vectors inserted with D-ODNs are better than K-ODNs at inducing humoral and cellular immune responses (21). Therefore, D-type CpG ODNs were used in the present study.
The plasmid vector pOS used for delivery of CpG ODNs is a “mini-plasmid,” because its length is only 812 bp. This vector contains only two elements, a pUC replication origin and a tyrosine tRNA suppressor (supF gene). The pUC origin drives plasmid replication at high copy numbers. The supF gene can be used as a selectable marker in bacteria carrying a P3 plasmid that contains amber AmpR and amber TetR genes. The supF gene allows a read-through of the amber codon, conferring ampicillin and tetracycline resistance to the transformed cells (31,32). The vector pOS is not able to confer antibiotic resistance to common bacteria, so it is safer to use in vivo compared with most of the commonly used plasmids carrying antibiotic resistance genes.
We reasoned that because treatment with plasmids pOS and pOS-woCG resulted in similar enhancing effects on proliferation and cytokine secretion from PBMCs isolated from healthy or HBV-infected individuals, pOS must contain immune-stimulatory sequences. Compared with pOS and pOS-woCG, however, pOS-CG3 significantly enhanced the proliferation and cytokine secretion responses of PBMCs. We propose that this effect is due to the presence of three copies of the CpG motifs in the plasmid. We also found that the responses of human PBMCs were further enhanced by increasing the number of CpG motifs in the plasmids, although different copies of the CpG ODNs appear to be required for maximum development of cell proliferation and cytokine secretion. We found that PBMCs obtained from healthy subjects were much more sensitive to CpG plasmids than PBMCs from HBV-infected subjects, suggesting that a defect in immune function exists in immune cells collected from HBV-infected subjects. This finding is consistent with recent reports (44,45), that the plasmids significantly augmented secretion of IFN-γ and IL-12 by human PBMCs in vitro, suggesting that the plasmids could promote Th-1 immune responses in vivo.
D-type CpG ODNs that have been shown to enhance proliferation and cytokine secretion of human PBMCs retain their immune-stimulatory activity in mice (37,38). Hence, we used mice as a model for the initial evaluation of the in vivo role of the plasmids used in this study. Compared with immunization using HBsAg alone, the vector pOS and plasmid-enriched CpG ODNs showed strong enhancement of anti-HBs total IgG and IgG2a antibodies (Fig. 4). Similar results were obtained with human PMBCs, for which treatment with pOS resulted in an obvious immune-stimulatory effect. Insertion of additional CpG ODN sequences significantly increased anti-HBs antibody titers. Compared with alum adjuvant, the CpG-enriched plasmids shifted the Th-2-biased response towards a strongly Th-1-biased response. This suggested that the increase in the humoral responses we observed was mainly dependent on the increased activation of HBsAg-specific T cells, because production of IgG2a antibody is dependent on the specific helper T cells.
Cellular immune responses play a pivotal role for viral clearance during acute or chronic HBV infection (35,36). In the present study, immunization with CpG-enriched plasmids strongly enhanced secretion of the Th-1 cytokines IL-2 and IFN-γ by spleen cells specific to HBsAg in vitro (Fig. 5). The potent effect of the CpG motifs was revealed by the strong correlation between the number of CpG motifs and the strength of the immune responses. For HBsAg, the antigen-presenting form of the endogenous antigen is not necessary for induction of CTL. HBsAg mainly exists as nanoparticles composed of more than 100 monomer molecules, which enable exogenous HBsAg to activate the CTL response through a cross-presentation mode (46 –48). CpG ODNs stimulate maturation and activation of DCs, while DCs are the key antigen-presenting cells for cross-presentation of exogenous antigens to CTL precursor cells in vivo. Moreover, the Th-1 cytokines, especially IFN-γ, act directly on CD8 T cells to stimulate the development of CTLs. Consistent with the enhanced IgG2a antibody and Th-1 cytokine levels measured in this study, the CpG plasmids also markedly enhanced HBsAg-specific CTL activity (Fig. 6).
DNA vaccines can effectively activate HBV-specific CTL in transgenic mice, and reverse immune tolerance, but in large animals (such as chimpanzees), the induced anti-HBV immune response is not ideal (49,50). The reasons for this could relate to the low transfection efficiency of DNA vaccines in large animals, which could lead to a correspondingly low level of antigen expression. However, immunization with a combination of recombinant HBsAg and DNA immune adjuvant can circumvent this problem. Synthetic CpG ODNs have been shown to be potent Th-1 adjuvants in several animal studies and in humans across a wide variety of antigens, including HBsAg (51 –53). Using plasmids to replace synthetic CpG ODNs as the adjuvant can avoid the tedious process of DNA synthesis and phosphorylation modification, and thus greatly reduce costs. In addition, supercoiled DNA is more stable than short synthetic ODNs in vivo. In recent years, technology has been developed for large-scale, high-efficiency plasmid purification, making the use of plasmids a convenient and feasible option for vaccine adjuvants. Similarly to previous reports (19,20), in the present study we also found that the adjuvant activity of plasmids is strongly correlated with the number of repeat CpG units; thus it is possible to optimize the copy number to achieve the optimal level of immune-stimulating activity. It is also possible to use a low dose of plasmids in the vaccine to promote favorable immune responses by increasing the number of CpG ODN units in the vectors.
In summary, we constructed a series of mini-plasmids, each carrying a different number of copies of D-type CpG motifs. These plasmids effectively activated PBMCs from healthy and HBV-infected individuals. They also significantly enhanced Th-1-type immune responses against recombinant HBsAg in mice. Our data provide strong evidence that plasmid-enriched CpG motifs could contribute to the development of cost-effective and efficacious therapeutic vaccines against HBV infection.
Footnotes
Acknowledgments
This work was supported by research grants from the Natural Science Foundation of China (30771929), and the Shanghai LAD Project (B901). We thank X.Q. Liu for help with manuscript preparation.
Author Disclosure Statement
No competing financial interests exist.