
Abstract
Bovine herpesvirus-1 (BoHV-1) infection is distributed worldwide and the development of new tools to fight against this pathogen has become extremely important. In this work a recombinant modified vaccinia virus Ankara (MVA) vector expressing the secreted version of glycoprotein D, MVA-gDs, was obtained and evaluated as a candidate vaccine. First, the correct expression, antigenicity, and N-glycosylation of glycoprotein D were confirmed by molecular techniques. Then MVA-gDs was used as parenteral immunogen in BALB/C mice in which a specific anti-gD humoral immune response was induced and maintained for 7 mo. Two doses of MVA-gDs supplemented with cholera toxin delivered by intranasal immunization induced IgA anti-gD humoral immune responses in nasal and bronchopulmonary washes, as well as IgG anti-gD antibodies in serum samples. In order to evaluate the protection conferred by MVA-gDs immunization, a rabbit BoHV-1 challenge assay was performed. A shorter viral excretion period and a reduction in the number of animals shedding BoHV-1 was observed in the group immunized with recombinant MVA-gDs. In conclusion our data encourage further studies to evaluate MVA-gDs, alone or combined with other immunogens, as a candidate vaccine for BoHV-1.
Introduction
Modified vaccinia virus Ankara (MVA) is a highly attenuated virus that has been used as a safe and efficient vaccine vector (17). In mammalian cells, MVA morphogenesis is impaired, allowing immunization to occur in the absence of viral replication, and eliminating the chances of vector dissemination between vaccinated animals, and consequently to unvaccinated ones or to the environment. Importantly, upon intranasal MVA inoculation inflammatory reactions have not been detected in the central nervous system or in the upper and lower airways (7,14,23). Therefore, non-replicative recombinant MVA vectors have gained increasing interest in the development of new veterinary vaccines (2,5,27).
Since BoHV-1 infection is widely distributed, the purpose of the present study was to develop a MVA-based vaccine able to express the secreted version of glycoprotein D of BoHV-1 in vivo. First, recombinant MVA-gDs virus was obtained and molecularly characterized. Then recombinant MVA-gDs immunogenicity and efficacy were assessed in mice and rabbits, respectively.
Materials and Methods
Cells
Chicken embryo fibroblasts (CEFs) were prepared from 11-day-old specific pathogen-free (SPF) embryos (Instituto Rosenbusch, Buenos Aires, Argentina) and maintained in 199 Earle medium supplemented with 2.95 mg/mL tryptose phosphate broth (BD, Sparks, MD), 2.2 mg/mL sodium bicarbonate (ICN Biomedicals Inc., Irvine, CA), 0.3 mg/mL L-glutamine, 50 μg/mL gentamicin, 66 μg/mL streptomycin, 100 U/mL penicillin, and 10% fetal calf serum (FCS; Internegocios, Buenos Aires, Argentina). Madin Darby bovine kidney (MDBK) cells (American Type Culture Collection [ATCC]) were maintained in Eagle minimal essential medium (E-MEM) supplemented with 100 μg/mL streptomycin, 100 U/mL penicillin, 0.25 μg/mL amphotericin B, and 2% FCS. Baby hamster kidney (BHK-21) cells (ATCC) were maintained in Dulbecco's modified medium supplemented with 2% FCS.
Construction and characterization of recombinant MVA-gDs virus
In order to obtain recombinant MVA viruses we constructed a transference vector (VT-MTK-gDsGus) carrying selected genes as well as viral genomic regions of the MVA086R gene (GeneBank Accession U94848) to allow in vivo recombination. The selected genes were the coding sequences of the secreted version of glycoprotein D (gDs) of BoHV-1, and the uid A (coding the β-glucuronidase enzyme [GUS]), under regulation of poxviral synthetic pE/L or H6 promoters, respectively. These sequences were excised from plasmid available in our laboratory, and they were subcloned sequentially into VT-MTK plasmid as described previously (10). Recombinant MVA viruses expressing gDs were made by transfecting VT-MTK-gDsGus into CEFs infected previously with MVA at a multiplicity of infection (moi) of 0.25 PFU per cell. The expression of GUS allowed screening and plaque purification on CEFs in the presence of β-glucuronidase substrate (X-Gluc, 5-bromo-4-chloro-3-indolyl-β-D-glucuronic acid; Inalco, Milan, Italy) (6). Viral stocks were prepared in CEFs and purified by ultracentrifugation through 25% v/v sucrose cushion prepared in TMN buffer (10 mM Tris-HCl, pH 7.5, 1.5 mM MgCl2, and 10 mM NaCl) at 154,000 × g for 2 h in SW41Ti rotor (Beckman Coulter, Brea, CA). In order to confirm genetic stability of recombinant MVA-gDs, CEFs monolayers were infected at a low moi (0.05–0.1), and after 48–72 h cells were collected, frozen, thawed, and used in a new round of infection at a low moi. This procedure was repeated 10 times. Then PCR and Western blot analysis were performed to evaluate the absence of wild-type DNA, and to confirm the expression of gD, respectively.
