
Abstract
Influenza virus-like particles (VLPs) are effective vaccines against influenza infection, which can be produced either in insect cells by recombinant baculovirus (BV) infection or in mammalian cells by DNA plasmid transfection. However, VLPs produced from baculovirus/insect cells are difficult to purify due to baculovirus contamination; VLPs produced by plasmid transfection are limited by scale-up capability. In this study, a BacMam BV, in which three CMV-promoters drive the hemagglutinin, neuraminidase, and matrix of influenza virus was constructed. This baculovirus can deliver these genes into mammalian cells/hosts and subsequently influenza VLPs can be produced and secreted from transduced cells. Transduction conditions were optimized and influenza VLPs were purified from transduced 293T cells. Mice were vaccinated with BV transduction-produced VLPs, plasmid transfection-produced VLPs, and BacMam BV. Two vaccinations of each vaccine induced high hemagglutination-inhibition (HAI) titers and prevented influenza virus infection. In contrast, following a single vaccination, all mice vaccinated with each vaccine had significantly lower lung viral titers compared to unvaccinated mice. Remarkably, mice vaccinated with a single dose of BV transduction-produced VLPs survived challenge, whereas mice vaccinated with one dose of BacMam BV- or plasmid transfection-produced VLPs had 60–80% survival. This finding is particularly significant for producing easily purified VLPs. The BacMam system is an alternative strategy for VLP production, which is easy to scale up and purify. Besides, BacMam BV can be used as a gene delivery vector to produce VLPs in vivo, to stimulate immune responses.
Introduction
In 1983, Volkman et al. found that baculovirus infected 35 vertebrate cell lines, but without detectable baculovirus replication (11). Subsequently, mammalian cells transduced by baculovirus could efficiently express some proteins under the transcriptional control of mammalian promoters (12 –14). Many cell types can be transduced by baculovirus, including cell lines derived from human, non-human primate, rabbit, rodent, porcine, bovine, and fish hosts (15). In addition, baculovirus can transduce non-dividing cells (16) and primary cells in vivo (17). Recombinant baculoviruses containing mammalian expression cassettes are referred to as BacMam baculoviruses. Due to their high efficiency and low cytotoxicity during the gene delivery process, BacMam baculoviruses have been used as novel vectors for gene therapy and vaccine development (18 –24).
In this study, we constructed a BacMam baculovirus containing HA, NA, and M1 expression cassettes that were driven by cytomegalovirus (CMV) promoters. We speculated that BacMam baculoviruses would deliver these influenza genes into mammalian cells and result in the production of influenza VLPs. Since baculoviruses cannot replicate in mammalian cells, this strategy would not produce any baculovirus particles during influenza VLP production. After transduction of mammalian cells in vitro, we found that influenza VLPs can be easily generated and purified. In addition, since BacMam baculoviruses can transduce primary cells in vivo without cytotoxicity, these baculoviruses can potentially be used as a safe gene delivery vector to produce influenza VLP in vivo. Mice were immunized with purified VLPs produced from baculovirus-transduced mammalian cells, or directly immunized with this BacMam baculovirus. Both induced influenza-specific immune responses and provided mice sterilizing protection against influenza challenge.
Materials and Methods
Cells and viruses
Spodoptera frugiperda (Sf9) cells were propagated at 28°C in Sf-900II serum-free medium (Invitrogen, Carlsbad, CA). Cells were infected by recombinant baculoviruses at a multiplicity of infection (MOI) of 0.1–1.0, and virus supernatants were collected 4 d post-infection. The influenza virus used in this study was a mouse-adapted influenza virus, A/Puerto Rico/8/1934 (PR8, H1N1).
Plasmids and recombinant baculovirus construction
In order to construct multiple genes expressing recombinant baculoviruses, pFastBac Dual plasmid (Invitrogen) was modified to contain two CMV promoters (p2CMV) and two multiple cloning sites based on a vector we previously modified (25). CMV promoter and HSV tk/SV40 poly A signal sequences were amplified by overlapping PCR and inserted into the pFastBac vector backbone. At the same time two compatible restriction sites (Asc I and Mlu I) were introduced into p2CMV as shown in Fig. 1A and Supplementary Fig. 1 (see online supplementary material at
