
Abstract
Respiratory epithelial cells are one of main targets for infections caused by influenza viruses. Recently, the induction of proinflammatory cytokines and toll-like receptors (TLRs) in normal human bronchial/tracheal epithelial cells infected with seasonal H1N1, 2009 pandemic H1N1, seasonal H3N2, or highly pathogenic H5N1 influenza virus were studied to understand the pathogenesis and early immune responses. The cells were productively infected with the viruses. Among the inflammatory cytokines tested, interleukin (IL)-8 was predominantly induced in virus-infected cells. Among the chemokines tested, interferon-γ-inducible protein-10 (IP-10) and growth-related oncogene-α (GRO-α) were predominantly induced in virus-infected cells. TLR-5 was predominantly induced in cells infected with seasonal H1N1, pandemic H1N1, or H5N1 influenza virus, and TLR-3 was predominantly induced in cells infected with seasonal H3N2 influenza virus. Taken together, the results suggest that IL-8, IP-10, and GRO-α are predominantly induced in respiratory epithelial cells infected with influenza A viruses, and that TLR-5 and TLR-3 are involved in the stimulation of virus-infected respiratory epithelial cells.
Introduction
Highly pathogenic (HP) H5N1 influenza virus first appeared in humans in 1997 when 6 out of 18 infected humans died (5). Since 2003, the infection rate of humans by HP H5N1 influenza has increased. As of October 18, 2010, 302 out of 507 infected humans with HP H5N1 influenza virus died (11). Most of the human infections resulted from direct contact with virally-infected poultry. However, the fear is that the virus will gain the capacity for direct person-to-person transmission, which could cause a pandemic comparable to or greater in mortality than the 1919 pandemic.
It has been suggested that inflammatory responses may be responsible for the pathological signs in humans infected with influenza viruses (26). Exuberant production of inflammatory cytokines and chemokines can be induced in people infected with HP H5N1 influenza virus (6,20), and in vitro in human monocyte-derived macrophages and respiratory epithelial cells (2,4).
Toll-like receptors (TLRs) are transmembrane signaling proteins that recognize proteins, lipids, carbohydrates, and nucleic acids of microbes, resulting in activation of cells and production of inflammatory cytokines (1). TLR-3, TLR-7, TLR-8, and TLR-9 recognize nucleic acids and reside in the endosomal membrane (23). TLRs that are expressed on dendritic cells, natural killer cells, macrophages, and respiratory epithelial cells are recognized by single-stranded RNA and double-stranded RNA during the replication of influenza viruses (23).
The respiratory system may be divided into the upper and lower tracts. The upper respiratory tract is composed of the nasal sinuses and pharynx, and the lower respiratory tract consists of trachea that is branched into bronchi and then into bronchioles (21). The epithelial cells in the respiratory tract are the main target cells for influenza infections.
In this study, to understand the interaction between respiratory epithelial cells and influenza viruses, we determined which cytokines and chemokines are produced and TLRs are induced in normal human primary epithelial (NHBE) cells infected with seasonal H1N1, 2009 pandemic H1N1, seasonal H3N2, or HP H5N1 influenza virus.
Materials and Methods
Viruses
Representative seasonal H1N1 (A/Brisbane/59/2007), 2009 pandemic H1N1 (A/California/04/2009), seasonal H3N2 (A/Wisconsin/15/09), and HP H5N1 (A/Vietnam/1194/04) influenza viruses were grown in 10-day-old fertilized eggs. Pandemic H1N1 and HP H5N1 influenza viruses were kindly provided by the Centers for Disease Control and Prevention (CDC, Atlanta, GA). All experiments involving HP H5N1 and pandemic H1N1 influenza viruses were performed in a BSL-3 facility approved by the Korean government.
Cell type
NHBE cells from a female or male donor were obtained from Lonza (Walkersville, MD). The cells were grown in wells of 6-well plates containing bronchial epithelial cell medium (Lonza). The medium was replenished every 3 d.
Infectivity of NHBE by influenza viruses
NHBE cell monolayers in wells of 6-well plates were infected with multiplicity of infection (MOI) of 0.01 of A/Brisbane/59/2007, A/California/04/2009, A/Wisconsin/15/09, or A/Vietnam/1194/04. The supernatants were collected at 24, 48, and 72 h post-infection (p.i.). The viral titers were determined as log 10 egg infectious dose 50/mL (log10EID50/mL).