PCR characterization of recombinant MVA-gDs viruses
Total DNA was extracted with extraction buffer 2 × (20 mM Tris, pH 8, 20 mM EDTA, 2% SDS, 4% β-mercaptoethanol, and 1 mg/mL proteinase K) from MVA- or MVA-gDs-infected CEFs at a moi of 1. In the “gDs PCR,” the presence of the gDs coding sequence was evidenced by the amplification of a 1.1 kbp fragment using HBV1sig: 5′AAGAATTCGGCTGCTGCGAGCGGGCCGAACA and HBV1sol: 5′AAGAATTCTCAGGCGTCGGGGGCCGCGGGCGTA primers (36). To confirm the purity of the recombinant viruses, a PCR reaction was performed to simultaneously amplify wild-type and/or recombinant genome (Fig. 1A). The primers used were: TK1: 5′TCCCCGCGGTGAACGGCGGACATATTC; TK4: 5′GGGGTACCTTATGAGCCGACGTAACA (2 × concentrated), and HBV1sig.
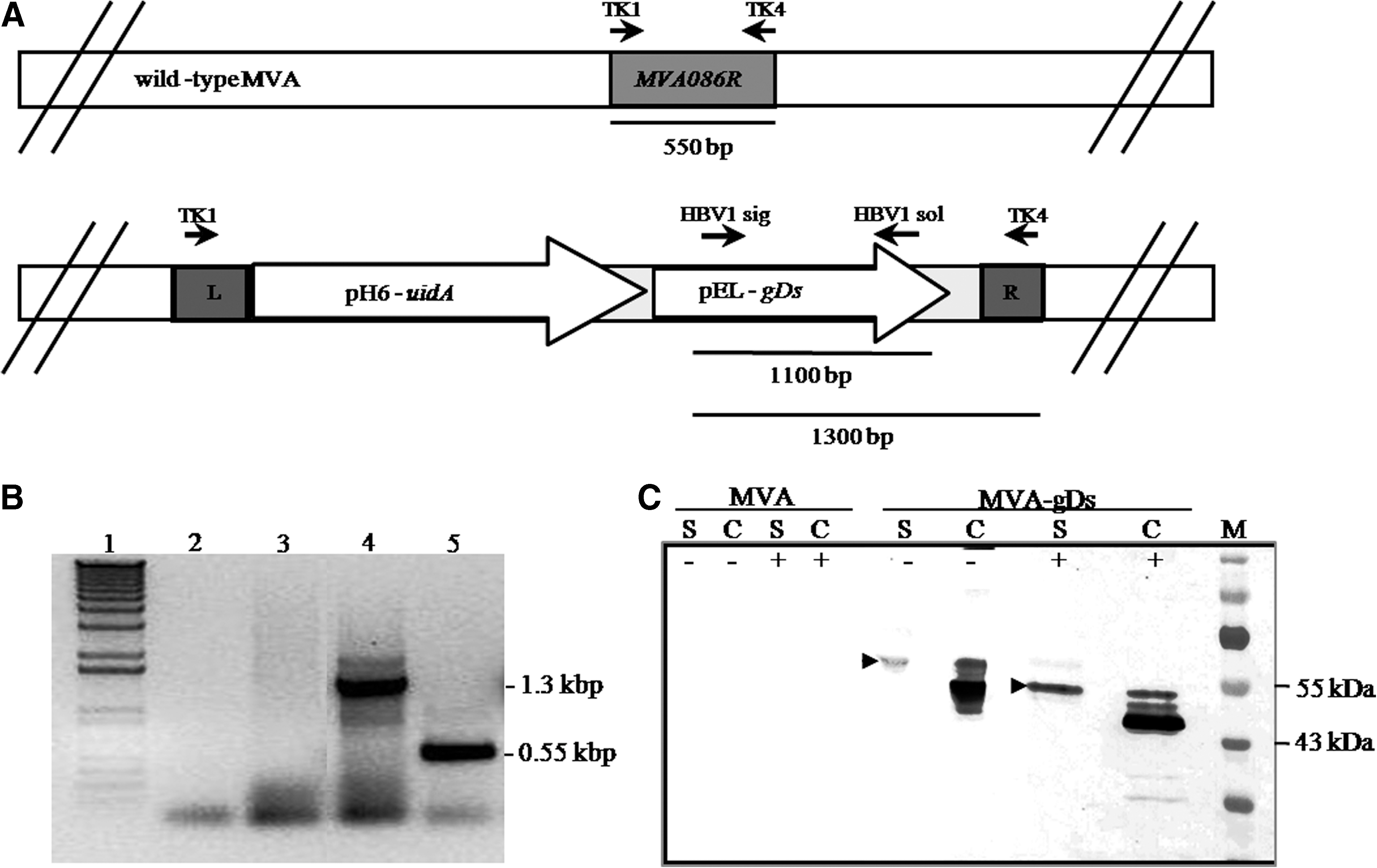
Characterization of MVA-gDs. (
PnGase F treatment
Supernatants and cell extracts of uninfected, MVA-, or MVA-gDs-infected CEFs were treated or mock treated with PnGaseF enzyme (New England BioLabs, Inc., Beverly, MA) according to the manufacturer's instructions and analyzed by Western blot as described below.
Western blot assays
Protein extracts were loaded onto polyacrylamide gel (10% PAGE-SDS) and, after electrophoresis, proteins were transferred onto nitrocellulose membranes. The expression of gD was assessed by Western blot using a mouse polyclonal anti-BoHV-1 serum and an alkaline phosphatase-conjugated anti-mouse-specific IgG (Sigma-Aldrich Inc., St. Louis, MO). Recombinant gD was revealed by BCIP/NBT substrate precipitation (Promega, Madison, WI). In the BoHV-1 challenge rabbit assay, the presence of specific anti-BoHV-1 antibodies in serum was assessed by Western blot using BoHV-1 sucrose gradient-purified virus as antigen, and an alkaline phosphatase-conjugated anti-rabbit-specific IgG (Sigma-Aldrich), followed by BCIP/NBT substrate precipitation.