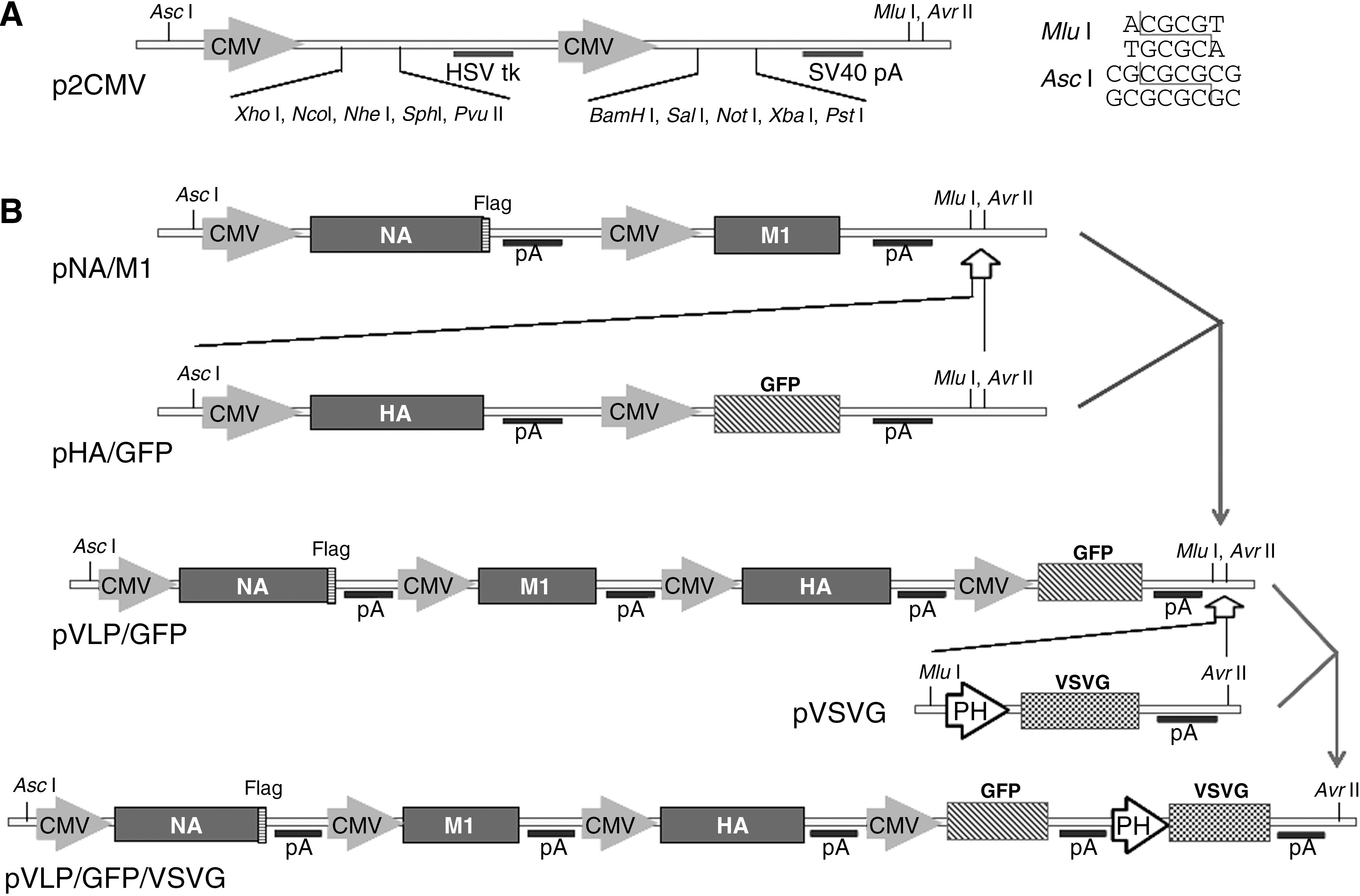
Construction of five-unit transfer vector that can express influenza HA, NA, and M1, as well as GFP and VSVG. (
Purification of recombinant baculoviruses
The recombinant baculoviruses were produced by infecting Sf9 cells at an MOI of 0.1. Supernatants were collected 4 d after infection and were clarified by centrifugation at 3000 g for 10 min at 4°C to remove cell debris. Viral particles were precipitated via ultracentrifugation (27,000 rpm using a Sorvall SW28 rotor) for 4 h at 4°C. The pellets were subsequently resuspended in phosphate-buffered saline (PBS) and stored at 4°C. The viral titer was determined using the BacPAK Baculovirus Rapid Titer Kit (Clontech, Mountain View, CA) and was expressed as infectious units per milliliter (ifu/mL).
Optimization of baculovirus transduction
To optimize the transduction efficiency, human embryonic kidney cells (293T) were seeded onto 6-well plates at a density of 5 × 105 cells/well in Dulbecco's modified Eagle medium (DMEM) supplemented with 10% fetal bovine serum (FBS) and incubated at 37°C overnight. Before transduction, the medium was removed and the cells were washed twice with PBS. The cells were transduced with BV-VLP/GFP or BV-VLP/GFP/VSVG at different MOI (20, 40, 60, 80, and 100) in 0.5 mL PBS. The cells were incubated at 28°C for a range of periods (1, 2, 4, 6, and 8 h). Following incubation, the viral inocula were removed, followed by two washes with PBS, and then 2 mL complete DMEM (cDMEM) with or without 5 mM sodium butyrate (Sigma-Aldrich, St. Louis, MO) were added. The transduced cells were cultured at 37°C for 72 h. To compare baculovirus transduction and plasmid transfection, 293T cells were seeded onto 6-well plates at a density of 1 × 106 cells/well in cDMEM and incubated at 37°C overnight. The cells were transfected with 1 μg plasmid pVLP/GFP and 5 μL lipofectamine 2000 in 0.5 mL OPTI-MEM (Invitrogen). After 5 h incubation at 37°C, the transfection medium was replaced with 2 mL cDMEM. The cells were cultured at 37°C for 72 h. The expression of GFP was monitored daily by fluorescence microscopy.
Western blot and hemagglutination assays for VLP production
Supernatants from transduced or transfected cells were harvested after 72 h of culture and were clarified by centrifugation at 3000 g for 10 min at 4°C to remove cell debris. The supernatants were subjected to Western blot analysis. The supernatants from BV-VLP transduced or pVLP-transfected 293T cells were examined for functional VLPs by hemagglutination assay. Briefly, 50 μL of serial twofold dilutions were prepared in PBS, followed by incubation with 50 μL of 1% turkey red blood cells (tRBCs; Lampire Biologicals, Pipersville, PA) at 25°C for 30 min. The extent of hemagglutination was inspected visually, and the highest dilution capable of agglutinating red blood cells was determined.
VLP production and purification
293T cells were transduced with BV-VLP/GFP/VSVG in PBS at an MOI of 60. The transduced cells were cultured in cDMEM containing 5 mM sodium butyrate for 72 h at 37°C. The VLP produced from baculovirus transduction was represented VLP-BV. Another VLP produced following transient transfection was used as a control. Briefly, 293T cells were transiently transfected with a plasmid (pVLP) encoding M1, NA, and HA of PR8 virus, and incubated for 72 h at 37°C. Culture supernatants (200 mL) from transduced or transfected cells were collected and were clarified by centrifugation at 3000 g for 10 min at 4°C to remove cell debris. VLPs were precipitated via ultracentrifugation as described above. The pellets were resuspended in 1 mL PBS and loaded onto 20–60% sucrose gradients, and sedimented by ultracentrifugation for 18 h at 27,000 rpm at 4°C. Fractions containing influenza VLPs were collected and diluted in PBS followed by ultracentrifugation again. The pellets were resuspended in PBS and stored at 4°C. Baculovirus residues were checked with the BacPAK Baculovirus Rapid Titer Kit. Protein concentration was determined by bicinchoninic acid (BCA) assay (Thermo Scientific, Rockford, IL). All VLPs used in this study were based on total protein. The hemagglutination titer of VLPs was examined with an HA assay.