Detection of inflammatory cytokines and chemokines
NHBE cell monolayers in wells of 6-well plates were infected with MOI of 0.01 of A/Brisbane/59/2007, A/California/04/2009, A/Wisconsin/15/09, or A/Vietnam/1194/04. The supernatants were collected at 24, 48, and 72 h p.i. The collected supernatants were first screened for human inflammatory cytokines or chemokines using a multi-analyte ELISArray kit (SABioscience, Frederick, MD). Individual ELISA kits for human interleukin-8 (IL-8), interferon-γ-inducible protein-10 (IP-10), or growth-related oncogene-α (GRO-α) were obtained from SABioscience. The assays were performed as described by the manufacturer. Briefly, 50 μL of sample dilution buffer was added to each well of an ELISA plate, followed by 50 μL of the particular supernatant. The plate was gently shaken and incubated for 30 min at room temperature. The wells were washed with wash buffer and 100 μL of a dilution of the particular detection antibody was added to each well. After incubation for 1 h at room temperature, each well was washed and 100 μL of horseradish peroxidase-conjugated avidin was added to each well. Following incubation for 20 min at room temperature, each well was washed and 100 μL of development solution was dispensed. After incubating for 15 min, 100 μL of stop solution was added to each well. The absorbance of the fluid in each well was read at 450 nm using an ELISA plate reader (Tecan, Männedorf, Switzerland). The amount of the individual cytokine and chemokine was determined based on the standard curve of each cytokine or chemokine.
Quantification of human toll-like receptors by real-time polymerase chain reaction
Total RNA was collected from NHBE cells infected with A/Brisbane/59/2007, A/Wisconsin/15/09, A/California/04/2009, or A/Vietnam/1194/04 at a MOI of 2 using TRIzol (Invitrogen, Carlsbad, CA). One milliliter of TRIzol (Invitrogen) was added to the cells in wells of a 6-well plate and incubated at room temperature for 5 min. Chloroform (200 μL) was added, the solution was mixed by vortexing for 15 sec and centrifuged for 15 min at 12,000 rpm and 4°C. The upper RNA-containing band was collected and mixed with 500 μL of isopropanol (Sigma-Aldrich, St. Louis, MO) in a new 1.5 mL tube. Each sample was centrifuged for 10 min at 10,000 rpm and 4°C, and the RNA-containing pellet was washed with 100 μL of 75% ethanol in water by centrifugation for 5 min at 10,000 rpm and 4°C. The washed RNA was resuspended in 50 μL of diethyl pyrocarbonate-treated water. The mRNA of TLRs was quantified using quantitative real-time polymerase chain reaction (PCR). To synthesize the cDNA, 1 μL of oligo dT primers (0.5 μmol) (Promega, Madison, WI) was added to a total volume of 9 μL in a 0.05-mL tube. The mixture was reacted for 5 min at 70°C prior to incubation for 5 min at 4°C. Then, each sample received 4 μL of 25 mM MgCl2, 4 μL of 5 × reverse transcriptase enzyme buffer, 1 μL of RNase inhibitor, 1 μL of reverse transcriptase, and 1 μL of dNTP (10 mM). Each sample was incubated for 5 min at 25°C for 60 min at 42°C and 15 min at 70°C. SYBR-green-based real-time PCR was performed using a Roto-Gene 6000 apparatus (Corbett, Mortlake, Australia), and SensiMix Plus SYBR (Quantace, London, U.K.), based on both manufacturers' recommendations. A duplicate of each sample was run. A total volume of 20 μL containing 2 μL cDNA, 10 μL SYBR mixture and human TLR-specific primers (Table 1) [1 μL forward primer (20 pmol) and 1 μL of reverse primer (20 pmol)] was used for 40 cycles of PCR: 5 sec at 95°C, 15 sec at 60°C, and 25 sec at 72°C. Cytokine expression levels in lung tissues were normalized to those of human glyceraldehyde-3-phosphate dehydrogenase (GAPDH). The results of real-time PCR were quantified by the comparative threshold method (16).
Statistical analysis
Statistical analysis was performed using the SPSS software package, version 10.0 (SPSS, Chicago, IL). The Student's t-test was performed to compare data of seasonal H1N1 with those of the other viruses. A p-value < 0.05 was considered statistically significant.
