
Abstract
Both T-helper 17 cells (Th-17) and CD4+CD25+ regulatory T cells (Tregs) play important roles in the pathogenesis of hepatitis B virus (HBV) infection. Recent studies have suggested that Th-17 and Treg cells are increased in patients with chronic hepatitis B (CHB). This study further characterizes Th-17 and Treg cells in the blood of HBV-associated acute-on-chronic liver failure (ACLF) patients, and aids our understanding of how the two subsets of CD4+ T cells affect each other and contribute to survival. Blood samples were obtained from 30 patients with HBV-associated ACLF, 30 patients with CHB, and 30 normal controls (NC). The frequencies of Th-17 and Treg cells were determined by intracellular cytokine staining analysis. To observe the suppressive function of Tregs, purified CD4+CD25+ Tregs from peripheral blood mononuclear cells (PBMCs) were co-cultured with CD4+CD25− T cells for 48 h, and then IFN-γ and IL-17A from the supernatants were measured by ELISA. We found that both Th-17 and FoxP3+ Treg cells were increased in ACLF patients. IL-17A secretion by CD4+ T cells was not regulated by Treg cells, even though Tregs exhibited significant inhibition of IFN-γ production. Most importantly, the ratio of Th-17 to Treg cells was associated inversely with the survival of ACLF patients. These findings provide new information regarding the pathogenesis of HBV-associated ACLF, and the ratio of Th-17 to Tregs may represent a potential prognostic marker for the disease.
Introduction
The pathophysiological basis of ACLF is uncertain but current hypotheses suggest that systemic inflammatory responses may underlie the transition of a patient from a stable state to developing progressive liver injury and end-organ failure (31). Recently, it was shown that a novel and unique subset of CD4+ T cells, T-helper 17 cells (Th-17), may play an important role in the pathogenesis of HBV infection (9,20,38,45). Th-17 cells are distinct from Th-1 and Th-2 cells in that they preferentially produce interleukins (IL)-17A and F, and require RORγt as a key transcription factor for their differentiation (10,13,28). Although some reports have shown roles for Th-17 cells in host defense against microbes and fungi (11,36,37), evidence accumulating from human and mouse studies suggests that Th-17 cells are the principal mediators during the pathogenesis of autoimmune and inflammatory disorders (18,26,36). Ge et al. reported that circulating Th-17 frequency was associated with disease progression in HBV-infected patients (9), and other studies demonstrated that Th-17 frequency is significantly increased in HBV-associated ACLF or severe CHB patients (9,45).
Conversely, another subset of CD4+ T cells, CD4+CD25+ regulatory T cells (Tregs), which are characterized by their constitutive expression of CD25 and FoxP3 and by immunological suppression (2,5,15,23), has been proposed to be able to restrain the immune response, thus contributing to limit liver damage (39,43). In chronic HBV infection, an imbalance between Tregs and effector T cells has recently been described by several groups, and suggests that either the number or functional imbalance of Tregs in the blood or liver may be a reason for persistent HBV infection and disease progression (7,33,39).
Although increased Th-17 and Treg frequencies in patients with chronic hepatitis B has been reported, little is known about the relationship between circulating Th-17 and Tregs, or about their frequencies in patients with HBV-associated ACLF. In addition, whether or not Tregs can regulate Th-17 in HBV-infected patients remains unknown. More importantly, the association between the survival of ACLF patients and Tregs or Th-17 is largely unclear. To address these issues, Th-17 and FoxP3+ Tregs in the peripheral circulation were evaluated in HBV-associated ACLF patients. The suppressive activity of Tregs on IL-17 production was also observed. Furthermore, the association between the survival of ACLF patients and Tregs or Th-17 cells was analyzed.
