
Abstract
Influenza viruses cause acute respiratory infections in humans that result in significant excessive morbidity and mortality rates every year. Current vaccines are limited in several aspects, including laborious manufacturing technology, non-sufficient efficacy, and time-consuming adjustments to new emerging virus variants. An alternative vaccine approach utilizes plasmid DNA encoding influenza virus antigens. Previous experiments have evaluated the protective efficacy of DNA vaccines expressing variable as well as conserved antigens. In this present study, several different combinations of influenza A virus (IAV) HA, NA, M1, M2, NS1, NS2, and NP sequences were cloned into the plasmid pVIVO, which allows the independent expression of two genes separately. These DNA vaccines were administered to induce protection against a lethal IAV infection, and to reduce immunopathology in lung tissue of surviving animals. The highest efficacy was provided by vaccines expressing HA and NA, as well as a mixture of plasmids encoding HA, NA, M1, M2, NS1, NS2, and NP (Mix). Three days post-infection, more than a 99.99% reduction of viral load and no inflammation was achieved in lung tissue of pVIVO/HA-NA-vaccinated mice. Animals vaccinated with pVIVO/HA-NA, pVIVO/HA-M2, or vaccine Mix, survived a lethal challenge with minor or no obvious pathologic abnormities in the lungs. All other surviving mice revealed extensive changes in the lung tissue, indicating possibly an ongoing bronchiolitis obliterans. In addition, pVIVO/HA-NA and the vaccine Mix were also protective against a heterologous IAV infection. Taken together, next to all combinations of different DNA vaccines, the intramuscular application of pVIVO/HA-NA was the most efficient procedure to decrease virus replication and to prevent immunopathology in lung tissue of IAV-infected mice.
Introduction
Vaccination is one important strategy to antagonize influenza as a serious public health issue. Despite the use of inactivated or live-attenuated vaccines, and the development of modern therapeutics to interfere with IAV replication, the World Health Organization (WHO) classifies pandemic influenza remains a major global disease. Due to the high genetic variability of the viral genome, occasional influenza pandemics can arise at any time (36). Moreover, the emergence of new virus variants—like the highly pathogenic avian H5N1 or the new swine H1N1 subtype—demands even more scientific efforts worldwide to antagonize IAV infections. Current vaccination procedures to prevent IAV infections are based on traditional vaccines, which are generated either from IAV-infected eggs or cell cultures. These manufacturing processes are expensive and time-consuming, and make it difficult to respond quickly to new emerging virus variants. Therefore, the search for novel vaccination procedures against IAV infection is one of the important research topics worldwide.
A fast-developing class of novel vaccines, based on immunization with plasmid DNA, may be promising to protect individuals against disease without the disadvantages of current vaccines (8). Plasmids can be developed and produced in a very short time, which may help to respond to new influenza epidemics or pandemics very promptly. DNA immunization represents an attractive alternative to induce robust humoral and cellular immune responses (37). This technique requires the in vivo administration of DNA encoding immunogenic viral proteins, which results in the synthesis of those proteins and is followed by protective immune responses (26). Here, DNA administration via electroporation represents a very efficient application method (13). Today DNA vaccine research has already progressed from laboratory tests to licensed animal vaccines (27).
Using several different animal models, DNA immunization has been shown to be effective against many pathogens, including IAV (4,39). DNA vaccines against IAV infections revealed that this procedure is capable of eliciting protective immune responses following delivery by various routes (12,29,38). Moreover, promising results of phase 1 clinical trials to analyze the safety and immunogenicity of such an IAV-specific vaccine were recently published (21,33). Despite these encouraging data, the successful application of this method in humans will be highly important for the future of DNA vaccine development in general. Therefore, increased efficacy and maximal safety are the major issues of DNA immunization today. Notably, the National Institutes of Health (NIH) and the WHO have recommended an emphasis on developing such an influenza DNA vaccine (14,37).
The IAV genome consists of 8 RNA segments that encode 10 to 11 proteins: hemagglutinin (HA), neuraminidase (NA), nucleoprotein (NP), matrix proteins 1 and 2 (M1 and M2), nonstructural proteins 1 and 2 (NS1 and NS2), and polymerase subunits PA, PB1, and PB2, as well as the apoptotic protein PB1-F2, which is expressed in some IAV strains (5). Natural protection against IAV infections is basically mediated by immune reactions against the two major antigenic surface proteins HA and NA. Several previous studies have confirmed that DNA vaccines encoding HA (1,12,20,43) or NA (2,16), as well as a combination of HA/NA (6,45), induced specific and protective immune responses. In terms of protection, other DNA vaccines encoding different IAV proteins were analyzed as well, including NP (39), M2 (19), M1 (44), and NS1 (46).
Previously it was demonstrated that the expression of HA and NA together with interleukin-2 (IL-2) via bicistronic plasmids induced protection in more than 80% of IAV-infected mice (16). In order to increase vaccination efficacy even more, the present study evaluates the protective capability of different DNA vaccines encoding IAV sequences of HA, NA, M1, M2, NS1, NS2, and NP individually or in combination. Here, a DNA vector was used that contains two transcription units allowing the combined but independent expression of two genes of interest from a single plasmid. One specific focus was directed towards pathologic changes in lung tissue of vaccinated animals that survived the lethal IAV challenge.
Materials and Methods
Cell culture
MDCK cells (ATCC no. CCL-34) were cultured in Eagle's Minimum Essential Medium (EMEM), supplemented with 10% fetal calf serum (FCS), 100 U/mL penicillin, 100 μg/mL streptomycin sulfate, 2 mM L-glutamine, and 1% non-essential amino acids at 37°C and 5% CO2-containing humidified air.
Virus
A mouse-adapted variant of the H1N1 IAV strain PR/8/34 was kindly provided by Jürgen Stech (Federal Research Institute of Animal Health, Riems, Germany). The mouse-adapted variant of the H1N1 IAV strain swine/Potsdam/15/81 was generated by Michaela Schmidtke (Department of Virology and Antiviral Therapy, University Hospital Jena, Germany). Viral propagation was performed on MDCK cell monolayers in EMEM without FCS, supplemented with 2 μg/mL trypsin (EMEM/trypsin). Aliquots of virus stocks were stored at −80°C.