Results
Infectivity of human primary respiratory epithelial cells by influenza A viruses
We determined whether NHBE cells could be productively infected with A/Brisbane/59/2007 (seasonal H1N1), A/Wisconsin/15/09 (seasonal H3N2), A/California/04/2009 (2009 pandemic H1N1), or A/Vietnam/1194/04 (H5N1). NHBE cells were infected at MOI of 0.01 and the viral titers in the supernatants were determined as log10EID50/mL. NHBE cells were productively infected with these viruses (Fig. 1). The viral titers in NHBE cells infected with A/Brisbane/59/2007 (seasonal H1N1), A/California/04/2009 (2009 pandemic H1N1), A/Wisconsin/15/09 (seasonal H3N2), and Vietnam/1194/04 (H5N1) at 72 h after infection were 5.0, 5.0, 5.0, and 5.5 log10EID50/mL, respectively.

Infectivity of NHBE cells by influenza viruses. NHBE cells grown in wells of 6-well plates were infected with A/Brisbane/59/2007 (seasonal H1N1), A/California/04/2009 (2009 pandemic H1N1), A/Wisconsin/15/09 (seasonal H3N2), or A/Vietnam/1194/04 (HP H5N1) at a MOI of 0.01. The viral titers in supernatants collected at 24, 48, and 72 h p.i. were determined as log10EID50/mL. The data are representative of five independent experiments (mean ± standard error). Color images available online at
Induction of inflammatory cytokines and chemokines in human primary respiratory epithelial cells infected with influenza A viruses
To deduce the inflammatory cytokines and chemokines that were predominantly induced in human primary respiratory epithelial cells, NHBE cells were infected with A/Brisbane/59/2007 (seasonal H1N1), A/California/04/2009 (2009 pandemic H1N1), A/Wisconsin/15/09 (seasonal H3N2), or A/Vietnam/1194/04 (H5N1) at MOI of 0.01. ELISA using the supernatants of NHBE-infected cells revealed that the inflammatory cytokine IL-8 was predominantly detected in infection with influenza A viruses (Fig. 2A). The induced amounts of IL-8 in the supernatants of NHBE cells infected with seasonal H1N1, 2009 pandemic H1N1, seasonal H3N2, or HP H5N1 influenza virus at 72 h p.i. were 480.6, 1259.97, 464.8, and 2735.6 pg/mL, respectively (Fig. 2B). Among the assayed chemokines, IP-10 and GRO-α were predominantly detected in infection with 2009 pandemic H1N1, seasonal H3N2, and HP H5N1 influenza virus (Fig. 3A). The amount of IP-10 in the supernatant of NHBE cells infected with seasonal H1N1, 2009 pandemic H1N1, seasonal H3N2, or HP H5N1 influenza virus at 72 h p.i. was 3700.6, 6610.4, 8518.8, and 17977.2 pg/mL, respectively (Fig. 3B). When the amount of GRO-α in the supernatants of NHBE cells infected with influenza A viruses was determined, the amounts of GRO-α induced in the supernatant of NHBE cells infected with seasonal H1N1, 2009 pandemic H1N1, seasonal H3N2, or HP H5N1 influenza virus at 72 h p.i. were 960.5, 3647.8, 1580.3, and 2218.49 pg/mL, respectively (Fig. 3C).