Materials and Methods
Clinical samples
The study protocol was approved by the ethics committee of our unit, and written informed consent was obtained from each subject before blood sampling. Blood samples were obtained from 30 patients with HBV-associated ACLF, 30 patients with CHB, and 30 normal controls (NC). The blood samples of ACLF patients were collected at 1 d to 1 wk after ACLF diagnosis. All patients and healthy donors were well matched for age and gender. The standards for the diagnoses of CHB and ACLF have been described in detail previously (31,32,39,45,47 –49). Patients were excluded if they had any causes of liver disease other than HBV infection, such as HCV, HEV, HGV, HIV, hepatocellular carcinoma, autoimmune disease, or alcoholic hepatitis. No patients received any anti-HBV agent or steroid during the 6 mo before sampling. The basic characteristics of enrolled subjects are listed in Table 1.
Data are shown as median and range.
ND, not determined; NC, normal controls; CHB, chronic hepatitis B; ACLF, acute-on-chronic liver failure; ALT, alanine aminotransferase, TBIL, total bilirubin; PTA, prothrombin activity; HBsAg, hepatitis B surface antigen; HBsAb, hepatitis B surface antibody; HBeAg, hepatitis B early antigen; HBeAb, hepatitis B early antibody; HBcAb, hepatitis B core antibody.
Flow cytometric analysis
PE-conjugated anti-CD25, PerCP-conjugated anti-CD3, and allophycocyanin (APC)-conjugated anti-CD4 and anti-CD8 were purchased from BD PharMingen (San Diego, CA). FITC-conjugated anti-FoxP3 and PE-conjugated anti-IL-17A mAbs were purchased from eBiosciences (San Diego, CA). For intracellular staining with FoxP3, cells were first stained with PerCP-conjugated anti-CD3, APC-conjugated anti-CD4, and PE-conjugated anti-CD25 Abs, then permeabilized and fixed using eBioscience fix/perm according to the manufacturer's instructions. FITC-anti-FoxP3 was added after permeabilization for 30 min. For Th-17 detection, whole blood samples (200 μL) were activated with phorbol-12-myristate 13-acetate (PMA; 50 ng/mL) and ionomycin (1 μg/mL) for 5 h. After surface staining for CD3+CD8+ T cells and lysis of red blood cells, the remaining cells were permeabilized and stained with and PE-conjugated anti-human IL-17A. The cells were fixed in 1% paraformaldehyde and 3- or 4-color flow cytometric analyses were performed using a FACSCalibur and CellQuest software (Becton Dickinson, San Jose, CA).
Cell isolation and stimulation
Peripheral blood mononuclear cells (PBMCs) were isolated by Ficoll-Hypaque density gradient centrifugation from heparinized blood. CD4+CD25+ Tregs were isolated from PBMCs by CD4 negative selection followed by CD25 positive selection, using a CD4+CD25+ T-cell isolation kit (Miltenyi Biotech, Auburn, CA) according to the manufacturer's instructions. The purity of CD4+CD25+FoxP3+ Tregs was ≥90% (39). For interferon-γ (IFN-γ) and IL-17A detection, CD4+CD25− T cells (1 × 105/well) in a U-bottom 96-well plate in triplicate were stimulated with T-cell expander (CD3/CD28 Dynabeads; Dynal Biotech, Lafayette Hill, PA) in RPMI 1640 medium containing 10% FCS and recombinant IL-2 (20 U/mL). The cells were cultured alone or with Tregs at a ratio of 1:1 using the same conditions for 48 h before supernatants were collected for IFN-γ and IL-17A measurement with commercial ELISA kits (Biosource International Inc., Camarillo, CA) according to the manufacturer's instructions (46).
Statistical analysis
Results are expressed as means ± SD for percentages. Statistical comparisons between two groups used a Mann-Whitney non-parametric U test. The Kruskal-Wallis H non-parametric test was used for multiple comparisons with a Bonferroni step-down (Holm) correction (39,46). Spearman correlation analysis was used to evaluate relationships between two variables. Data analysis was done using SPSS version 13.0 for Windows (SPSS Inc., Chicago, IL). Significance was set at p < 0.05.
Results
Th-17 and Treg frequencies are both increased in patients with ACLF
We first compared the Th-17 proportions in normal controls and HBV-infected patients. Th-17 was defined as the population of CD3+CD8−IL-17A+ T cells and the percentage of total CD4+ T cells. There was a significantly higher frequency of circulating Th-17 in ACLF patients compared to normal controls (6.47 ± 1.87% versus 1.98 ± 0.62%; p < 0.001) and CHB patients (3.62 ± 1.09%; p < 0.001; Fig. 1A and B). CHB patients had a higher frequency of Th-17 than normal controls (p < 0.001; Fig. 1A and B).