Mice
In this study, male inbred BALB/C mice 6–8 wk of age were obtained (Charles River Laboratory) and used throughout the experiments. Test groups consisted of 3–4 mice. The experiments were usually carried out three times. Animal experiments complied with all federal permissions, guidelines, and international policies.
Eukaryotic expression plasmid construction
All DNA vaccines were constructed by inserting the IAV open reading frame sequences of HA, NA, M1, M2, NS1, NS2, and NP, either alone or in combination, into the expression plasmid pVIVO2-mcs (pVIVO; InvivoGen, Toulouse, France), which contains the human ferritin heavy (hFerH) and light (hFerL) promoters, the 5′-UTR of the mouse (m), and chimpanzee (ch) elongation factor 1 (EF1), the SV40 enhancer, the CMV enhancer, the SV40 and EF1 polyadenylation signal, and the hygromycin resistance gene. Two multiple cloning sites (mcs) are located downstream of the hFerH or hFerL, and the mEF1 or chEF1 5′-UTR, respectively. This permits the translation of two different genes independently. Prior construction of DNA vaccines, and equal expression from both mcs were confirmed using two different reporter proteins. Here, the enhanced green fluorescent protein (EGFP) was cloned into the mcs 1, and the red fluorescent protein from Discosoma sp. (DsRed2) was cloned into the mcs 2. The obtained plasmid pVIVO/EGFP-DsRed2 was used to transfect HEK 293 cells. The expression from both mcs was analyzed by fluorescence microscopy 24 h later, indicating equal expression from both sites (data not shown). Based on this observation, HA, M1, or NS1 were cloned into the mcs 1, and M2, NA, NS2, or NP were cloned into the mcs 2. Then viral RNA was reverse transcribed into single-stranded cDNA (Invitrogen, Carlsbad, CA) using RNA samples of IAV PR/8/34-infected MDCK cells. Virus sequences were subsequently amplified by PCR and cloned into pVIVO to generate pVIVO/M1, pVIVO/M2, pVIVO/NS1, pVIVO/NS2, pVIVO/NP, pVIVO/HA-M2, p/VIVO/NP-M1, pVIVO/NS1-NS2, and pVIVO/HA-NA. The correctness of all constructs was confirmed by sequence analysis. Plasmid isolation from bacteria was performed using an endotoxin-free plasmid preparation kit (Qiagen, Inc., Valencia, CA).
Transient transfection
Human HEK-293 cells (ATCC no. CRL-1573) were grown in EMEM supplemented with 10% FCS, 100 U/mL penicillin, 100 μg/mL streptomycin sulfate, 2 mM L-glutamine, and 1% non-essential amino acids until 40–60% confluence was achieved. Then transient transfection was performed using the Effectene reaction kit (Qiagene) according to the manufacturer's instructions. After 48 h samples were taken and either used for protein isolation or immunocytochemistry.
Protein detection
Western blot analysis
Protein was extracted from transfected cells using NTE-buffer (100 mM NaCl, and 10 mM Tris/HCl, pH 7.4), and 10% NP-40, and were stored at −80°C. Then concentrations were determined with the Bradford method and Coomassie brilliant blue G250 (BioRad Laboratories, Inc., Hercules, CA). Protein samples (150 μg) were incubated for 5 min at 95°C, separated by 12% sodium dodecyl sulfate polyacrylamide gel electrophoresis (SDS-PAGE), and blotted onto Protran® nitrocellulose membranes (Schleicher & Schuell, Riviera Beach, FL). Non-specific binding sites were blocked with skim milk incubation. Tris-buffered saline plus Tween 20 (150 mM NaCl and 10 mM Tris/HCl, pH 8.0, and 0.05% Tween 20) was used for all washing procedures. Depending on the experiments, different combinations of primary and secondary antibodies were used. Proteins were visualized using the NBT/BCIP detection system (Roche, Diagnostics GmbH, Mannheim, Germany).
Immunocytochemistry
Non- or plasmid-transfected MDCK cells were grown on sterile cover-slips in six-well cell culture plates. Three days after transfection, the slides were transferred into new wells and fixed with 4% paraformaldehyde at 4°C for 20 min. Then the slides were washed twice with PBS, and the cells were permeabilized with cold acetone/methanol (1/2 vol) for 10 min at 20°C. All following incubation steps were accomplished at room temperature. Depending on the experiments, different combinations of primary and secondary antibodies were used. All slides were washed and mounted with Aquatex (Merck, Darmstadt, Germany), and positive cells were analyzed with light or fluorescence microscopy.
Primary antibodies
anti-IAV to detect HA and NA (1:500, anti-IAV polyclonal mouse serum, in-house); anti-NP (1:500; Santa Cruz Biotechnology, Santa Cruz, CA); anti-M1 (1:100; Acris); anti-NS1 (1:500; Santa Cruz Biotechnology); anti-NS2 (1:100; Santa Cruz Biotechnology); anti-M2 (1:1000; Biozol Diagnostica, Eching, Germany); and anti-β-actin (1:1000; Cell Signaling Technology, Inc., Beverly, MA).
Secondary antibodies
Rabbit anti-mouse IgG alkaline phosphatase (AP)-conjugated (1:2000; Cell Signaling); goat anti-rabbit IgG AP-conjugated (1:1000; Acris, Herford, Germany); rabbit anti-goat IgG AP-conjugated (1:1000; Acris); goat anti-mouse IgG Cy3-conjugated (1:400); and donkey anti-goat IgG fluorescein-conjugated (1:300) antibodies.
Immunization Protocol
Anesthetized BALB/C mice received bilateral intramuscular injections (50 μg DNA in 50 μL saline per leg, a total of 100 μg) in each musculus quadriceps femoris on day 0 and day 14. In case of the administration of mixed DNA vaccines (Mix), equal amounts of pVIVO/HA-NA, pVIVO/NP-M1, pVIVO/NS1-NS2, and pVIVO/M2 (25 μg each) were mixed and separated into two 50-μg doses prior to injection. On day 28, anesthetized mice were challenged with IAV.