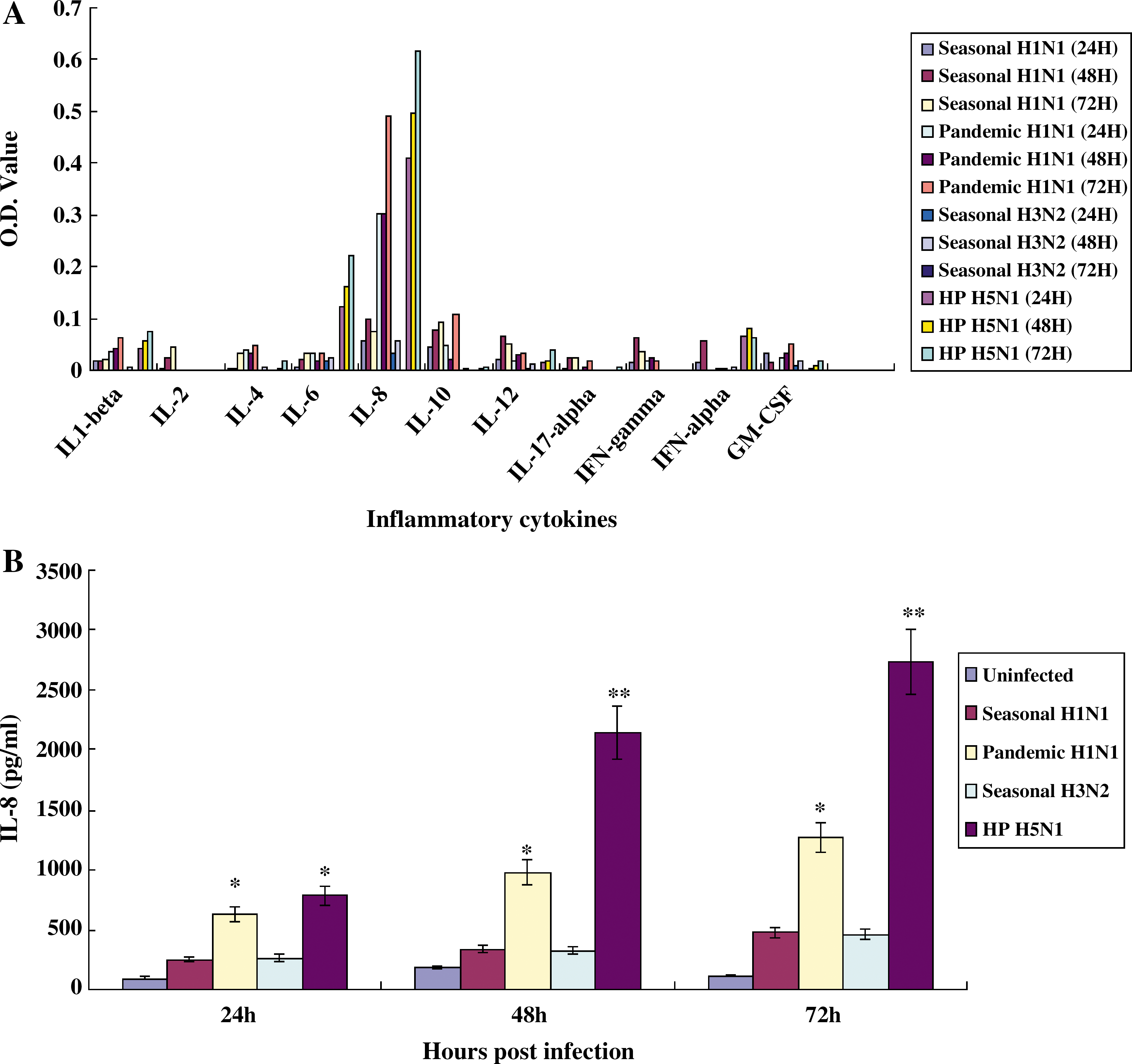
Detection of proinflammatory cytokines in NHBE cells infected with influenza A viruses. NHBE cells grown in wells of 6-well plates were infected with A/Brisbane/59/2007 (seasonal H1N1), A/California/04/2009 (2009 pandemic H1N1), A/Wisconsin/15/09 (seasonal H3N2), or A/Vietnam/1194/04 (HP H5N1) (MOI 0.01), and supernatants were collected at 24, 48, and 72 h p.i. The secretion of proinflammatory cytokines was first screened with a human inflammatory-cytokine multi-analyte ELISArray kit (
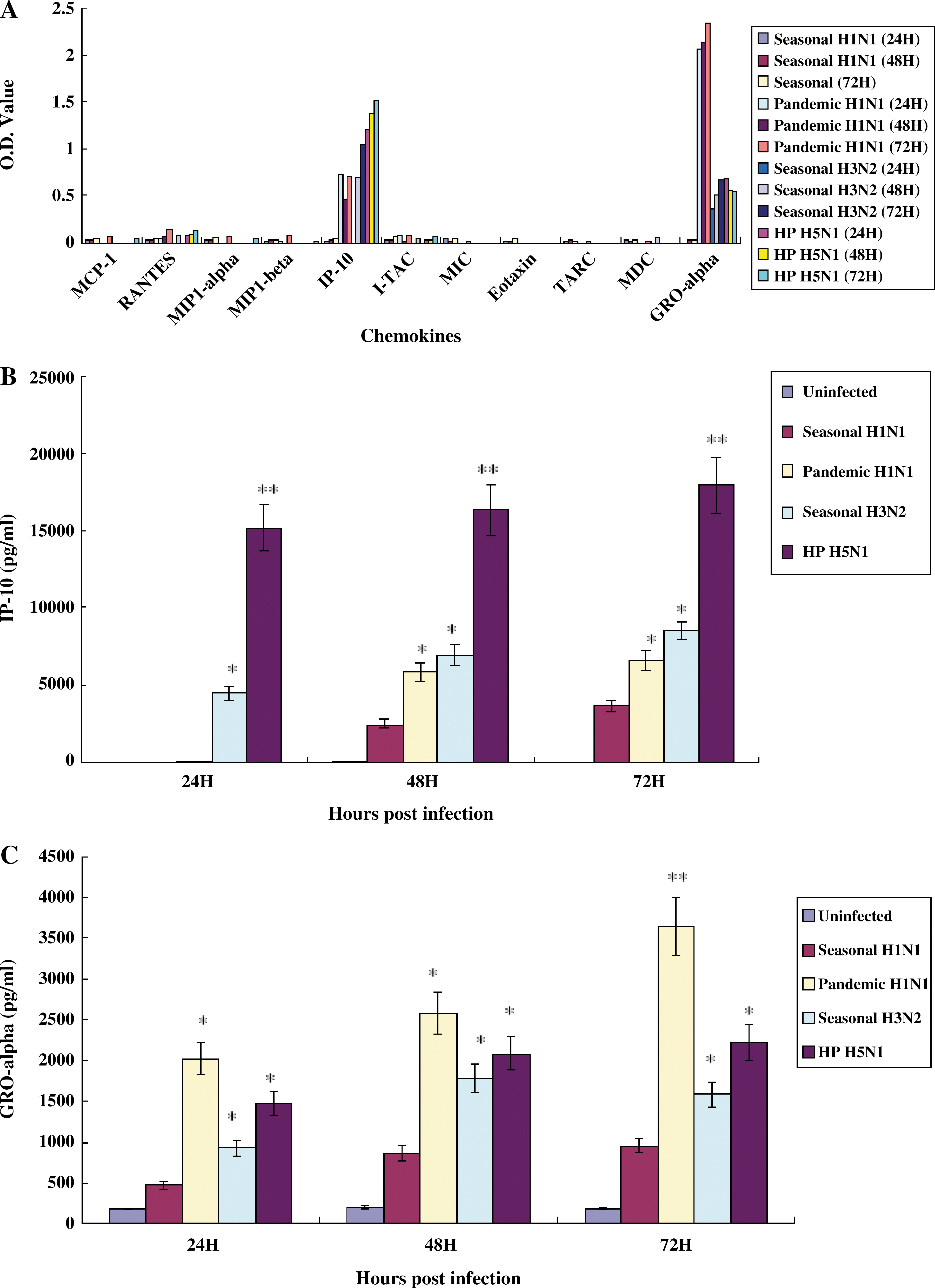