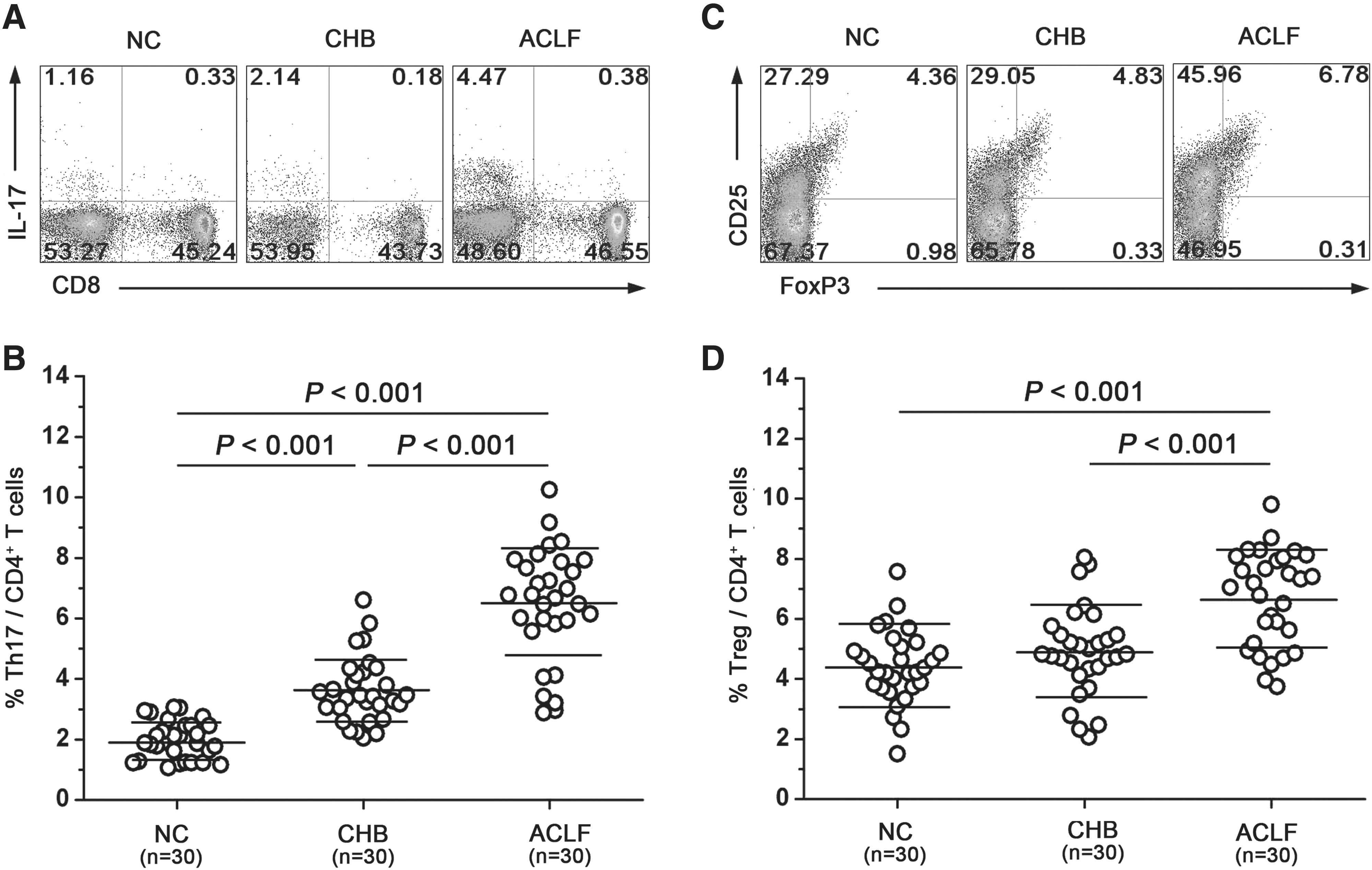
Th-17 and Treg frequencies are both increased in patients with ACLF. (
We next compared the Treg frequencies with Tregs defined as the population of CD3+CD4+CD25+FoxP3+ T cells taken as a percentage of total CD4+ T cells, in normal controls and patients. In contrast to the Th-17 frequency, there was no significant difference in Treg frequency in CHB patients than in normal controls (4.91 ± 1.47% versus 4.40 ± 1.23%; p > 0.05; Fig. 1C and D). But a significant increase in Treg frequency was observed among ACLF patients (6.69 ± 1.58% ACLF versus NC, p < 0.001; ACLF versus CHB, p < 0.001; Fig. 1C and D).