Mouse immunization and challenge
Female BALB/c mice (6–8 wk old) were purchased from Harlan Sprague Dawley (Indianapolis, IN). The mice (8 groups, 10 mice per group) were vaccinated: (1) once or twice with 3 μg VLP-BV representing VLPs derived from BV-VLP/GFP/VSVG transduction; (2) once or twice with 3 μg VLP-tsf representing VLP derived from pVLP transfection; (3) once or twice with 1 × 108 ifu BV-VLP representing purified BV-VLP/GFP/VSVG; (4) twice with 1 × 108 ifu BV representing purified wild-type baculovirus; and (5) with PBS. All vaccines were administered intramuscularly to the mice without any adjuvants.
For virus challenge, ketamine-anesthetized mice were intranasally infected with 1500 pfu of A/PR/8/1934 virus (equivalent to 10 × the 50% lethal dose [LD50]) in 50 μL of PBS at 3 wk after the final immunization. For measurement of virus replication in the lungs, five mice from each group were sacrificed on day 3 post-challenge. The other five mice were monitored daily for clinical signs of influenza infection (i.e., weight loss, ruffling fur, or inactivity) and body weight was recorded each day. Mice that lost greater than 20% of body weight were euthanized.
Hemagglutination inhibition (HAI) titer in sera and viral titer in lungs
Blood samples were collected from anesthetized mice via retro-orbital plexus puncture at 2 wk after each immunization. After the blood samples were clotted and centrifuged, serum samples were collected. The HAI assay was used to assess the functional antibodies to HA able to inhibit agglutination of erythrocytes as previously described (3).
Three days after challenge, five mice from each group were euthanized. The lungs were harvested and were homogenized by passing them through a cell strainer (BD Biosciences, Bedford, MA) in 1 mL of PBS. The homogenates were centrifuged at 1000 × g for 10 min. The lung supernatants were collected for plaque assay on Madin-Darby canine kidney (MDCK) cells as previously described (3).
Results
Optimization of transduction conditions
Previous studies reported that many factors affect the efficiency of baculovirus transduction, including cell lines, transduction medium, incubation time, MOI, VSVG, and sodium butyrate (26