Detection of chemokines in NHBE cells infected with influenza A viruses. NHBE cells grown in wells of 6-well plates were infected with A/Brisbane/59/2007 (seasonal H1N1), A/California/04/2009 (2009 pandemic H1N1), A/Wisconsin/15/09 (seasonal H3N2), or A/Vietnam/1194/04 (HP H5N1) (MOI 0.01), and supernatants were collected at 24, 48, and 72 h p.i. The secretion of chemokines was first screened with a human chemokine multi-analyte ELISArray kit (
Induction of TLRs in human primary respiratory epithelial cells infected with influenza A viruses
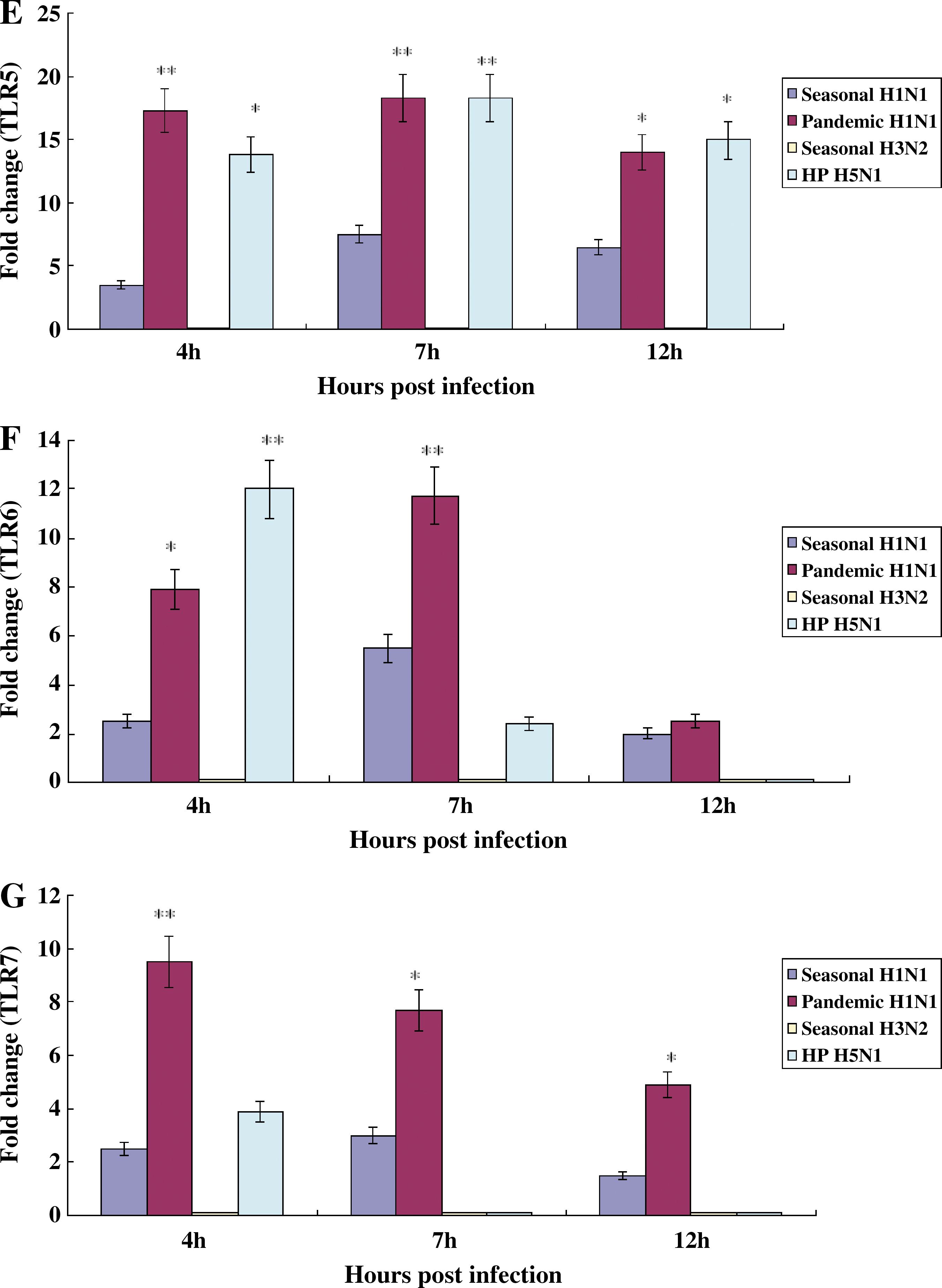
We quantified the mRNA of TLRs obtained from NHBE cells infected with influenza A viruses at 4, 7, or 12 h p.i. The TLR4, TLR5, TLR6, TLR7, and TLR8 were weakly induced in NHBE cells infected with seasonal H1N1 influenza virus (Fig. 4D, E, F, G, and H), the TLR4, TLR5, TLR6, TLR7, and TLR8 were actively induced in NHBE cells infected with pandemic H1N1 influenza virus (Fig. 4D, E, F, G, & H), TLR2 and TLR3 were dominantly induced in NHBE cells infected with seasonal H3N2 influenza virus (Fig. 4B and C), and TLR1, TLR4, TLR5, TLR6, and TLR9 were strongly induced in NHBE cells infected with HP H5N1 influenza virus (Fig. 4A, D, E, F, and I).