Preferentially increased Th-17 are positively correlated with Tregs in ACLF patients
Th-17 and Tregs were both significantly increased in ACLF patients. Thus, it is likely that there was a relationship between Tregs and Th-17 in ACLF patients. Strikingly, there were positive correlations between Th-17 and Tregs (r = 0.723, p < 0.001; Fig. 2A). In CHB patients and normal controls, there was no correlation between Th-17 and Tregs (data not shown).
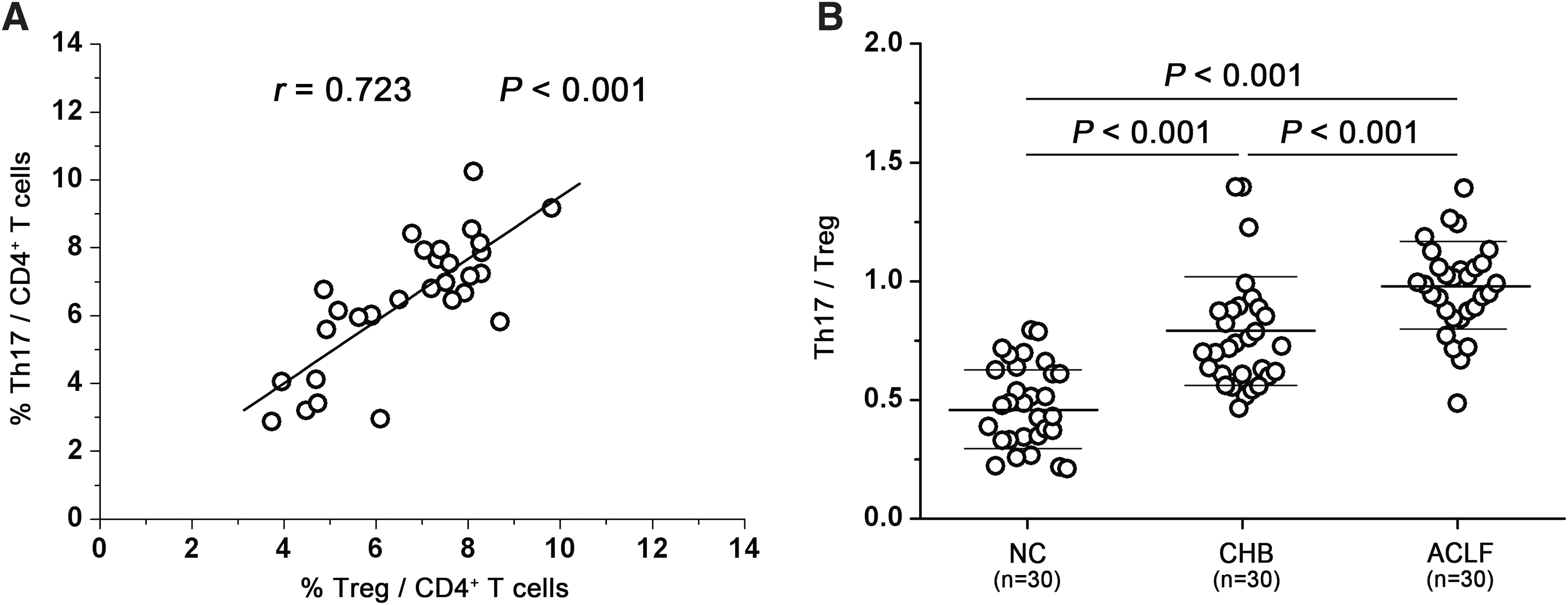
Preferentially increased Th-17 cells are positively correlated with Tregs in ACLF patients. (
To further explore if either Th-17 or Tregs were preferentially increased in ACLF patients, we analyzed the ratios of Th-17 to Treg frequency in normal controls, CHB patients, and ACLF patients. We found that the ratio in ACLF patients was significantly increased compared with normal controls and CHB patients (NC 0.48 ± 0.17, CHB 0.77 ± 0.24, ACLF 0.98 ± 0.19; p < 0.001; Fig. 2B). CHB patients had a higher ratio than normal controls (p < 0.001). We also analyzed the mean value of Treg and Th-17 frequency, and found that compared with normal controls, the frequency of Tregs was increased 1.12-fold in CHB patients, and increased 1.52-fold in ACLF patients, Th-17 frequency was increased 1.83-fold in CHB patients, and was increased 3.27-fold in ACLF patients. These data demonstrate that Th-17 cells were preferentially increased in ACLF patients, but Tregs were not.