Influenza A virus infection in vivo
Isoflurane-anesthetized mice were infected intranasally (IN) with 5 LD50 doses of IAV in 50 μL saline. The mice were monitored daily for body weight and survival. Animals found in moribund condition (loss of weight, ruffled fur, complete unresponsiveness, or crooked posture) were euthanized and were considered to have succumbed to infection on the day of euthanasia.
Quantitative real-time RT-PCR (qRT-PCR)
Total RNA was extracted from lung tissues using the phenol chloroform method (7). RNA quantities were analyzed and equal amounts were subjected to reverse transcription. After annealing of oligo (dT) primers for 10 min at 70°C, the samples were incubated for 50 min at 42°C using 100 U Superscript II reverse transcriptase (Invitrogen), and the appropriate buffer containing 100 mM DTT and 10 mM dNTPs. The reaction was terminated by heating. Thereafter, levels of relative gene expression of the IAV HA gene in comparison to murine β-actin as internal control were analyzed by SYBR Green-based quantitative PCR (Qiagen), using LightCycler 3 technology (Roche). Primer combinations were 5′-GAAGTGTCAAACACCCCTGGGAGC-3′ (sense) and 5′-CCGGCAATGGCTCCAAATAGACC-3′ (anti-sense) for HA, and 5′-TGGGCGACGAGGCCCAGAGCAAGAAG-3′ (sense) and 5′-GCTCATTGTAGA-AGGTGTGGTGCCAG-3′ (anti-sense) for β-actin. First, the qRT-PCR was validated for comparable PCR efficiencies between HA and β-actin expression according to the supplier's instructions (Roche). Thereafter, amounts of HA sequences in samples of IAV-infected mice were analyzed using a relative quantification method (30).
Histology
At 3 or 14 d post-infection (dpi) murine lungs were removed, embedded in paraffin, and stained with hematoxylin and eosin. Thereafter, tissue was examined for the presence of inflammatory infiltrates and pathologic tissue destruction by light microscopy.
Immunohistochemistry with frozen sections
Immunohistochemical studies were carried out with cryomicrotome sections of lung tissue (8 μm) as described previously (15). Primary antibody was applied for 1 h. This consisted of the rat anti-mouse Ly6B.2 alloantigen, and monoclonal antibody (MCA771GA, clone 7/4, 1:50; AbD Serotech, Raleigh, CA). The polymorphic 40-kD antigen Ly6B.2 is expressed by polymorphonuclear granulocytes. Thereafter, a secondary Cy3-conjugated goat anti-rat IgG antibody (1:100; Jackson ImmunoResearch Laboratories, West Grove, PA) was used.
Results
Construction of DNA vaccines
In order to obtain the necessary sequences, MDCK cells were infected with IAV PR/8/34. After the appearance of cytopathic effects, RNA was isolated and reverse transcribed into cDNA. Full-length amplicons of HA, NA, M1, M2, NS1, NS2, and NP sequences were generated and cloned into the plasmid pVIVO. This vector system allows the simultaneous expression of two genes of interest at high levels by cloning them into two different multiple cloning sites of one plasmid. The following plasmids were obtained: pVIVO/M1, pVIVO/M2, pVIVO/NS1, pVIVO/NS2, pVIVO/NP, pVIVO/HA-M2, p/VIVO/NP-M1, pVIVO/NS1-NS2, and pVIVO/HA-NA. After transient transfection, translational activity was determined by Western blot analysis or by immunocytochemistry detection (data not shown).
Impact of vaccination on acute infection
After the expression of IAV sequences by those DNA vaccines was confirmed in vitro, male BALB/C mice were plasmid-inoculated in each quadriceps muscle separately. Control mice received pVIVO or saline only. Two weeks later the mice were boostered with the same DNA and were challenged with 5 LD50 doses of IAV PR/8/34 after two additional weeks. The lethality of this challenge was confirmed during previous experiments. In order to analyze the effect of the application of different DNA vaccines on the virus replication in lung tissue of IAV-infected mice, vaccinated as well as control animals were sacrificed 3 d post-challenge. Samples of lung tissue were obtained and RNA was isolated and transcribed into cDNA. Successful cDNA synthesis was controlled via detection of the housekeeping gene β-actin by qRT-PCR. Thereafter, the amount of HA expression was analyzed and normalized to the expression of β-actin, as shown in Fig. 1. The most efficient protection was achieved when mice were immunized with pVIVO/HA-NA. In lung tissues of these animals, the lowest concentration of viral RNA was detected in comparison to pVIVO-treated control mice. This vaccination decreased the virus load more than 3000-fold. Other vaccines, like pVIVO/NP, pVIVO/HA-M2, and the vaccine mixture (Mix), were successful as well, but without statistical significance. Here, reductions of 38-, 43-, and 100-fold were observed. All the other plasmid DNA administrations had no influence (pVIVO/M1 and pVIVO/NS1-NS2), or only marginal effects (pVIVO/M2, pVIVO/NS1, pVIVO/NS2, and pVIVO/NP-M1) on the viral load in lung tissue after IAV challenge. These results were confirmed by experiments to analyze the impact of IAV infection on the integrity of murine lung tissue following different vaccine applications. For this, lung samples were obtained 3 dpi and analyzed microscopically in view of tissue destruction. As shown in Fig. 2, NaCl-treated and virus-infected mice developed severe pathologic changes typical for IAV PR/8/34 throughout the lung tissue, spreading widely from blood vessels. Similar reactions were observed in the tissue of pVIVO-, pVIVO/NP-, pVIVO/NS2-, pVIVO/NP-M1-, and pVIVO/NS1-NS2-inoculated mice. Reduced infiltration and tissue damage were detectable in the lungs of pVIVO/M1- and pVIVO/NS1-vaccinated animals. In contrast, mice that were immunized with the DNA constructs pVIVO/M2, pVIVO/HA-M2, pVIVO/HA-NA, or Mix, revealed the same healthy tissue structure as non-infected controls, demonstrating that these plasmids were able to induce very efficient protection during acute IAV PR/8/34-caused disease. But IAV PR/8/34 may overcome this early reduction of virus replication in vaccinated mice. Therefore, the next set of experiments focused on the outcome of IAV infection after 2 wk.