–28). Initially, we investigated the protein expression after baculovirus transduction in several cell lines, such as Vero, HeLa, MDCK, HEK293, and 293T, and found that 293T is the most susceptible cell line and yields the highest protein expression (data not shown). As previously reported, PBS had the highest transduction efficiency as the transduction medium (26). Two hours is long enough for baculovirus transduction into 293T cells. Therefore, transduction was conducted in PBS for 2 h followed by two washes with PBS or DMEM. Then, the effect of MOI was investigated by transducing 293T cells with BV-VLP/GFP at different MOIs. Following 72 h of incubation, supernatants were harvested and NA expression was determined by Western blot (Fig. 2A). The intensity of each band in the Western blot was analyzed with ImageJ software (
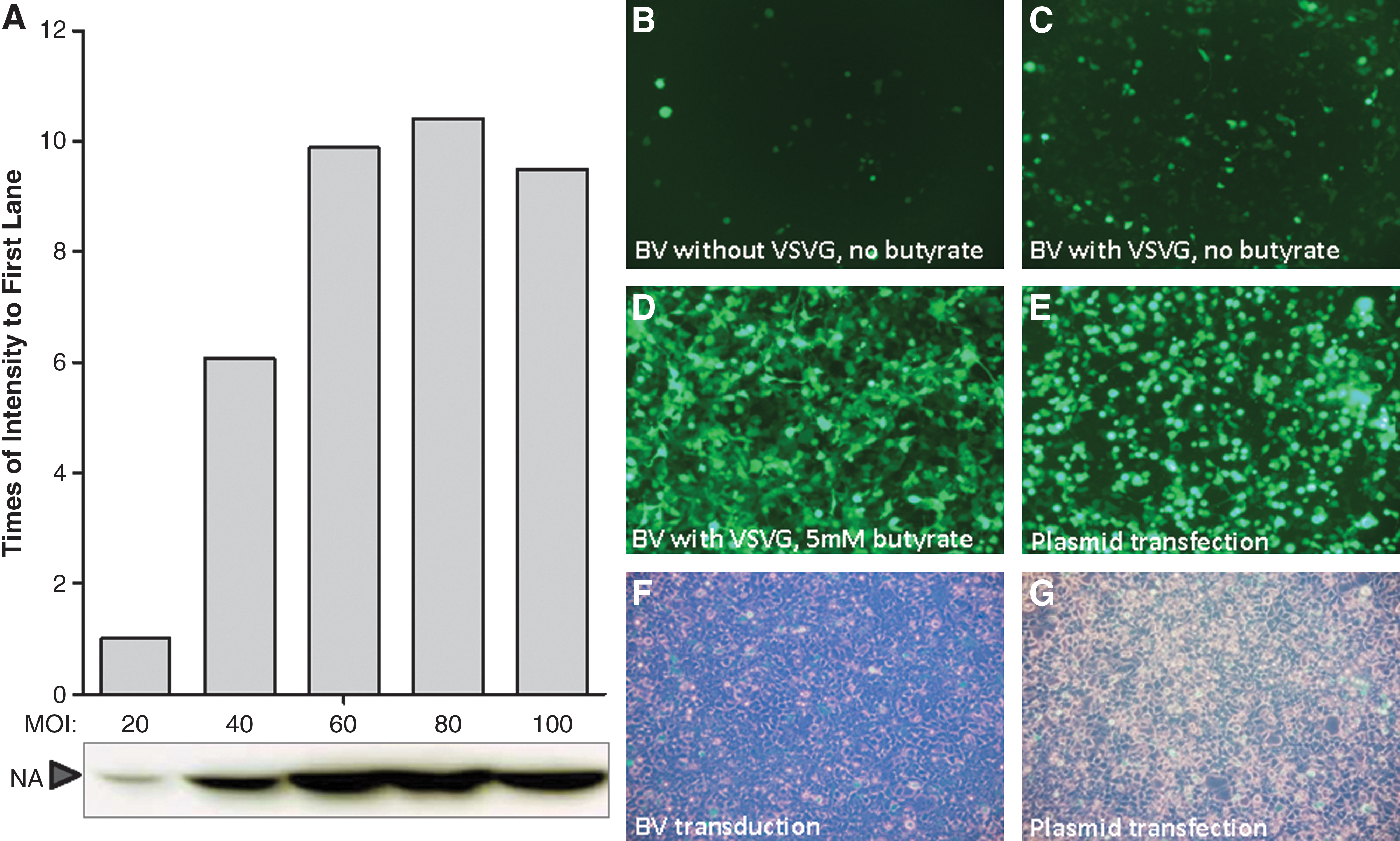
Optimization of BV transduction conditions. (
Previous studies demonstrated that VSVG can affect baculovirus transduction both in vitro and in vivo (30), and sodium butyrate can enhance gene expression in baculovirus-transduced cells (28). Therefore, the effects of VSVG and butyrate on baculovirus transduction of 293T cells were examined. At MOI of 60, 293T cells were transduced with BV-VLP/GFP or BV-VLP/GFP/VSVG. After transduction, the cells were cultured with or without sodium butyrate. Under the same transduction and culture conditions, the cells transduced with BV-VLP/GFP/VSVG showed a higher level of fluorescence intensity than with BV-VLP/GFP, indicating that these cells were more efficiently transduced (Fig. 2B and C) (30). Furthermore, the addition of 5 mM sodium butyrate enhanced GFP expression (Fig. 2C and D). In addition, plasmid transfection results in a high level of GFP expression (Fig. 2E), but the cells transfected with DNA plasmid had more dead/dying cells compared to cells transduced with baculovirus (Fig. 2F and G) (31).
Influenza VLP production
To investigate whether the HA, NA, and M1 can be expressed simultaneously in baculovirus-transduced or plasmid-transfected mammalian cells, 293T cells were transduced with BV-VLP/GFP/VSVG at an MOI of 60, or transfected with pVLP/GFP. As shown in Fig. 3A, three bands corresponding to the molecular weights of HA, NA, and M1 were detected in the supernatants of BV-VLP/GFP/VSVG-transduced and pVLP/GFP-transfected cells, but not in the supernatant of wild-type BV-transduced cells.