Anti-gD antibody ELISA
Detection of specific anti-gD antibodies was assessed by a modified ELISA as described previously (36). Anti-mouse IgG + IgM peroxidase-conjugated (Axell, Westbury, NY), or anti-rabbit IgG peroxidase-conjugated (Sigma-Aldrich) antibodies were used for humoral immune response detection. For the mucosal response, anti-mouse IgA peroxidase-conjugated (Southern Biotechnology, Birmingham, AL) antibody was employed. The reaction was developed by addition of 0.4 mg/mL ABTS (2-2′-azino-bis-3-ethylbenzthiazoline-6-sulphonic acid; ICN Biochemicals) and 0.0015% H2O2 in 50 mM citric acid buffer (pH 5), and read at 405 nm in a Multiskan spectrophotometer (Labsystems, Basingstoke Hants, U.K.). Anti-gD Ig ELISA titers were calculated as the highest dilution of serum from MVA-gDs-immunized animals that yielded an optical density value (OD) of at least twice as high as the corresponding OD from MVA-vaccinated animal sera.
In vitro BoHV-1 neutralization assays
Neutralizing antibodies against BoHV-1 were tested in MDBK cells in 96-well microtiter plates using the constant virus-variable serum method. Briefly, serum samples were inactivated for 30 min at 56°C and two-fold serial dilutions were prepared. Serum samples were incubated for 1 h at 37°C with 200 TCID50/mL of BoHV-1 (Los Angeles strain). Each dilution was tested in four wells. Then the serum-virus mixture was transferred to the cell monolayer and incubated for 1 h at 37°C and 5% CO2. Once the incubation period was finished, the supernatant was removed and E-MEM medium supplemented with 2% FCS was added. Cell plates were incubated for 72 h at 37°C and 5% CO2, and the cytopathic effect in the different wells was scored. Results were calculated by the Reed and Muench method (24), and the reciprocal value of the highest serum dilution in which a cytopathic effect was prevented was considered as the seroneutralizing antibody titer.
Mice immunization protocols, sample collection and processing
SPF female BALB/c mice, 8–10 wk old, were purchased from the Animal Services Laboratory, Faculty of Veterinary Medicine, University of La Plata (Buenos Aires, Argentina), and then housed in animal facilities at the Biotechnology Institute, INTA. All experiments were done in compliance with international legal and institutional guidelines. For systemic immune response assays, groups of four mice were immunized intraperitoneally (IP) with purified MVA or MVA-gDs viruses. Each dose consisted of 2 × 107 PFU/200 μL, and the second dose was administered 21 d later. As controls, animals were inoculated IP twice (days 0 and 21) with sucrose discontinuous gradient (30–50%) purified BoHV-1 (Los Angeles strain). The viral purification was performed as described previously (20). Each dose consisted of 1 × 106 TCID50 formulated 1:1 in incomplete oil adjuvant (200 μL final volume). For mucosal immune response assays, the mice were anesthetized IP with one dose of 50 mg/kg ketamine (Holliday-Scott, Buenos Aires, Argentina) and 2.5 mg/kg xylazine. Then immunizations with purified MVA or MVA-gDs viruses were given intranasally (IN) in 50 μL of TMN buffer. Each dose consisted of 1.5 × 107 PFU and 10 μg of cholera toxin. Two groups of animals (6 mice/group) received only one dose, and two other groups received a second dose 14 d later. At 7, 14, and 21 d after receiving the last immunization the mice were euthanized, the trachea was exposed, and nasal and bronchopulmonary washes were performed with a catheter by injecting 500 μL of PBS containing 20 μg/mL of aprotinin (Sigma-Aldrich). The recovered fluids were centrifuged at 3000 × g for 10 min at 4°C to remove cellular debris and kept at −20°C until used for ELISA tests. In all cases, serum samples were taken at different time points and assayed for the presence of anti-gD antibodies by ELISA.
Rabbit immunization protocols, sample collection and processing
New Zealand male and female rabbits, 6–8 wk old, were purchased from a breeding farm located in La Plata, Buenos Aires, Argentina, and housed in the animal facilities belonging to the Virology Laboratory of the Faculty of Veterinary Medicine (University of La Plata, Buenos Aires, Argentina). All experiments were done in compliance with international legal and institutional guidelines. The first experiment consisted of two groups (two animals each). Two doses of MVA or MVA-gDs virus were administered intramuscularly (IM) (1 mL containing 1 × 107 PFU) 15 d apart. Serum samples were obtained at 0, 15, 18, 21, 24, and 27 days post-immunization (dpi), and the presence of anti-gD antibodies was evaluated by ELISA. The second experiment consisted of three groups (four animals each) named MVA, MVA-gDs, or unvaccinated. Animals from the MVA and MVA-gDs groups were IM vaccinated with an initial virus dose (1 × 107 PFU), and received a booster dose on day 17. Nine days after the second immunization (26 dpi), all groups were IN challenged with 5 × 105 TCID50 of BoHV-1 (Los Angeles strain) using a protocol described previously (30). This method was selected because it mimics the natural route of BoHV-1 infection. Serum samples were taken from all groups at the beginning of the experiment (0 dpi), after the first dose of immunogen (13 dpi), the day of challenge (26 dpi), and 2 wk after challenge (40 dpi). Serum samples were assayed for the presence of anti-gD or anti-BoHV-1 antibodies by ELISA and Western blot, respectively. After BoHV-1 challenge, nasal swabs were collected daily and processed individually using standard methods for viral isolation. The amount of isolated virus was determined by infectivity titration with 10-fold serial dilutions of each swab over MDBK cells using the Reed and Muench method (24). The titer was expressed as log TCID50/mL. Three blind passages in MDBK monolayers were performed with negative samples for BoHV-1 isolation. Also, respiratory clinical signs (serous or mucus rhinitis) were recorded during the whole experiment.