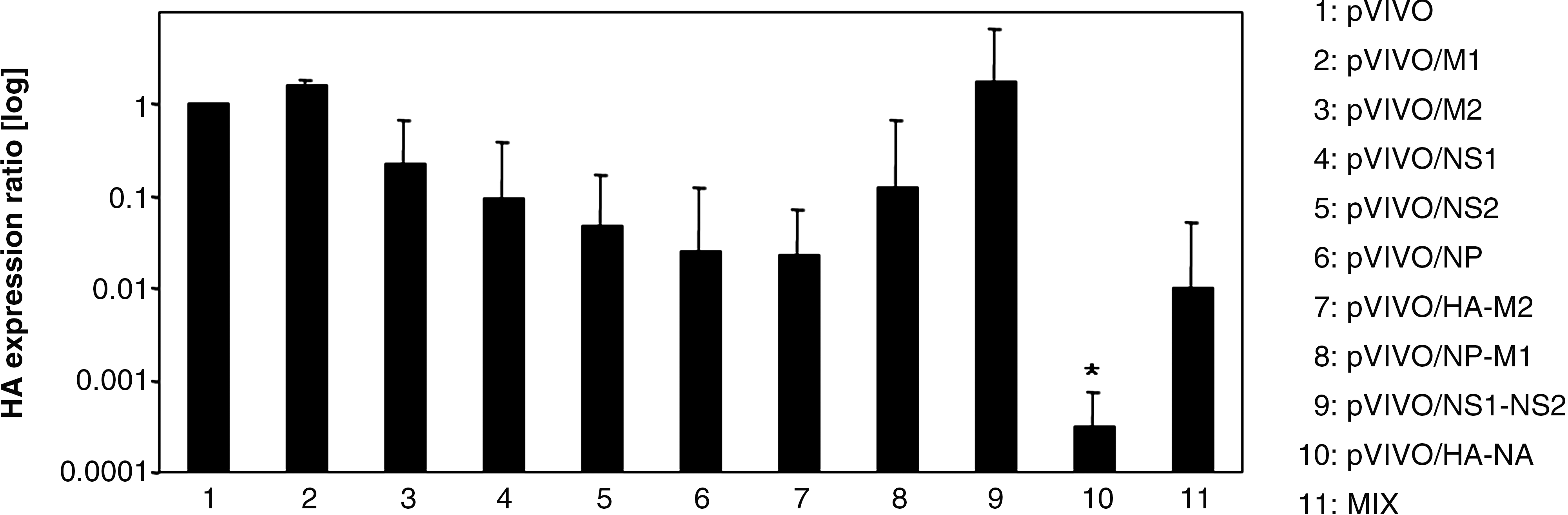
Male BALB/C mice were inoculated with different plasmids to analyze the influence on IAV replication. Three days after challenge with 5 LD50 doses of IAV, lung samples were obtained and analyzed in view of the amount of viral HA by qRT-PCR. Data were normalized to individual β-actin expression, compared to pVIVO-treated, IAV-infected control mice (set as 1), and are shown as mean relative values ± standard deviation of three experiments. A significant decrease in viral load between control mice (group 1) and pVIVO/HA-NA-immunized mice (group 10) is clearly indicated (*p < 0.05).
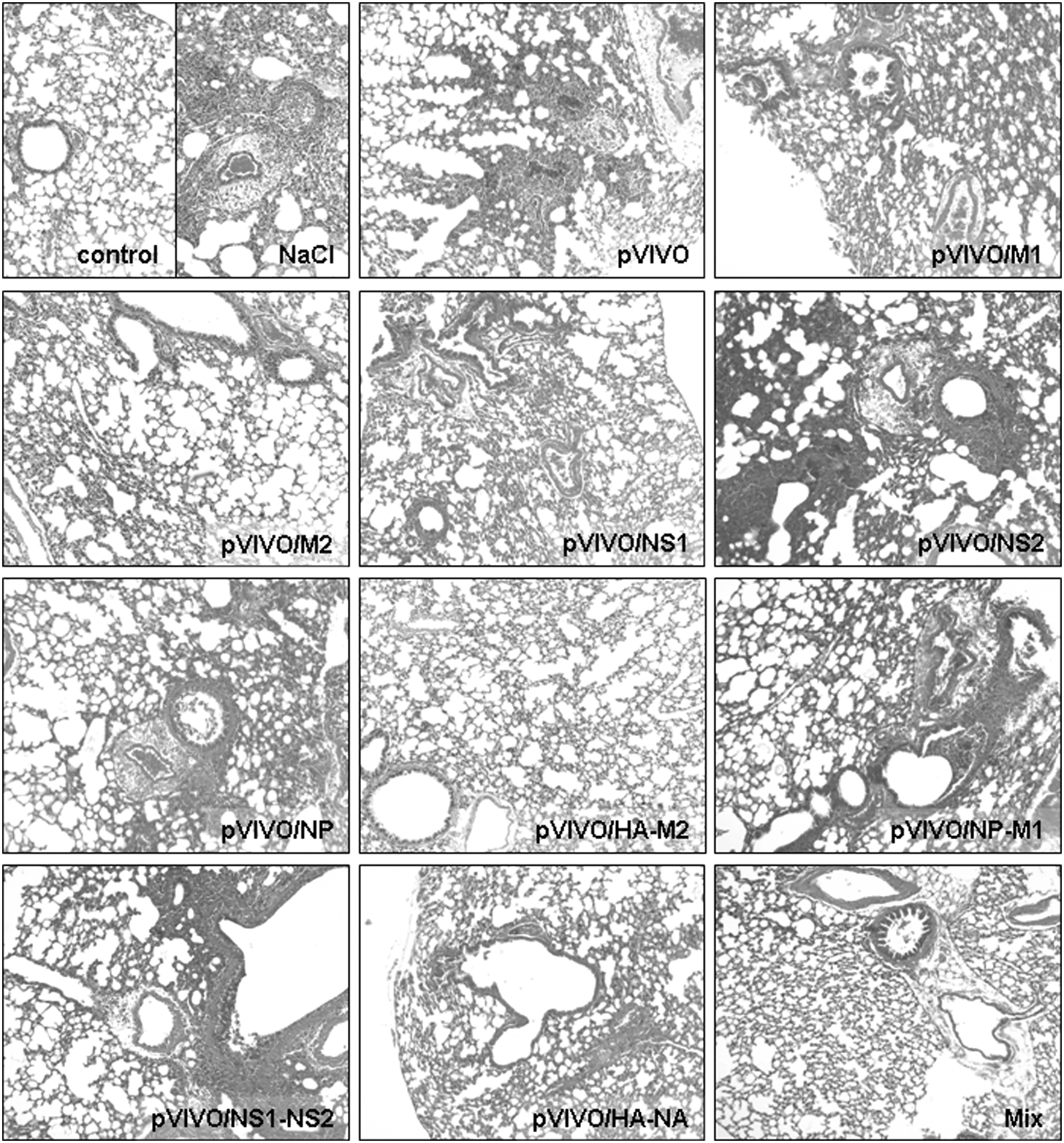
Male BALB/C mice were inoculated with different plasmids to study the effect on IAV-induced inflammation. Three days after challenge with 5 LD50 doses of IAV, lung samples were obtained, fixed with formalin, stained with hematoxylin and eosin, and analyzed microscopically concerning the degree of virus-caused pathology. Representative sections are shown (magnification 100 × ).