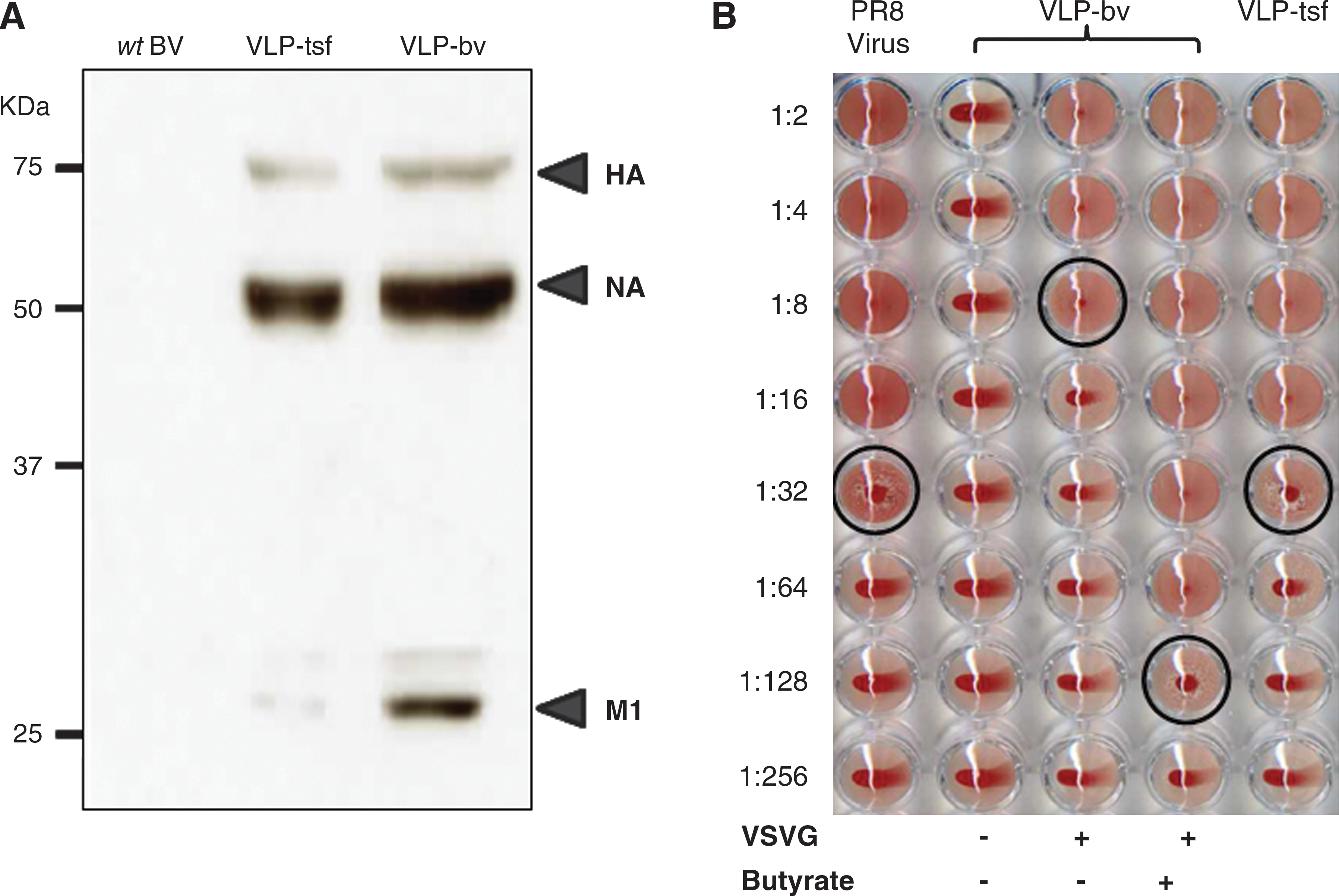
VLP production from BV transduction or plasmid transfection. (
HA on viral particles or VLPs has the ability to agglutinate red blood cells, while soluble HA cannot agglutinate RBCs, and the hemagglutination titer can reflect the concentration of viral particles (32). Therefore, the functional VLPs can be examined by hemagglutination assay. The supernatants from 293T cells transduced with BV-VLP without VSVG had no detectable HA titer (Fig. 3B). However, the addition of VSVG in BacMam BV increased the HA titer to 1:8. The addition of 5 mM sodium butyrate to the culture medium increased the HA titer to 1:128, which was 4 times higher than 293T cells directly transfected with plasmid pVLP/EGFP. In addition, following sucrose gradient purification, there was no detectable baculovirus in the purified VLP products by plaque assay, although baculovirus DNA could be detected by PCR (data not shown).
Analysis of the immune response
The immunogenicity of purified VLPs and BacMam baculovirus were examined in BALB/c mice by intramuscular injection at week 0 and/or 3. Serum samples were collected 2 wk after each vaccination (weeks 2 and 5), and evaluated for the ability to inhibit influenza virus-induced hemagglutination of tRBCs (HAI), which is a main indicator for the HA-specific immune response. The seroprotection rate (defined as the percentage of subjects with an HAI titer ≥1:40) is a well-accepted criterion for the evaluation of influenza vaccine efficacy. A seroprotection rate >70% is one requirement for an influenza vaccine, according to the EU Committee for Human Medicinal Products (CHMP) (33).
After one immunization with VLP-BV, 80% of mice had detectable HAI titers (≥1:10) against PR8 virus; 35% of mice had an HAI titer ≥1:40. After one immunization with VLP-tsf, 35% of mice had detectable HAI titers, while none had an HAI titer ≥1:40. After one immunization with BV-VLP, 50% of mice had detectable HAI titers, while 10% of mice had an HAI titer ≥1:40. After the second vaccination, a significant rise in HAI titers of all immunized mice could be observed. The