Statistical analysis
Statistical differences between experimental groups were analyzed using the appropriate non-parametric test included in GraphPad Prism 5 software. A Mann-Whitney U test (two-tailed), Friedman test, and Dunn's multiple comparison test were used. p Values <0.05 were considered statistically significant.
Results
Construction and in vitro characterization of recombinant MVA-gDs
In order to obtain a safe recombinant vaccine to prevent the diseases caused by BoHV-1, we constructed a recombinant MVA containing the coding sequence for the secreted version of glycoprotein D (gDs) interrupting MVA086R viral gene (Fig. 1A). After 11 rounds of screening, the presence of the foreign gene was confirmed by PCR using specific primers (data not shown). Isolated recombinant virus MVA-gDs was pure (wild-type-virus free), as only a 1.3-kbp fragment corresponding to recombinant genome was amplified (Fig. 1B). Then expression of gD was evaluated by Western blot using polyclonal anti-BoHV-1 serum. A specific band with an apparent molecular weight of 60 kDa was detected in the supernatant of MVA-gDs-infected CEFs, corresponding to the secreted version of gD (Fig. 1C). The cellular extracts of MVA-gDs-infected CEFs also revealed reactive bands (Fig. 1C). In order to confirm the N-glycosylation of gD expressed by MVA-gDs, protein extracts from infected CEFs were treated with PnGase F enzyme as described in the material and methods section. An electrophoretic mobility shift was observed both in gD protein from the supernatant and the cellular extracts of MVA-gDs-infected CEFs when evaluated (Fig. 1C). As expected, gD was not detected in uninfected or MVA-infected CEFs. Finally, MVA-gDs genetic stability was confirmed by PCR and Western blot assays after 10 passages in CEFs (data not shown). Taken together, these results demonstrate that the secreted version of glycoprotein D of BoHV-1 is expressed by MVA-gDs.
Characterization of MVA-gDs immunogenicity
Systemic immune response induced by IP administration of MVA-gDs in mice
Once the correct expression of gD by MVA-gDs was confirmed, we evaluated its immunogenicity in the mouse model. One week after the booster, an increase in the level of humoral response was observed in MVA-gDs- and BoHV-1-vaccinated animals (Fig. 2). In particular, two doses of recombinant MVA-gDs induced a strong systemic humoral immune response specific to gD (MVA-gDs #2 versus MVA #2, p < 0.05; Fig. 2). In this regard, at 36 dpi (15 d post-booster) MVA-gDs-inoculated mice showed an anti-gD antibody ELISA titer >64,000. As expected, wild-type MVA immunization was not able to induce anti-gD antibodies independently of the number of doses applied. The group vaccinated with BoHV-1 was included as a positive immunization group and developed detectable levels of specific anti-gD antibodies when two doses were administered. The MVA-, MVA-gDs-, and BoHV-1-immunized groups presented anti-BoHV-1 seroneutralizing titers of <0.7, 0.9, and 1.9, respectively, 14 d after the booster. Importantly, the strong immune response achieved when two doses of MVA-gDs were administered IP was detected for at least 210 dpi (the last time point evaluated) (Fig. 2). At 161 dpi, mice inoculated with MVA-gDs had an anti-gD antibody ELISA titer >16,000. These results suggest that two doses of MVA-gDs resulted in the priming of the mouse immune system and conferred a strong and long-lasting systemic humoral immune response specific to BoHV-1 gD.