Protection against lethal IAV PR/8/34 infection
Here, groups of BALB/C mice were treated according to the immunization protocol. After viral challenge, the mice were monitored closely up to 2 wk for any signs of illness and death. Previous experiments revealed that after 12 dpi no further death could be detected using this animal model. Four to five days post-challenge non-protected mice revealed typical symptoms of acute disease, as characterized by weight loss, ruffled fur, unresponsiveness, and a crooked posture. Normally, after administration of 5 LD50 doses of IAV, all non-vaccinated mice succumbed to death from 3–7 dpi, as shown in Fig. 3. Mice that received only saline or pVIVO died quickly 3–6 or 4–7 dpi, respectively. Prolonged survival was detectable in pVIVO/NS2-injected mice. Here, all animals died from 8–9 dpi. All other immunized mice revealed protection between 33 and 100%. Partial protection was generated in mice that were vaccinated with pVIVO/M2, pVIVO/NS1, pVIVO/NP, pVIVO/NS1-NS2, or pVIVO/NP-M1 (all 33% survival), as well as pVIVO/M1 (66% survival). The most efficient vaccines were pVIVO/HA-NA, pVIVO/HA-M2, and the vaccine Mix. In these groups, no mice succumbed to death due to the lethal challenge.
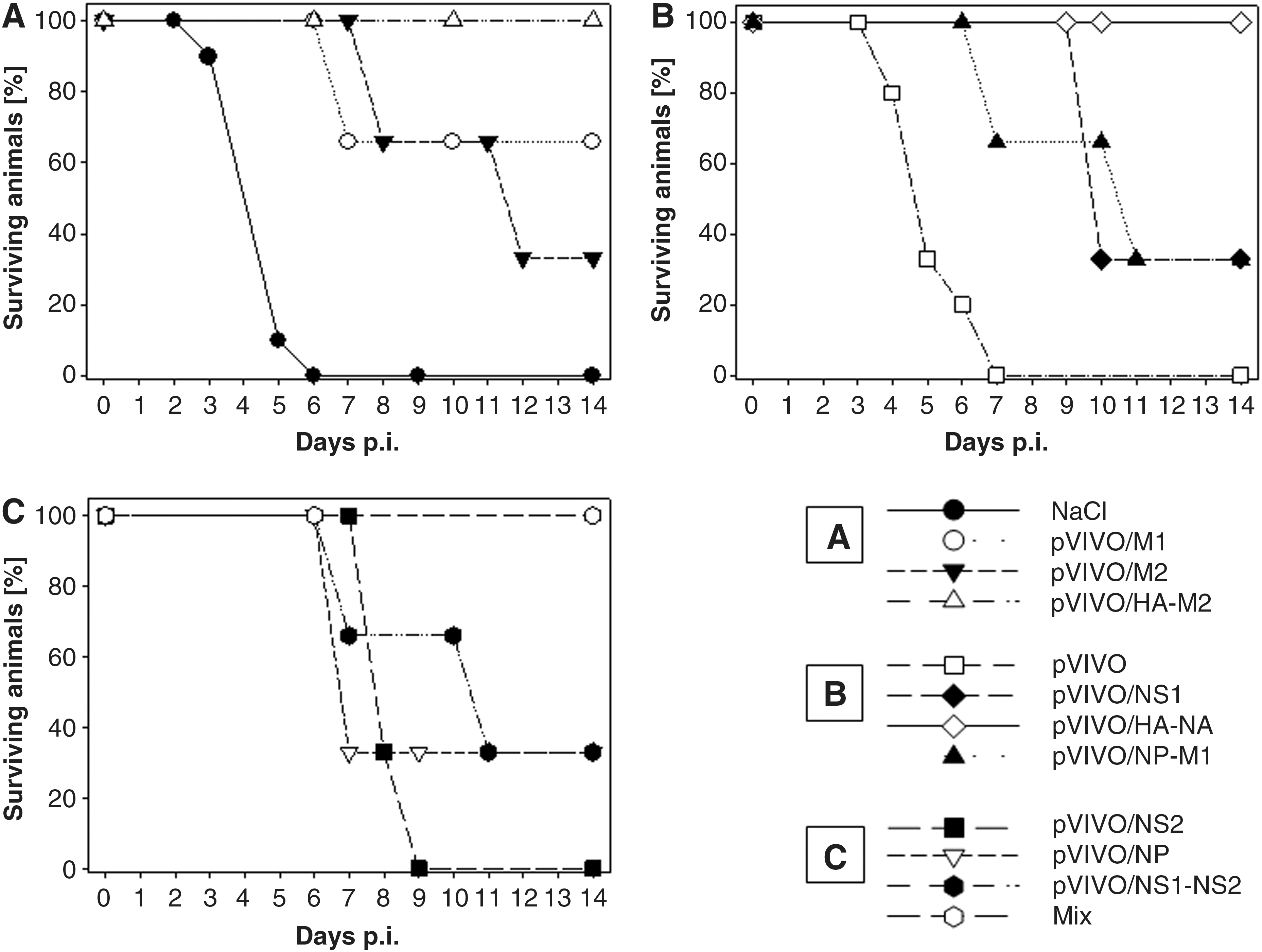
Male BALB/C mice were vaccinated with different plasmids (
Impact of vaccination on the state of health of surviving animals
Surviving animals were sacrificed 14 dpi. At this time all mice appeared healthy, regained weight, and did not show any signs of disease. Interestingly, this observation was not reflected by the histological condition of lung tissue, as shown in Fig. 4A. Surviving animals of the pVIVO/M1-, pVIVO/NP-, pVIVO/NS1-, pVIVO/M2-, pVIVO/NP-M1-, and pVIVO/NS1-NS2-immunized groups revealed extensive changes in the lung tissue. Large areas were characterized by a complete loss of the typical lung structure. Most of the bronchioles appear to be plugged. In contrast, lung tissue from mice that received pVIVO/HA-M2 prior to challenge revealed only minor pathology, and the tissue of pVIVO/HA-NA- and