average HAI titers of mice vaccinated with VLP-BV, VLP-tsf, and BV-VLP were 1:240, 1:100, and 1:112, respectively (Fig. 4). The seroprotection rates for all three vaccines ranged between 80 and 100%, and met the CHMP requirement for immunogenicity. Remarkably, mice vaccinated with VLP-BV had significantly higher HAI titers compared to mice vaccinated with VLP-tsf, after both priming and boosting immunization. As expected, all mice vaccinated with wt BV and mock vaccinated had no detectable HAI titers.
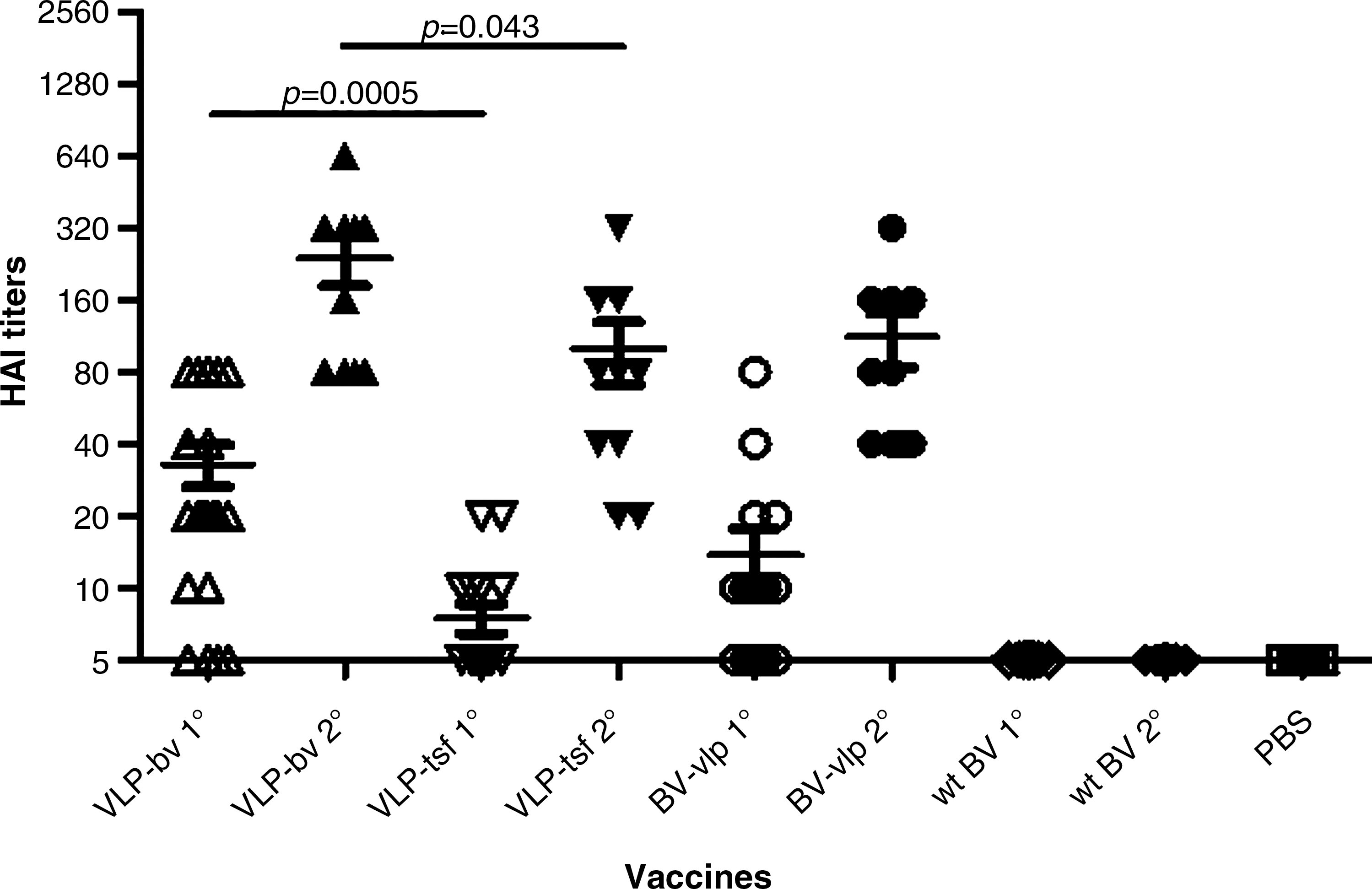
Hemagglutination-inhibition (HAI) titers. Mice were immunized once (1°) or twice (2°) intramuscularly with VLP derived from BV transduction (VLP-BV), VLP derived from plasmid transfection (VLP-tsf ), BV producing PR8 VLP (BV-VLP), wild-type BV (wt BV), and PBS at a 3-week interval (10 mice/group). Blood samples were taken 2 wk after each vaccination. Serum HAI antibody titers were assessed against PR8 virus. Bars indicate geometric mean titer (GMT) ± SEM.
Protection from lethal viral challenge
To evaluate the protective efficacy of each vaccine, all vaccinated mice were challenged intranasally with a lethal dose of mouse-adapted PR8 virus. Lung viral titers were determined at day 3 post-challenge. Mice vaccinated with wt BV or PBS had high viral lung titers (∼1 × 106 pfu/mL), while the mice that received only one vaccination with VLP-BV, VLP-tsf, or BV-VLP had significantly lower viral lung titers (2 × 104 to 2 × 105 pfu/mL) (Fig. 5). There were no significant differences between the three different vaccines. All mice that received two vaccinations had undetectable viral titers (<10 pfu/mL) in their lungs.
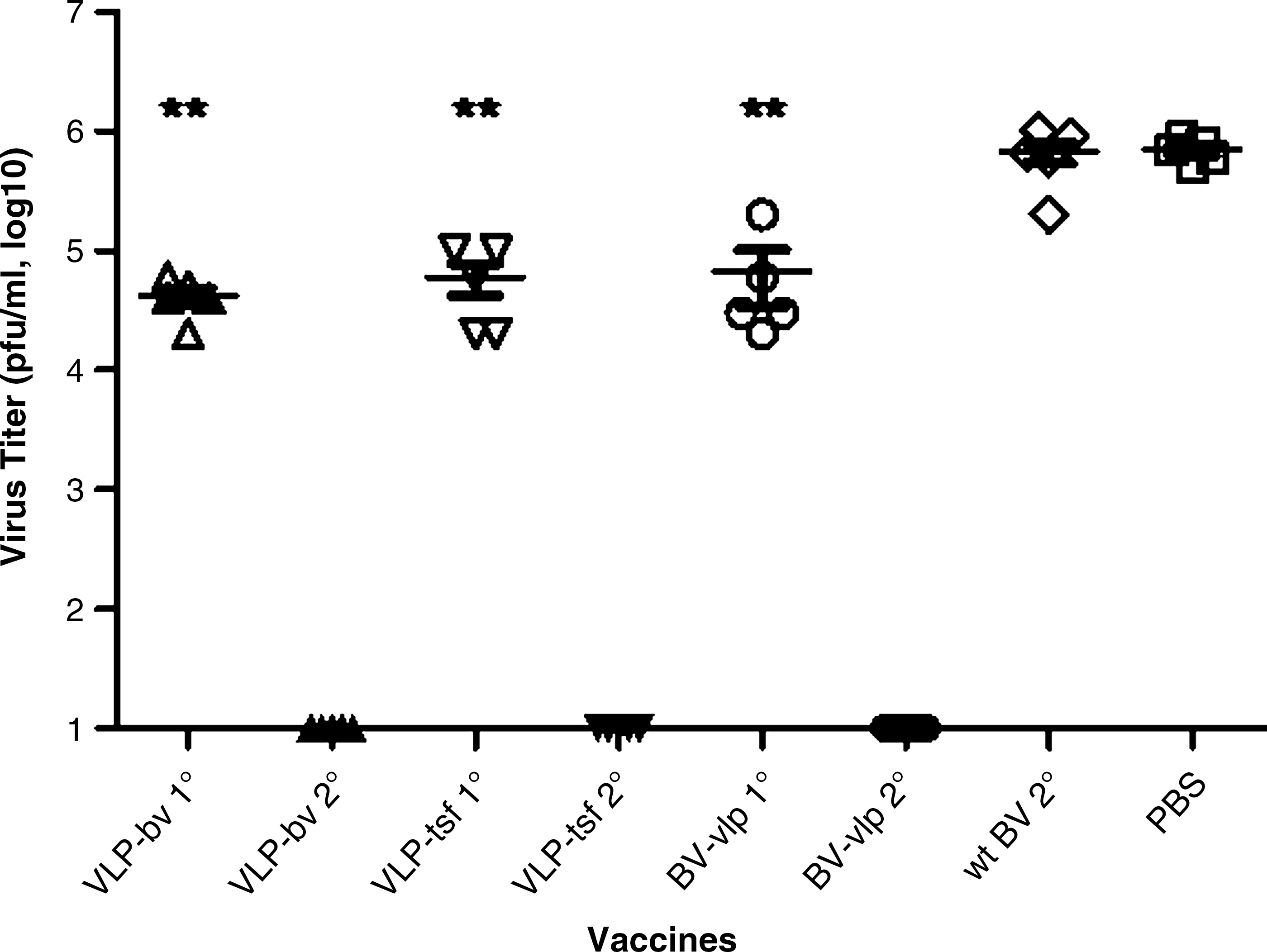
Virus titers in lungs at day 3 post-challenge. Mice were immunized once (1°) or twice (2°) intramuscularly with VLP-BV, VLP-tsf, BV-VLP, wt BV, and PBS. At week 3 after the last immunization, the immunized mice were intranasally infected with a lethal dose of mouse-adapted PR8 virus (10 LD50). Lung samples from individual mice in each group (five mice per group) were collected on day 3 post-challenge, and each sample was diluted in 1 mL PBS. The titers are presented as log10 pfu per mL (**p < 0.01 compared with the wt BV 2° and PBS groups).