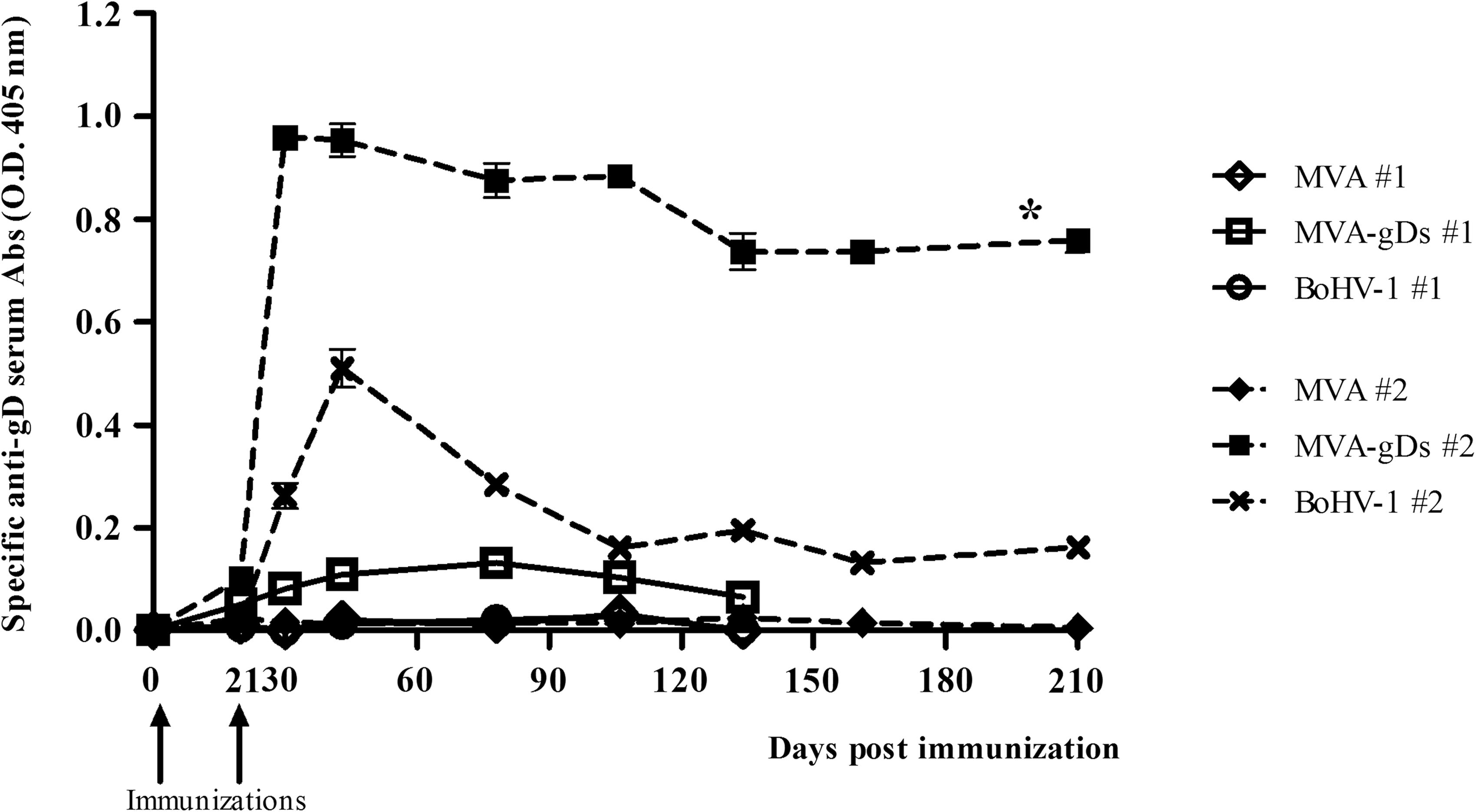
The systemic immune response induced by IP administration of MVA-gDs in mice. Mice were immunized once (#1, day 0) or twice (#2, days 0 and 21) with MVA, MVA-gDs, or BoHV-1 purified viruses. Serum samples were evaluated by ELISA for detection of anti-gD antibodies. The curves represent the average values of the measured optical density (OD) at 405 nm over time. Each sample was assayed in triplicate (*MVA #2 and MVA-gDs #2 means were statistically different, p < 0.05).
Efficacy of MVA-gDs in a rabbit BoHV-1 challenge model
As the mouse model is adequate to initially evaluate the induction of immune responses, BoHV-1 challenge experiments must be performed in other hosts like rabbits or cattle. First, the kinetics of the humoral immune response was evaluated in New Zealand rabbits immunized IM with two doses of MVA or MVA-gDs. This inoculation route was immunogenic and the maximal anti-gD antibody response was reached 9 d after booster (data not shown). Rabbits immunized with two doses of MVA did not develop any specific immune response to gD. Then, in order to evaluate the protection conferred by MVA-gDs, animals were immunized IM with MVA or MVA-gDs (0 and 17 dpi), and challenged IN with BoHV-1 (26 dpi). Rabbits in the MVA-gDs-vaccinated group developed a specific humoral immune response, which was further increased after booster, being maximal at the day of BoHV-1 challenge (26 dpi) (Fig. 3A). As expected, only the group immunized with MVA-gDs presented anti-gD antibodies before challenge (Fig. 3A and C, upper panel). At this time point, in vitro anti-BoHV-1 seroneutralizing titers of <0.7, 1.1, and <0.7, respectively, were detected in serum samples obtained from unvaccinated MVA-gD- and MVA-immunized rabbits. After challenge, clinical signs were observed in animals from all groups, but they were completely recovered after 7 d. In unvaccinated and MVA-vaccinated animals viral excretion from nasal swabs was detected from 2 to 6 d after challenge, with maximal mean BoHV-1 titers of 1.03 and 2.1 log TCID50/mL, respectively (Fig. 3B). Furthermore, only one rabbit from the MVA-gDs-vaccinated group presented BoHV-1 shedding (maximal group mean titer of 0.87 log TCID50/mL; Fig. 3B). Interestingly, the MVA-gDs-vaccinated group had the shortest viral excretion period, occurring between days 3 and 5 after challenge. The mean BoHV-1 titer obtained in all groups from 2 to 6 d after challenge was analyzed using the Friedman test, indicating that viral excretion means were statistically different (p = 0.0239). Dunn's multiple comparison test revealed significant differences in BoHV-1 shedding between MVA- and MVA-gDs vaccinated groups (p < 0.05) (Fig. 3B).