IL-17A secretion by CD4+ T cells is not regulated by Tregs
To address whether IL-17A secretion by CD4+ T cells was regulated by Tregs, we stimulated CD4+CD25- T cells with or without Tregs using CD3/CD28 Dynabead stimulation for 48 h. Then, IL-17A and IFN-γ production was detected by ELISA. We found that Tregs significantly inhibited IFN-γ production, but not IL-17A production (Fig. 3A and 3B). There was no significant difference in the proliferation between CD4+CD25- T cells and those co-cultured with Tregs after 48 h of stimulation as assessed by CFSE staining and (3H)-thymidine incorporation assays (data not shown). These data demonstrate that Tregs have a suppressive role on IFN-γ production, but cannot suppress IL-17A production.
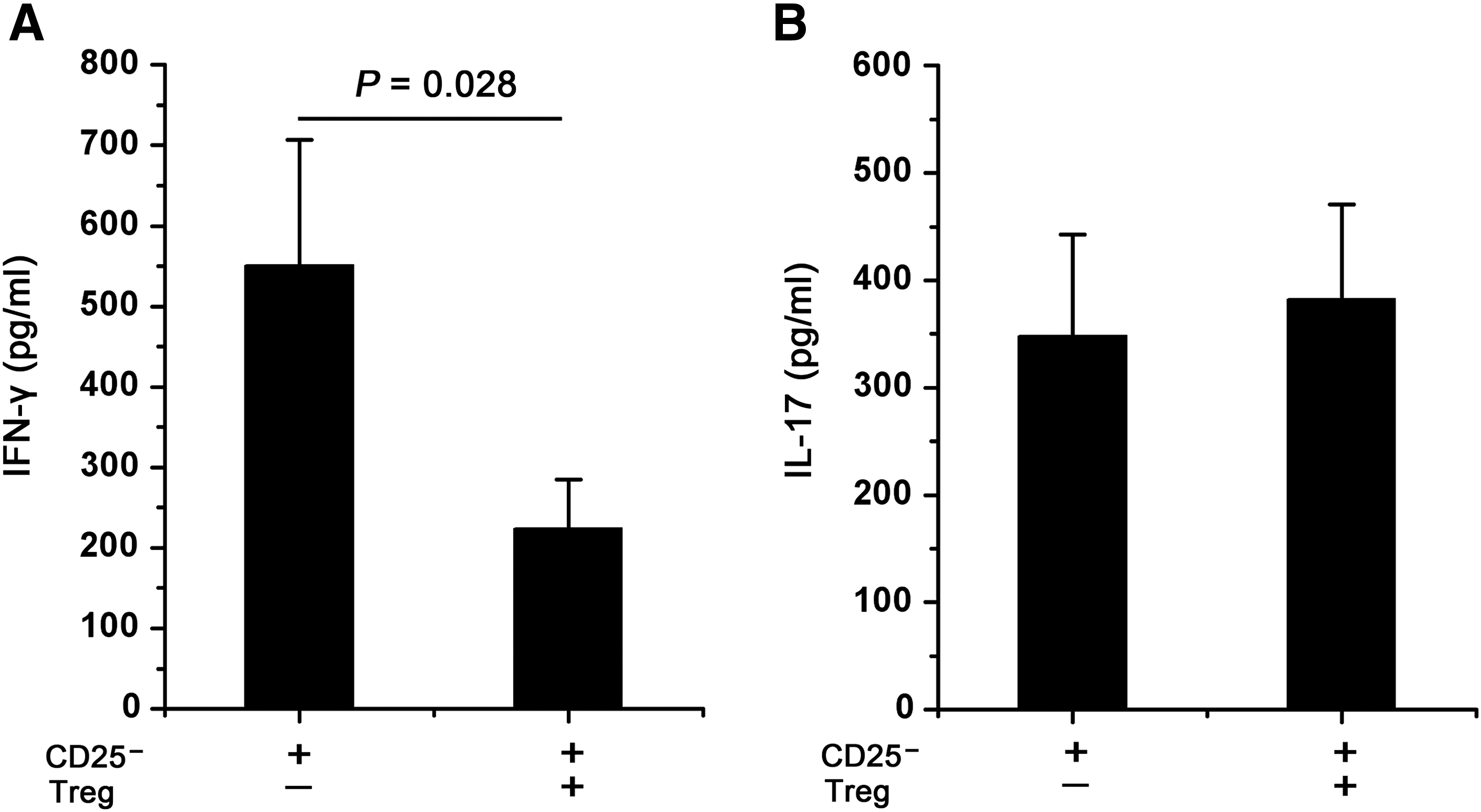
IL-17A secretion by CD4+ T cells is not regulated by Tregs. CD4+CD25− T cells were cultured with Tregs at a 1:1 ratio for CD3/CD28 Dynabead stimulation for 48 h. Culture supernatants were assayed for IFN-γ (
The ratio of Th-17 to Tregs is associated with survival of patients with ACLF
To address whether increased Th-17 and Tregs are associated with ACLF prognosis, 30 ACLF patients were divided into two groups by the final clinical outcome of these patients. The non-survival group includes patients whose condition deteriorated gradually and died within 6 m after blood sampling; however, all the non-survival patients died in 3 mo. The survival group includes patients whose liver function and blood coagulation function reverted gradually after blood sampling in 6 mo, and were still living when the study was completed. There was no significant difference in clinical characteristics between the two groups, such as age, total bilirubin, and prothrombin time. The results showed that the frequency of Th-17 cells in the non-survival group was slightly increased (Fig. 4A), but the frequency of Tregs was slightly decreased in comparison with the survival group (Fig. 4B). However, the differences were not significant. We next compared the ratios of Th-17 to Treg frequency between the two groups. As shown in Fig. 4C, there was a significantly decreased ratio of Th-17 to Treg frequency in the survival group compared with the non-survival group (0.84 ± 0.14 versus 1.08 ± 0.14; p < 0.001). It was striking that 1 out of 14 patients in the survival group had a ratio higher than 1.0. In contrast, the vast majority patients (12 out of 16) in the non-survival group had a ratio higher than 1.0.