All mice vaccinated with wt BV or PBS had signs of morbidity (e.g., ruffling fur, shivering, and inactivity), and had lost greater than 20% of their original body weight by days 8–9 post-challenge (Fig. 6). All mice vaccinated once with VLP-BV were protected with an average of 7% weight loss (open triangles in Fig. 6). Eighty percent of mice vaccinated once with VLP-tsf were protected against influenza challenge, and 60% of mice vaccinated with BV-VLP were protected, even though some mice showed signs of sickness. However, all mice survived without any signs of sickness or weight loss following two vaccinations with VLP-BV, VLP-tsf, or BV-VLP.
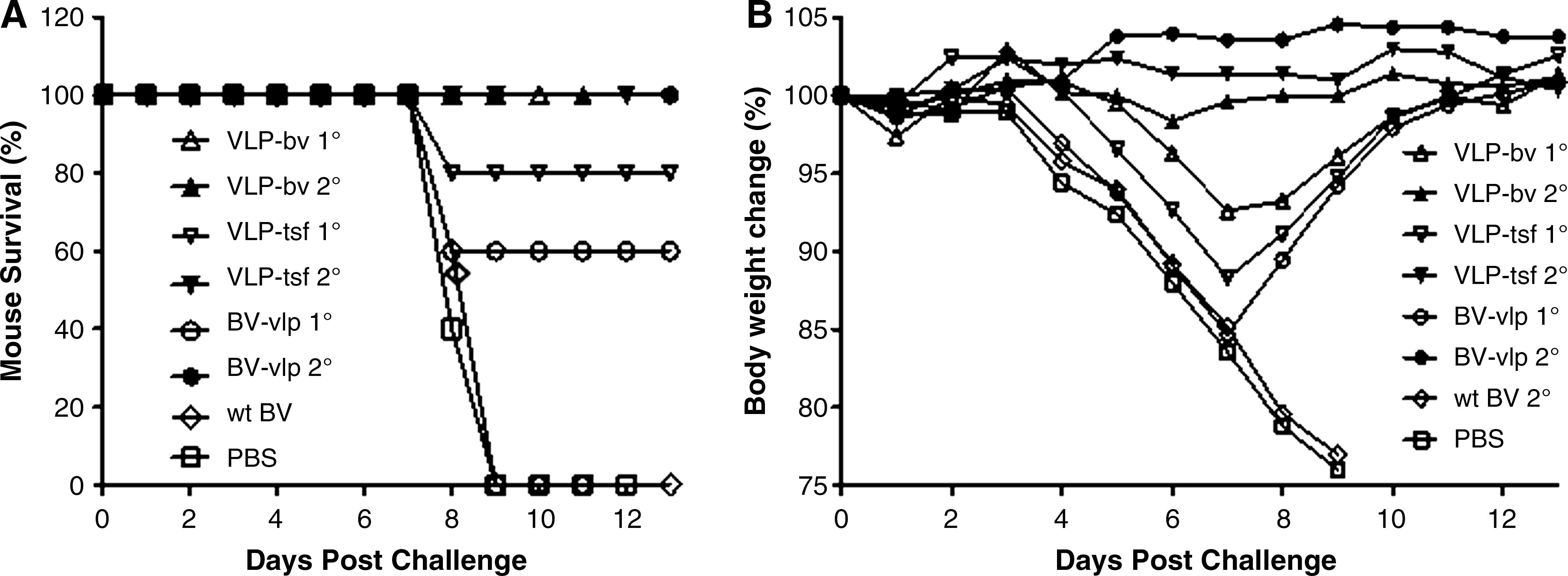
Protection of mice from lethal PR8 challenge. Mice were immunized once (1°) or twice (2°) intramuscularly with VLP-BV, VLP-tsf, BV-VLP, wt BV, and PBS. At week 3 after the final immunization, the mice were intranasally infected with a lethal dose of mouse-adapted PR8 virus (10 LD50) (five mice per group). The mice were monitored daily for 13 d. (
Discussion
VLPs mimic the overall structure of parental virus particles without packaging infectious genetic material (34). It is a promising approach to the production of vaccines due to its low risk and high immunogenicity (35). VLP vaccines have been produced from yeast-, insect-, mammalian-, and plant-based systems for a variety of viral pathogens (5). VLPs produced from mammalian cells have several advantages, such as ease of purification and similar post-translational modifications and cytoplasmic trafficking processes as wild-type viruses. However, mammalian-based production systems are hampered by high costs and difficulties with production scale-up. The use of yeast and insect expression systems are inexpensive and relatively easy to scale-up, but the VLP purification is always a challenge because VLPs produced in yeast cells are usually not secreted, and VLPs produced in insect cells are accompanied by baculovirus particles (10,36). Baculoviruses are rod-shaped particles roughly 70 nm × 320 nm in size (37,38), which are similar in size to influenza virions. Many methods have been developed to improve the purification of VLPs derived from baculovirus/insect-cell systems, including density-gradient ultracentrifugation, size-exclusion chromatography, and affinity chromatography. These methods are efficiently purifying some small, non-enveloped VLPs, such as norovirus (38 nm) and human papillomavirus (40–50 nm) (5,39), but they are not efficient for other larger VLPs, such as HIV (100–120 nm) or influenza VLPs (80–120 nm) (6,40 –42).