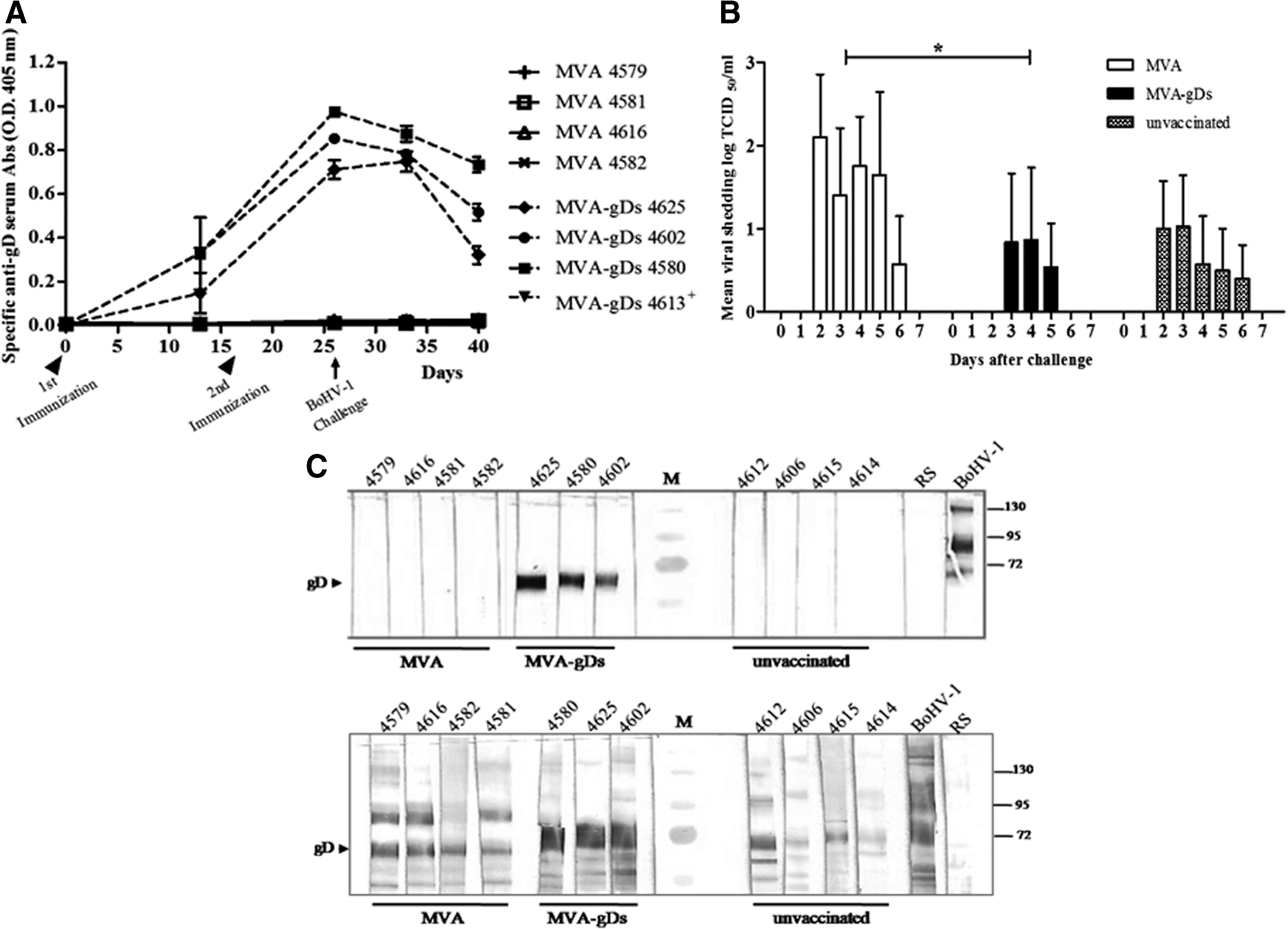
Efficacy induced by MVA-gDs in a rabbit BoHV-1 challenge model. Each animal is identified by a number. (
Fourteen days after challenge (40 dpi) serum samples from vaccinated and unvaccinated animals showed anti-gD as well as anti-BoHV-1 antibodies (Fig. 3C, lower panel), confirming the successfulness of the BoHV-1 infection procedure.
Mucosal immune response induced by IN administration of MVA-gDs formulated with cholera toxin in mice
Since natural BoHV-1 infections in cattle occur mainly through mucosal membranes of the upper respiratory tract, we evaluated recombinant MVA-gDs as a possible mucosal candidate vaccine. BALB/C mice were immunized IN with one or two doses of MVA-gDs combined with cholera toxin as mucosal adjuvant. Animals receiving one dose of MVA-gDs had very low but detectable levels of anti-gD antibodies at the analyzed mucosal surfaces (data not shown). However, two doses of MVA-gDs induced a specific humoral immune response with presence of IgA antibodies in bronchopulmonary and nasal washes. In both cases, the highest response was detected at 14 d after booster (Fig. 4A and B). Additionally, IN administration of two doses of MVA-gDs combined with cholera toxin also induced a systemic IgG anti-gD immune response (Fig. 4C).