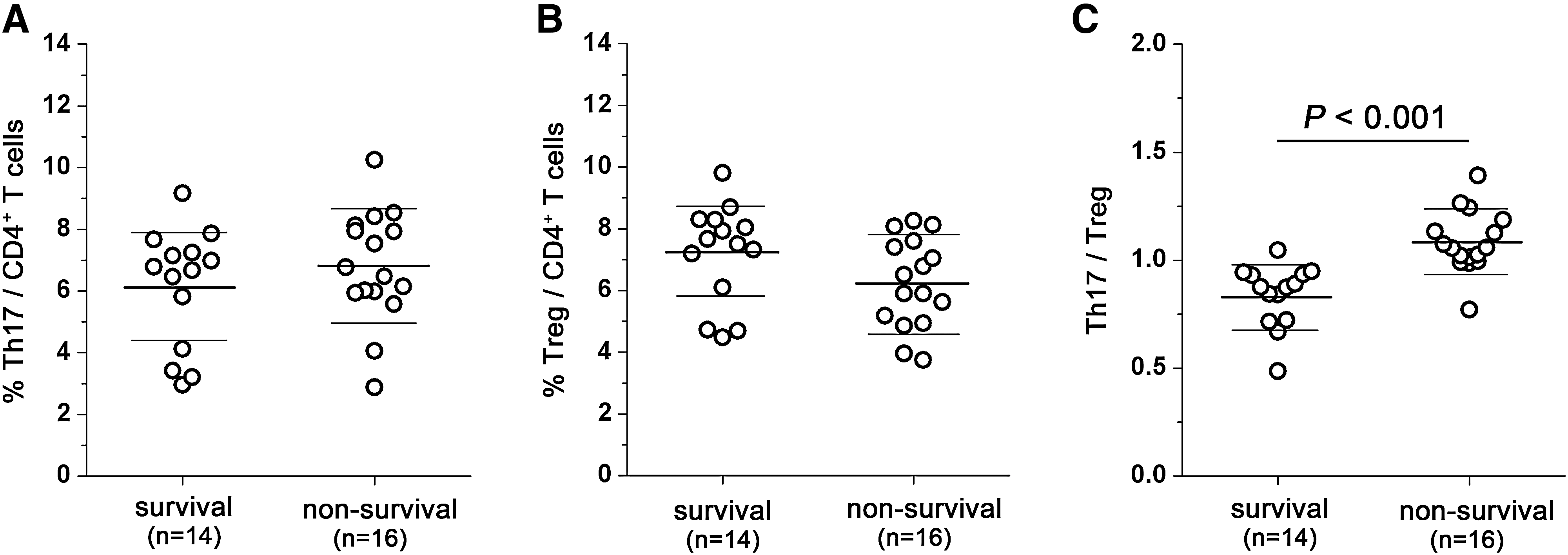
The ratio of Th-17 to Tregs is associated with survival of patients with ACLF. The differences in Th-17 (
Discussion
In the present study, we have demonstrated that HBV-associated ACLF patients have increased proportions of both circulating Th-17 cells and FoxP3+ Tregs. However, Th-17 cells were preferentially increased in ACLF patients compared with Tregs, even though the two subsets of CD4+ T cells were positively correlated with each other. We also show that the IL-17A secretion by CD4+ T cells was not regulated by Tregs. More importantly, we have demonstrated that the ratio of Th-17 to Tregs was associated with the survival of ACLF patients.
Th-17 cells are potent inducers of tissue inflammation and have been associated with the pathogenesis of many autoimmune diseases and inflammatory conditions (18). It has been demonstrated that the Th-17 pathway plays an important role in the development of various liver diseases, including autoimmune liver disease, alcoholic liver disease, hepatocellular carcinoma, and HBV- and HCV-associated liver disease in humans and mice (9,19,21,22,29,30,38,41 –43,45). To date, several reports have shown that Th-17 frequency and the levels of IL-17A in serum of CHB patients are increased and correlate with liver injury (9,38,43,45). Tregs play a central role in inducing and maintaining immunological tolerance and terminating immune responses. The majority of reports on Tregs have studied their influence on agents that usually establish chronic persistence. Conversely, in many chronic infections Tregs appear to restrain immune reactivity in order to limit host tissue damage (1,25).