Mix-vaccinated mice remained completely intact like the lungs of non-infected control mice (for comparison see Fig. 2). No signs of lung destruction were detectable in these tissue sections, indicating that those immunized mice were protected not only against acute IAV-caused disease, but also against acute and ongoing inflammatory responses, which may induce a chronic non-reversible obstructive lung disease. As an example, two different magnifications that show this are presented in Fig. 4B (the upper series of images are low magnification [Fig. 4A], and the insets show them at higher magnification in Fig. 4B). Tissue samples of surviving mice clearly revealed the difference between morbid (pVIVO/NS1-NS2) and healthy (Mix) tissues. Surviving IAV-infected animals that received the vaccine Mix revealed no differences in the histology of lung tissue in comparison to the non-infected controls. This also applies to pVIVO/HA-NA-immunized mice at 14 d post-challenge. In addition, corresponding cryomicrotome sections of the same lung tissues were analyzed in view of the nature of infiltrating cells (lower series of images in Fig. 4B). Here, an intensive staining of Ly6B.2-positive cells is only detectable in morbid tissues of surviving pVIVO/NS1-NS2-immunized mice.
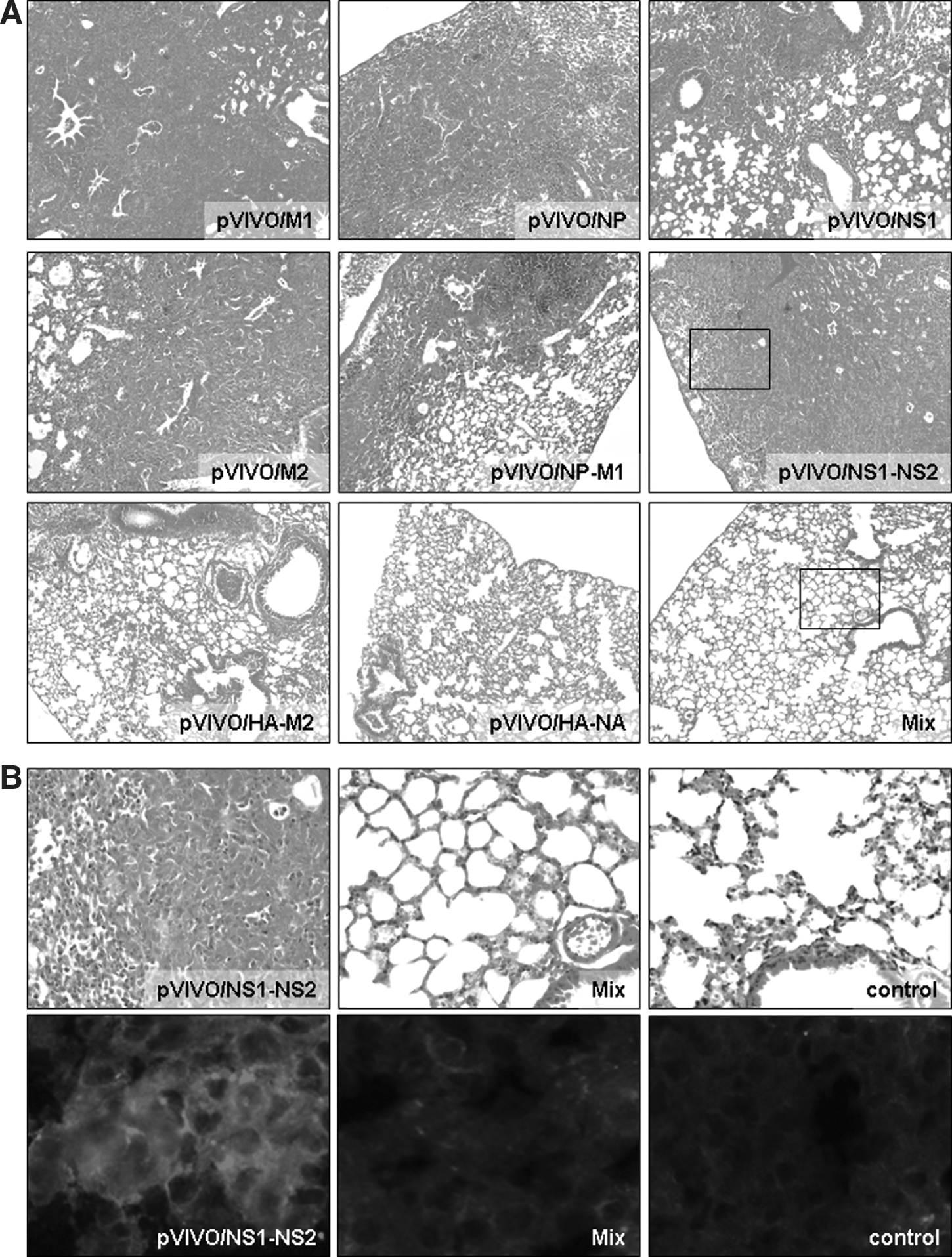
Male BALB/C mice were vaccinated with different plasmids to study the influence of these treatments on lung pathology of surviving animals. Control mice received only NaCl. Two weeks after challenge with 5 LD50 doses of IAV, lung samples of surviving animals were obtained, fixed with formalin, stained with hematoxylin and eosin, and analyzed microscopically for the degree of virus-caused pathology (
Interestingly, in lung tissues of surviving animals, only in the pVIVO/M2-immunized group were low amounts of IAV gene sequences detected 2 wk post-infection. However, direct isolation of replicating virus or detection of IAV RNA sequences from tissue samples was not successful, indicating that at 14 dpi no virus remained in lung tissues of all other surviving animals (data not shown).