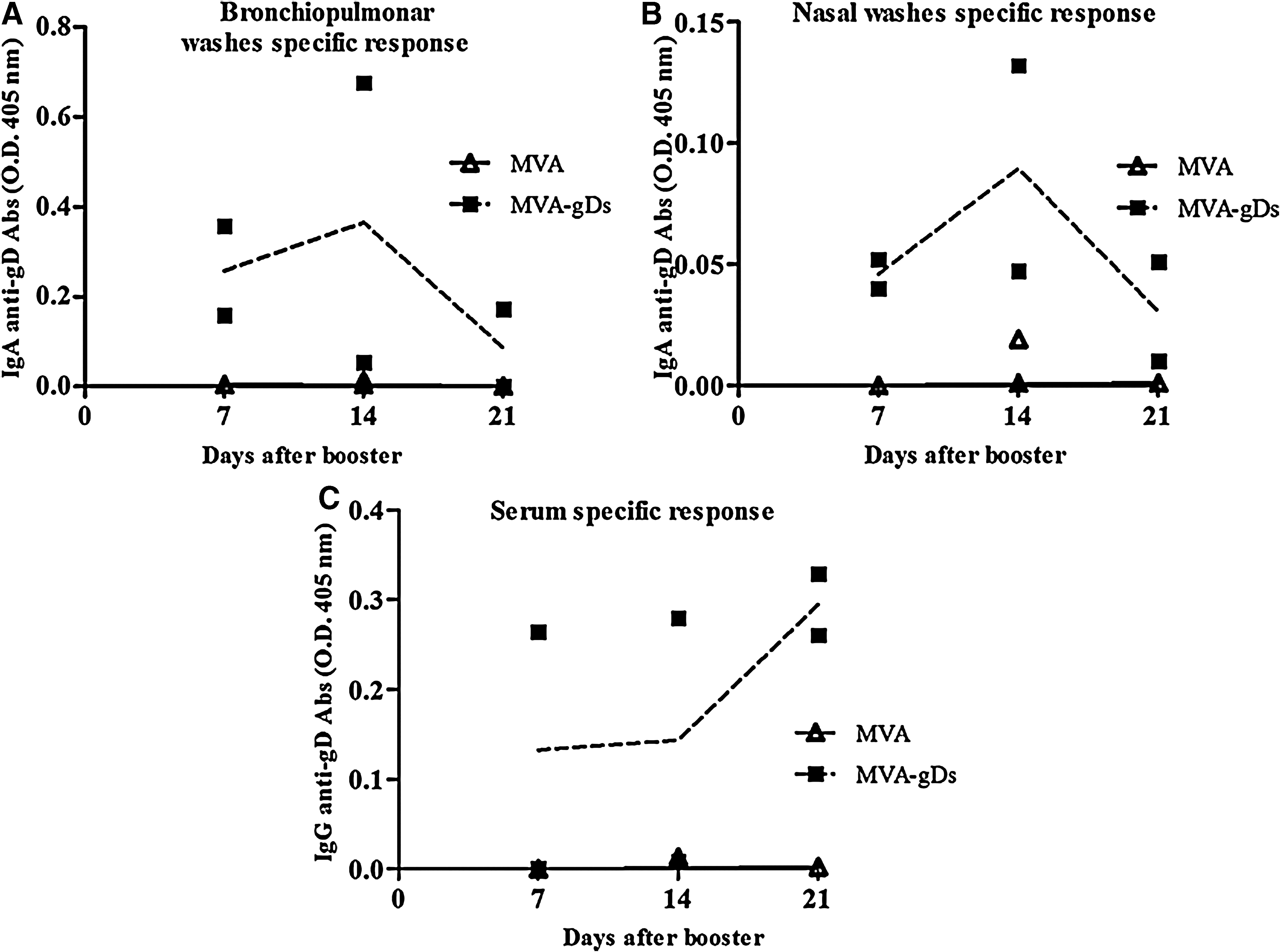
Mucosal immune response induced by IN administration of MVA-gDs in mice. Groups of six mice were immunized with two doses (days 0 and 14) of MVA or MVA-gDs co-administered with cholera toxin. Samples were evaluated by ELISA. Bronchopulmonary (
Discussion
The aim of this work was the development and evaluation of a recombinant MVA virus expressing secreted glycoprotein D (gDs) of BoHV-1. Isolated recombinant MVA-gDs virus secreted gDs protein efficiently into culture medium. The gDs band pattern observed in MVA-gDs-infected CEFs could correspond to different stages of protein glycosilation, in agreement with results reported previously by others (19). The oligosaccharides present in gD play an important role in antigenicity. The N-glycosylation of gDs expressed from MVA-gDs was clearly demonstrated by treatment with PnGase F, which removed the already synthesized N-oligosaccharide chains, altering the electrophoretic mobility of gDs. Then we focused our study on the evaluation of the humoral immune response elicited by MVA-gDs in two animal models: mice and rabbits. First, we demonstrated that IP inoculation of mice with two doses of MVA-gDs was able to induce a specific strong immune response in the mouse model (p < 0.05), which was detected even after 7 mo post-initial immunization. It has been reported that to induce a long-lasting humoral immune response, heterologous prime/boost schemes (DNA/poxvirus) must be used (12,13,26). Nevertheless, the present report demonstrated that inoculation with solely recombinant MVA viruses can develop long-lasting immunity. Importantly, MVA vector-induced immunity (due to priming) did not interfere with the specific anti-gD immunity developed after boosting with the same recombinant MVA, in agreement with previous reports (16,34). Regarding cellular immunity, IFN-γ secretion of in vitro re-stimulated splenocytes was evaluated as described by Rodriguez et al. (26). It was not possible to detect differences in IFN-γ secretion when splenocytes obtained from MVA- or MVA-gDs-immunized mice were in vitro re-stimulated with inactivated purified BoHV-1 or gD recombinant protein. In contrast, an anti-MVA cellular immunity was clearly detected in both MVA- and MVA-gDs-immunized mice (using inactivated purified MVA, data not shown). However, the fact that MVA-gDs immunization failed to induce detectable anti-gD cellular immune responses was not surprising, since the target sequence (gDs) was directed neither to the cytoplasm nor to the membrane.