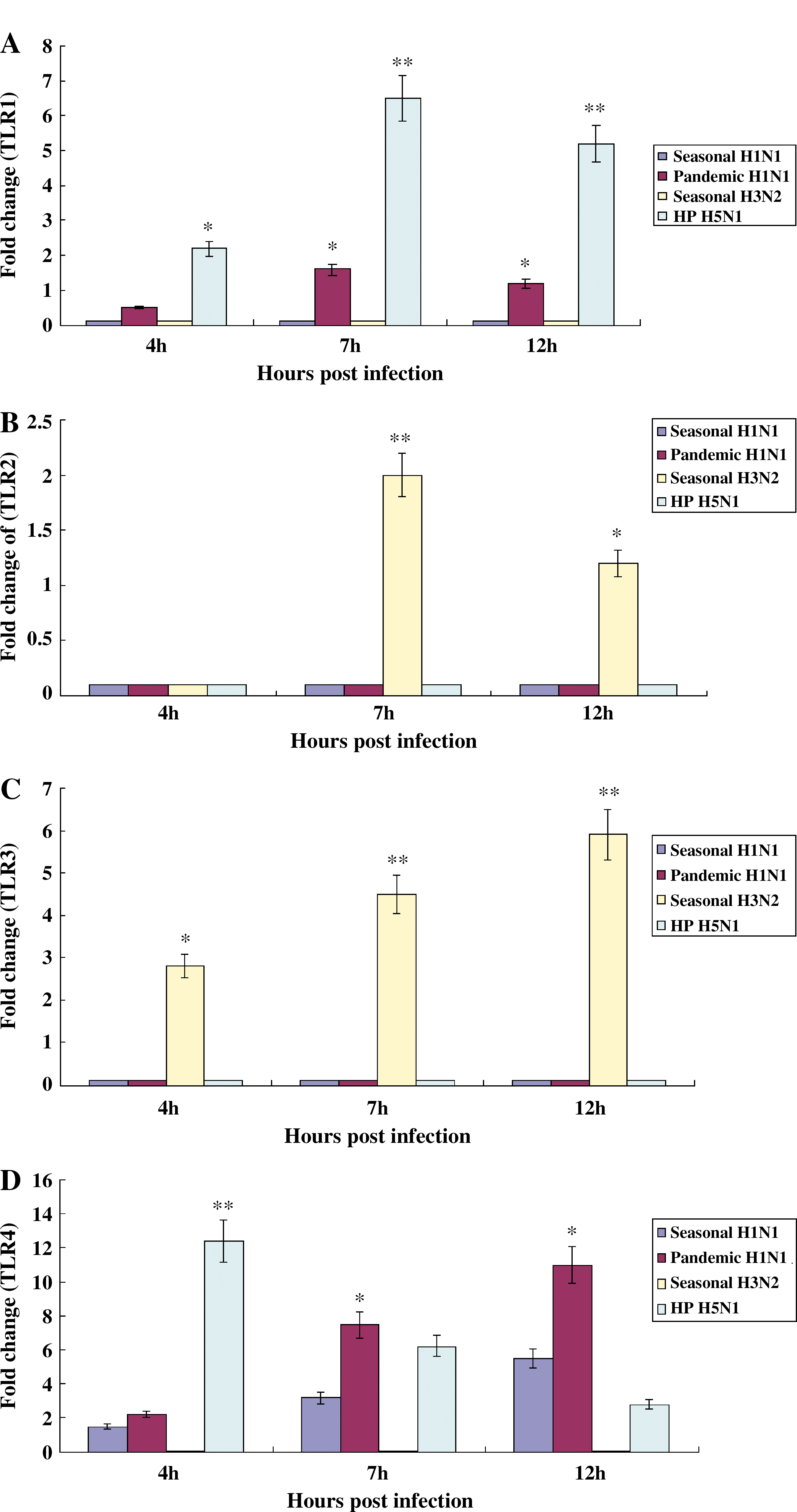
Induction of TLRs in NHBE cells infected with influenza A viruses. Total RNA was collected on 4, 7, or 12 h after infection of NHBE cells with A/Brisbane/59/2007 (seasonal H1N1), A/California/04/2009 (2009 pandemic H1N1), A/Wisconsin/15/09 (seasonal H3N2), or A/Vietnam/1194/04 (HP H5N1) (MOI 2), and mRNA of TLRs was quantified using SYBR-based quantitative real-time PCR and human TLR-specific primers. TLR expression levels in cells were normalized to those of human GAPDH. The statistical analysis was performed compared to data of seasonal H1N1 influenza virus. We did not show the result of TLR10 since its induction was no different from that of uninfected NHBE cells (*p < 0.05; **p < 0.001;

Discussion
Human respiratory epithelial cells are one of the main targets for infection by influenza viruses. Studying the interaction between respiratory epithelial cells and influenza viruses, such as seasonal, pandemic, or HP H5N1 influenza viruses, will help to understand the early immune responses and viral pathogenesis. The present study found that normal respiratory epithelial cells could be productively infected with seasonal H1N1, 2009 pandemic H1N1, seasonal H3N2, and HP H5N1 influenza viruses, resulting in the predominant production of the proinflammatory cytokine IL-8 and the chemokines IP-10 and GRO-α, and the predominant induction of TLR-5.