DNA vaccination induced protection against heterologous infection
In order to analyze if immunization with two of the most effective vaccines induces protection against a heterologous IAV strain as well, animals were inoculated twice with pVIVO (as control), or with pVIVO/HA-NA or the vaccine Mix, according to the immunization protocol. Then, mice were challenged with 5 LD50 doses of either IAV PR/8/34 or IAV swine/Potsdam/15/81 and monitored in view of body weight and survival for a period of 2 wk. After virus infection, all control mice that received pVIVO succumbed to death from 7–10 dpi in the case of IAV PR/8/34, or 3–5 dpi in the case of IAV swine/Potsdam/15/81. In contrast, all immunized animals survived the lethal challenge with the homologous IAV/PR/8/34. In addition, DNA vaccines caused protection in 66% of mice after infection with the heterologous IAV swine/Potsdam/15/81. During a period of 14 d, body weights of all challenged mice were monitored individually. As demonstrated at Fig. 5A, all vaccinated animals lost 5–15% of their body weight, with a maximum seen between 6 and 9 dpi with IAV PR/8/34. It appeared that Mix-vaccinated mice were slightly less affected by IAV PR/8/34 than pVIVO/HA-NA-immunized mice. After 2 wk all animals regained their original weight independently of vaccine administration. Fig. 5B demonstrates the changes of body weight of mice that were challenged with the heterologous virus. Obviously, infection with IAV swine/Potsdam/15/81 caused an increased body weight loss, starting at 1 dpi, and with maximal values of 15–35% seen at 4–7 dpi. Here, pVIVO/HA-NA-immunized mice appeared less affected by IAV swine/Potsdam/15/81 than Mix-vaccinated mice. But at the end of the experiment, all surviving animals had regained most of their original weight.
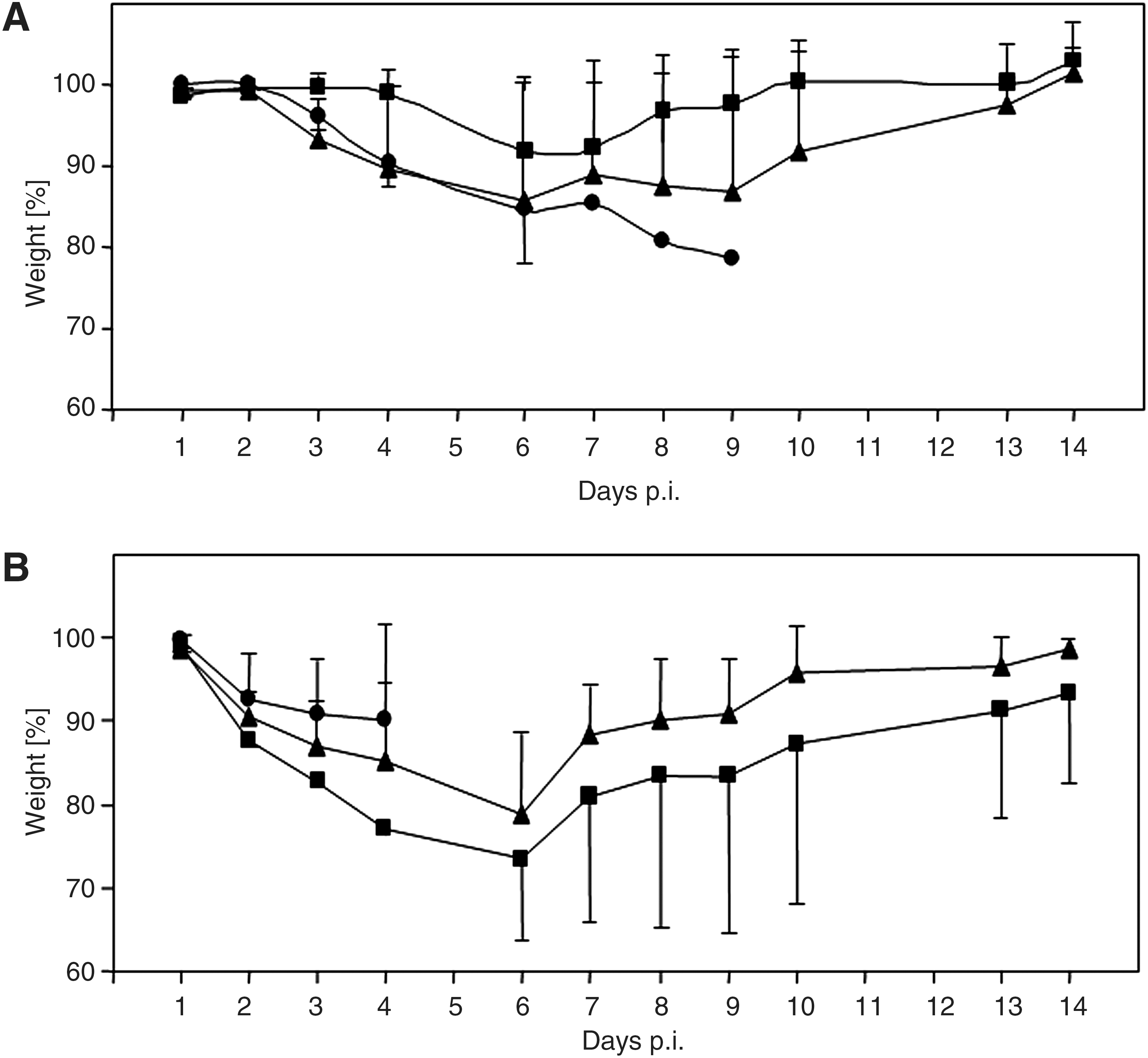
Male BALB/C mice were inoculated with pVIVO (circles), pVIVO/HA-NA (triangles), or Mix (squares). During the 2 wk after challenge with 5 LD50 doses of either IAV/PR/8/34 (
Discussion
At present, the majority of the licensed vaccines against IAV infections are based on inactivated antigen preparations. Clinical studies have shown that such vaccines have an efficiency of an average of 70% in people between 14 and 60 y of age (9), but efficacy is reduced in infants and elderly people (17). Therefore, potent vaccines capable of conferring broad protection against both seasonal and pandemic IAV strains are urgently needed. In order to enhance the potency of influenza vaccination in general, several approaches are under investigation today (reviewed in 10). Among these, DNA vaccination presents a rather new strategy to prevent influenza. Plasmids expressing viral HA or NA in combination with other IAV genes or immunostimulatory cytokines have been studied in different animal models, demonstrating the efficacy of this method (11,16,24,31).