Rabbits have been used as an animal model to study pathogenesis of acute and latent infections with BoHV-1 and BoHV-5 (9,25). So, once the immunogenicity of MVA-gDs was confirmed, we decided to evaluate its efficacy using a rabbit challenge model described previously (30). In the challenge assay, rabbits were inoculated IM with MVA or MVA-gDs and were infected with BoHV-1 9 d after the booster. In BoHV-1 challenge experiments, protection is commonly evaluated in terms of virus shedding levels, excretion periods after infection with the virulent strain, and severity of clinical disease compared with unvaccinated animals. The nasal swab analysis indicated that only one animal from the MVA-gDs-vaccinated group had BoHV-1 shedding, in contrast to unvaccinated and wild-type MVA-vaccinated rabbits. Overall, significant differences between the MVA and MVA-gDs-vaccinated groups were detected (p < 0.05), indicating that immunization with two doses of MVA-gDs induced some degree of protection against BoHV-1 infection, as evidenced both by a shorter viral excretion period and a decrease in the number of rabbits showing viral shedding.
In a primary BoHV-1 infection antibodies are of lesser importance than cell-mediated immunity. In contrast, in secondary infections antibodies are probably more important in preventing infection and aiding viral clearance through neutralizing antibodies and/or antibody-dependent cellular cytotoxicity (ADCC) mechanisms (4). Currently available vaccines against BoHV-1 generally induce strong humoral immune responses, but fail to develop cellular immunity. Consequently, the success of herd vaccination has been correlated with neutralizing antibody responses and the protection level achieved after viral challenge (33). Unfortunately, there are no such correlations established either for the mice or the rabbit model employed here. Besides, the results obtained suggest that to achieve higher seroneutralizing titers, more than two doses of immunogen are required. Our results agree with data published previously (3,18,29,32), and suggest that other glycoproteins of BoHV-1 may contribute to neutralize viral infection in vitro and in vivo, independently of the animal model used. Consequently, we postulate that gD alone may be not be sufficient for complete protection, and inclusion of gC and gB along with gD may be necessary for complete protection against BoHV-1.
In addition, an effective candidate vaccine against a mucosal pathogen must prevent its replication at the site of infection. Primary replication of most α-herpesviruses occurs in epithelial cells of the upper respiratory tract. BoHV-1 anti-gD antibodies have been reported to possess neutralizing capacity even at the mucosal level, eliciting IgA and IgG1 responses in cattle nasal mucosa (37). Interestingly, the effectiveness of mucosal vaccines using recombinant poxvirus vectors is enhanced by the addition of adjuvants, such as toxins, CpG oligonucleotides, or cytokines (11). In one study, cholera toxin was co-administered IN with MVA expressing HIV-1 Env IIIB antigen, resulting in enhanced humoral and cellular immunogenicity of the MVA vector (12). Considering the promising results we obtained when MVA-gDs was assayed through systemic routes of inoculation, we evaluated its utilization as a mucosal candidate vaccine. Two doses of MVA-gDs co-administered with cholera toxin were effective in inducing an IgA-specific immune response in nasal and bronchopulmonary washes, as well as a systemic humoral specific response as evidenced by the presence of IgG anti-gD antibodies in serum. These results suggest that gDs could be a useful component of a mucosal vaccine against BoHV-1 infection. In the future, a challenge test will be done administering MVA-gDs IN in the rabbit model in order to test whether the protection conferred is greater than that afforded upon parenteral immunization.
Conclusion
In the present report, a recombinant MVA expressing the secreted form of glycoprotein D of BoHV-1 was constructed and molecularly characterized. Its immunogenicity was tested in mice, in which it induced anti-gD antibodies. In a BoHV-1 rabbit challenge model, MVA-gDs induced specific humoral immunity and reduced viral replication in the upper respiratory tract. Also, when MVA-gDs was evaluated as a mucosal vaccine in mice it induced anti-gD IgA responses in nasal and bronchopulmonary washes, as well as systemic immunity. In all cases, two immunizations with MVA-gDs were necessary to induce these specific immune responses.
In conclusion, the results obtained in this work encourage further studies to evaluate MVA-gDs as a candidate vaccine for BoHV-1 in cattle challenge assays.
Footnotes
Acknowledgments
Authors are grateful to Mr. S. Díaz for animal assistance and to Mrs. M.J. Mónaco for technical assistance. This work was supported by grants AEGR 2414 and AERG 232141 from Instituto Nacional de Tecnología Agropecuaria (INTA), and PICT 2002-11195 from Agencia Nacional de Promoción Científica y Tecnológica (ANPCyT).
Author Disclosure Statement
No competing financial interests exists.