When NHBE cells were infected with seasonal H1N1, 2009 pandemic H1N1, seasonal H3N2, or HP H5N1 influenza virus, productive infections resulted, with similar titers in the absence of trypsin. This result suggests that NHBE cells may mimic normal respiratory cells in humans. Previous studies also showed that human bronchial epithelial cells can be productively infected with seasonal H1N1 or HP H5N1 influenza viruses (2,3,29).
Presently, the proinflammatory cytokine IL-8 was predominantly produced in NHBE cells infected with seasonal H1N1, 2009 pandemic H1N1, seasonal H3N2, or HP H5N1 influenza viruses, with infection by the 2009 pandemic H1N1 virus most potently inducing IL-8 in NHBE cells, as compared with seasonal H1N1 and seasonal H3N2. This result suggests that IL-8 may be one of the main proinflammatory cytokines causing the pathological damage and immune responses to human bronchial and tracheal epithelial cells. An earlier study of cytokine profiles of pulmonary secretions from children infected with the 2009 pandemic H1N1 influenza virus also showed that IL-8 was predominantly produced in the lungs (15).
Presently, chemokine IP-10 was strongly induced in NHBE cells infected with 2009 pandemic H1N1, seasonal H3N2, and most potently, by HP H5N1 influenza virus. A previous study reported that compared to human H1N1 influenza viruses, HP H5N1/97 and H5N1/04 influenza viruses were more potent inducers of IP-10, interferon-β, RANTES, and IL-6, in human alveolar and bronchial epithelial cells (2). Another study with human alveolar epithelial cells and lung microvascular endothelial cells also showed that H5N1 influenza virus induced higher amounts of IP-10 in human alveolar epithelial cells than did seasonal H1N1 influenza virus (3).
Interestingly, the GRO-α chemokine was induced more potently in NHBE cells infected with 2009 pandemic H1N1 influenza virus than in cells infected with seasonal H1N1, seasonal H3N2, and HP H5N1 influenza viruses. GRO-α may thus be exploited as a chemokine marker for human infections caused by the 2009 pandemic H1N1 influenza virus. An earlier study on the systemic cytokine and chemokine responses in children infected with 2009 pandemic H1N1 influenza virus reported that patients suffering from pneumonia had the highest elevated serum levels of interferon-γ and IL-5 than patients without pneumonia (24). Among the patients with pneumonia, the serum level of the chemokine monocyte chemoattractant protein-1 was higher in patients with severe respiratory failure than patients with mild pneumonia (24). However, these studies did not measure GRO-α levels in serum of patients infected with 2009 pandemic H1N1 influenza virus.
Of the TLRs, TLR-5 was most potently induced in NHBE cells infected with seasonal H1N1, 2009 pandemic H1N1, or HP H5N1 influenza viruses, and TLR-3 was predominantly induced in NHBE cells infected with seasonal H3N2 virus. This suggests that TLR-5 may be a main TLR involved in recognizing these viruses in bronchial epithelial cells in humans. Until now, little has been known about the induction of TLRs in cells infected with influenza viruses. One study using human alveolar and bronchial epithelial cell lines showed that the infection of these cells with human H3N2 influenza virus (A/Scotland/20/74) increased the induction of TLR-3 (10). In another study, stimulation of TLR-5 with ligand increases the vaccine efficiency of influenza matrix protein M2 in mice (12). The vaccination of mice with a fusion protein of TLR-5 ligand, flagellin, and M2 protein protected mice from a lethal challenge with influenza A virus (12).
In conclusion, our data suggest that influenza A viruses may induce TLR-5, resulting in proinflammatory cytokines and chemokines in normal human respiratory epithelial cells.
Footnotes
Acknowledgments
This study was supported in part by a grant (A084411) from the Korea Healthcare Technology R&D Project, Ministry for Health, Welfare and Family Affairs, Republic of Korea. A staff in HARRISCO, an English editing company, edited this manuscript.
Author Disclosure Statement
No competing financial interests exist.