In this study, we developed a new strategy to produce influenza VLPs in mammalian cells with baculovirus vector. BacMam baculoviruses can be produced using a standard method in insect cells. Gene expression in transduced cells can last over 16 days (43). Due to its rapid and facile gene expression in various mammalian cells, the BacMam system has been used as a substitute for direct plasmid transfection (44). Influenza VLPs can be produced by simultaneously expressing HA, NA, and M1 in insect/mammalian cells. To ensure gene uptake and expression in target cells, we developed a new BacMam baculovirus carrying all information necessary to induce influenza VLP production. The resulting constructs, BV-VLP/EGFP and BV-VLP/EGFP/VSVG, allowed for the production of influenza VLPs. Transduction efficiency was optimized and VLP production was characterized. Since baculovirus cannot replicate in mammalian cells as it does in insect cells (11), the purification of VLPs produced from the BacMam system is much easier than VLPs produced from insect cells. The BacMam system is also easy to scale up by using bioreactors such as the baculovirus/insect-cell expression system (44). Therefore, the BacMam system combines the advantages of both baculovirus/insect cell expression and DNA plasmid transfection methods: it is inexpensive, rapid, and facile for experiments for gene overexpression, and is easy to scale up and purify. Of course, there are also some limitations, such as repetitious washing of transduced cells that would be labor-intensive for suspension culture; two-tiered banking systems for both mammalian and insect cells are needed to implement this strategy.
Vaccination of mice with in vitro-produced influenza VLPs or with BacMam BV induced equally high antibody responses following two vaccinations (HAI titers 1:20–1:640). Remarkably, VLPs produced via BV transduction induced significantly higher HAI titers than VLPs produced via plasmid transfection (Fig. 4). This effect may be because VLPs produced via plasmid transfection contain a higher content of cellular proteins. During direct transfection of culture cells with plasmids, the transfection reagents are potentially cytotoxic, which may result in a high degree of cell death and subsequent cell debris contamination of the VLP preparation. However, the BacMam BV transduction process is not associated with cytotoxicity, even at high MOI (>500) (31). At equal protein concentrations, the VLPs produced via BV transduction have 2–4 times higher HA titers than VLPs produced via plasmid transfection, suggesting a higher portion of influenza VLPs with functional HA on the particles compared to VLPs produced via plasmid transfection. An additional reason could be that baculovirus DNA may remain in the VLP products derived from the BacMam system, which could serve as adjuvant for this vaccine. Baculovirus has strong adjuvant effects even at low concentrations (103 pfu), and baculoviral DNA, but not protein, has adjuvant activity (45). We found that baculovirus DNA was detectable by PCR from the purified VLPs, although no baculovirus could be detected by plaque assay (data not shown).
To evaluate the protection induced by VLP vaccines, all vaccinated mice were challenged with a homogeneous, mouse-adapted influenza virus (A/PR/8/1934), administered at a lethal dose. The viral clearance in lungs provides a sensitive indicator for assessing protective efficacy. At 3 d post-challenge, all mice in the control groups showed high lung viral titers. Mice vaccinated two times with VLP-BV, VLP-tsf, and BV-VLP had no detectable virus post-challenge. Mice receiving one vaccination had two- to 50-fold reductions in viral lung titers compared to unvaccinated or wt BV vaccinated mice (Fig. 5). These results indicate that for all three vaccines, two vaccinations can induce strong immune responses that are sufficient to prevent infection. In contrast, one vaccination reduces viral replication, but does not prevent infection. As for the protection seen after one vaccination, the results indicated that one vaccination with VLP-BV provided 100% protection, while one vaccination with VLP-tsf or BV-VLP conferred 80% and 60% protection, respectively. Although BV-VLP is a little less effective in protection compared to other VLPs, it is still a promising vaccine strategy because of its ease of scale-up. Since BV cannot infect mammals and does not cause cytopathic effects (11,18), safety issues are greatly reduced. Of course, more studies are needed to address its safety. Better protection should be achievable by giving higher doses. Recently, it was demonstrated that high doses of BV delivering influenza HA protected chickens against avian influenza virus challenge, while a low dose did not elicit protection (21). Therefore, protection with a single vaccination would be expected if a higher dose of VLP-tsf or BV-VLP is administered.
When considered together with previous data, the results of our current studies suggest that BacMam system-derived influenza VLPs represent a promising and safe vaccine candidate for protection against influenza infections. Due to its low cytotoxicity and high gene delivery efficiency, the BacMam baculovirus represents a significant advance, and it is a promising alternative vaccine strategy. The major drawback of directly using BV as a gene delivery vector is that it can induce immune responses against the BV vector, which will limit the repeat use of this vector. Future studies to evaluate the potential benefits of using this strategy in controlling other infectious diseases are needed.
Footnotes
Acknowledgments
This project was funded in part by a grant from the Pennsylvania Department of Health. The Department specifically disclaims responsibility for any analyses, interpretations, or conclusions. We thank Dr. Jared D. Evans for helpful suggestions.
Author Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.