The close relationship between Th-17 and induced Tregs has been noted, as they share common developmental pathways, with TGF-β essential for their differentiation (18,26). In line with previous studies (38,39,45), we have confirmed that circulating Th-17 and Tregs in HBV-associated ACLF patients are significantly higher than those in healthy individuals and CHB patients. We also found that Th-17 frequencies are significantly increased in CHB patients, and positively correlated with serum ALT levels (data not shown). Peripheral Treg frequency in CHB patients enrolled in the present study is comparable to healthy controls. This result is consistent with the findings of Xu et al. (39), but inconsistent with those of others (33,40). Differences in detection methodology, numbers of enrolled subjects, and clinical therapy may explain the inconsistencies. In addition to the increase in FoxP3+ Tregs, we also found that the frequency of CD25+FoxP3− T cells was significantly increased in ACLF patients (Fig. 1C and data not shown). This probably resulted from excessive immune activation in ACLF patients, since the patients had serious liver damage. A recent report has shown that CD4+FoxP3− T cells are a main source of IL-10-producing cells (27), and IL-10 plays a critical role in counteracting excessive immunity in ACLF patients (48). This may be another regulator of serious liver inflammation in ACLF, and the precise mechanism behind the increasing numbers of CD25+FoxP3− T cells in ACLF patients needs further study.
Although it is now recognized that both Th-17 and Tregs are significantly increased in ACLF patients, our knowledge about the interplay between Tregs and Th-17 cells in HBV-associated ACLF patients is limited. In this study, we found that increases in Th-17 cells are accompanied by FoxP3+ Treg accumulation in CHB and ACLF patients. However, there were more Th-17 cells than Tregs in ACLF patients. More importantly, when we compared the ratios of Th-17 to Tregs, we found that there was a significantly decreased ratio of Th-17 to Tregs in the survival group, and most survival patients had a ratio lower than 1.0. According to these data, we speculate that the interplay between Tregs and Th-17 is important for maintaining the balance between a limited immune response and pathological damage. In ACLF, the increase in Tregs may be due to the negative feedback effects of high levels of Th-17, in addition to the common developmental pathways they share. However, this balance is disrupted by increased Th-17 infiltration compared with Tregs. As a consequence, the uncontrolled inflammation results in end-stage liver disease. The demonstration of this hypothesis needs further research, and similar phenomena have been observed in previous studies of psoriasis and hepatocellular carcinoma (8,44,46). However, there is no association between survival and Th-17 or Treg frequency in ACLF patients. This further demonstrates that the balance between Tregs and Th-17, but neither alone, plays an important role in the clinical outcome of ACLF patients.
To our knowledge, there is little information on whether or not Th-17 responses can be effectively controlled by Tregs in human disease. In this study, we used CD3/CD28 Dynabeads to stimulate CD4+CD25− T cells, and found that Tregs do not downregulate IL-17A production, but do inhibit IFN-γ secretion. We noted that the addition of Tregs resulted in a slight increase in IL-17A production (Fig. 3B). Suppression of IFN-γ secretion by Tregs might account for this, as many studies have demonstrated that IFN-γ was capable of suppressing IL-17A production both in vitro and in vivo (4,10,12,14,17,24,28,35). Notably, a recent study indicated that CD39+ Tregs can suppress pathogenic Th-17 responses, whereas CD39− Tregs produce IL-17A (6). Therefore, the CD39+ Treg would be a good candidate for focus in further studies of ACLF, especially in liver in situ.
In summary, our findings suggest that both Th-17 and FoxP3+ Tregs are increased in ACLF patients, and are positively correlated with each other. However, IL-17A secretion by CD4+ T cells is not regulated by Tregs, even though Tregs exhibit significant inhibition of IFN-γ production. Most importantly, the ratio of Th-17 to Tregs is associated with the survival of ACLF patients. This information furthers our understanding of the associations between Th-17 and Tregs in HBV-associated ACLF and their influence on the final outcome of the disease.
Conclusions
We have shown that Th-17 cells are preferentially increased in ACLF patients, and are positively correlated with Treg numbers. Furthermore, our data indicate that IL-17A secretion by CD4+ T cells is not regulated by Tregs, even though Tregs exhibit significant inhibition of IFN-γ production. Most importantly, we found that there is a significantly decreased ratio of Th-17 to Treg frequency in the survival group, and most survival patients had a ratio lower than 1.0. These findings provide new information regarding the pathogenesis of HBV-associated ACLF, and the ratio of Th-17 to Tregs may represent a potential prognostic marker for the disease.
Footnotes
Acknowledgments
We thank all of the subjects who agreed to participate in this study. This work is supported by National Natural Science Foundation of China grant no. 30670961.
Author Disclosure Statement
No competing financial interests exist.