In the present study, the value of different DNA vaccines to induce protection and to reduce inflammatory reactions in lung tissue of IAV-infected mice was analyzed in the context of the expressed viral gene. During acute viral infection the most protective effect could be observed in pVIVO/HA-NA-immunized mice. This was reflected by significantly reduced IAV replication and the absence of inflammatory cell infiltration. Comparably high protective efficiency was also reported in IAV-infected animals after administration of bicistronic plasmids expressing HA and NA simultaneously (45). Other experiments demonstrated that the single expression of either HA or NA was less sufficient to protect IAV-infected mice (16).
In lung tissue of IAV PR/8/34-infected control mice, typical perivascular infiltrations of immune cells into interstitial regions, and areas of focal necrosis of terminal airways, as well as acute hemorrhage into alveoli were observed 3 dpi (Fig. 2A). This situation was also detectable in lung tissues of pVIVO-, pVIVO/M1-, pVIVO/NS1-, pVIVO/NS2-, pVIVO/NP, pVIVO/NP-M1-, and pVIVO/NS1-NS2-immunized mice (Fig. 2), and was generally accompanied by a severe viral load. However, lung tissues of pVIVO/M2-immunized mice revealed almost no inflammation, but also no substantial decrease in IAV replication. This observation may reflect the previously reported low immunogenicity of M2 (18), which, in addition, may also depend on the genetic control of the infected host (28). Additional DNA-based expression of either NP or HA improved the protective value of M2 (28,44). However, other DNA vaccines offered much higher potentials to protect against IAV infections (Figs. 1 –4). Here, lung tissues of mice that received pVIVO/HA-M2 prior to challenge revealed only minor pathology, and in mice vaccinated with pVIVO/HA-NA or the vaccine Mix prior to challenge, the low viral titer was reflected by the complete absence of detectable immunopathology.
Challenge experiments in mice immunized with different DNA plasmids revealed the protective potential of each vaccine. As expected, all non-inoculated as well as pVIVO-inoculated mice died during the observation period, together with mice of the pVIVO/NS2 group, indicating that no protective immune response was generated. This observation is in line with the results of experiments done to characterize the generation of virus-specific antibodies in vaccinated pigs to certain swine influenza virus proteins (23). The administration of plasmids expressing M1, M2, NS1, NP, NP-M1, or NS1-NS2 was only somewhat effective, verifying the former results (3,19,46). But lung tissues of most of these surviving animals revealed massive changes in the normal lung structure, which were, in contrast, low in lungs of pVIVO/HA-M2-inocluated animals, and absent in lungs of pVIVO/HA-NA- and Mix-inoculated animals 2 weeks post-infection. Evidence of this is that immunostaining of such tissue sections indicated an increased number of Ly6B.2-positive cells in the lungs of pVIVO/NS1-NS2-vaccinated mice 2 wk after challenge (Fig. 4B). This protein is expressed by granulocytes but is absent on resident tissue macrophages. The observed immunopathology in lung tissues of some surviving animals could be explained by an ongoing inflammatory response, which is typical for bronchiolitis obliterans (BO). This term describes an inflammation of the bronchioles causing a partial/complete obliteration of the airways. It is known that viral infections represent a distinct risk for patients of developing BO, followed by death (22). The clinical role of influenza A and B viruses or respiratory syncytial virus (40), as well as Epstein-Barr virus (25), in inducing BO in lung transplant patients was discussed previously. Moreover it has been shown recently that IAV could be involved in BO development in humans, and appropriate treatment for BO improved the patients' conditions (34). Such observations raise the question if individuals who survived an initial IAV infection, but developed severe pathological changes in lung tissue possibly connected with BO, may succumb to death without appropriate treatment later on. Further experiments using the well established model of IAV-infected mice should focus on this question.
Finally, two of the most efficient vaccines—pVIVO/HA-NA and Mix—were analyzed in view of their protective capability against a heterologous IAV infection. Here, the mouse-adapted IAV swine/Potsdam/15/81 isolate was used, which has been studied previously to determine its drug susceptibility (32). In comparison to IAV PR/8/34 (Fig. 5A), this IAV variant appeared more virulent in mice, because non-immunized mice died earlier and immunized mice lost more weight during challenge experiments (Fig. 5B). Nevertheless, both vaccine applications induced notable protection against a heterologous infection, and surviving animals regained most of their weight by 14 dpi.
Conclusion
Here, a set of DNA plasmids to express different IAV sequences were analyzed in view of their capability to protect against homologous or heterologous IAV challenges. Among those vaccines, vectors encoding HA and NA, or a combination of HA, NA, M1, M2, NS1, NS2, and NP, revealed the best protective efficacy. But to identify an ideal vaccine, one additional specific aspect should be focused on, namely the potential degradation processes seen in the lungs of challenged individuals after vaccination. Therefore, decreased viral replication and host survival alone might not be enough to characterize the potential of a certain vaccine. These results might be important in view of the design and characterization of new human IAV vaccines. Further analysis should also be focused on this aspect.
Footnotes
Author Disclosure Statement
No conflicting financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.