
Abstract
A live system to release heterologous antigens using an attenuated Salmonella strain was developed. We transformed Salmonella typhimurium LVR03 (S. LVR03) with a recombinant pTECH2 vector encoding 0, 1, 2, and 4 tandem copies of an imunogenic peptide of bovine herpes virus-1 (BoHV-1) glycoprotein D (gD). The system used yielded peptides fused to the non-toxic C fragment of the tetanus toxin (TetC), which has been shown to have adjuvant properties. Inoculation of BALB/c mice with the transformed Salmonella strains gave rise to a mild self-limited infection, with primary replication of bacteria occurring in Peyer's patches, even when the bacteria was administered intranasally. Humoral and cellular immune responses directed against the BoHV-1 antigens were evaluated after oral or intranasal administration of the recombinant bacteria. The results showed that the S. LVR03-dimer vaccine induced specific humoral (IgG in serum and IgG1 and IgA in saliva), and cellular immune responses (lymphoproliferation and lymphokine secretion), against not only the selected peptide and whole gD, but also against BoHV-1, when administered intranasally. This is the first time Salmonella has been used as an expression vector to induce immunity against BoHV-1. This work demonstrates the feasibility of using this antigen-release system and encourages future experimentation with a bovine experimental model.
Introduction
Vaccination with conventional attenuated or inactivated vaccines cannot prevent either the efficient transmission of the virus or the establishment of latency; however, they may protect individual animals against clinical disease. Additionally, live-attenuated virus vaccines are not entirely safe, because they may cause abortion and latency (31), with subsequent reactivation and viral shedding. Most importantly, the available vaccine strains, based on wild-type virus, may downregulate the cell surface expression of major histocompatibility complex (MHC) class I molecules (24), which likely compromises the development of cytotoxic T lymphocytes (CTL) against not only BoHV-1, but also other viruses and intracellular pathogens. On the other hand, inactivated viral vaccines are generally poor inducers of cellular immune responses, and have a relatively short duration of immunity (8). Because of these limitations, there is a need for novel vaccines that contain as few viral products as possible, but that are able to induce efficient cell-mediated and humoral immune responses.
The main target of the host protective immune response against BoHV-1 is the glycoprotein D (gD) (16), which elicits the production of CD8+ CTL (7,8), and specific neutralizing antibodies (2,25). Apparently gD is responsible for the penetration of the virus into the host cells (5), and participates in viral adsorption and membrane fusion (26).
It has been shown that plasmids encoding BoHV-1 gD induce immune responses in mice and cattle (29). However, the potency of naked DNA vaccines is limited by their inability to amplify and spread. The live bacteria vaccines (LBVs) offer a better option than plasmid vaccines to increase the magnitude of the immune response. The universal approach with LBVs has normally been the use of multicopy plasmid vectors for expression of the heterologous antigen within an attenuated host strain (15). Thus we hypothesized that a vaccine based on an attenuated Salmonella strain expressing gD epitopes could provide a strategy for protection against BoHV-1. Mapping of gD neutralization-sensitive B-cell epitopes (1) showed that synthetic peptides corresponding to residues 92–106 and 106–202 competed with the virus for neutralizing monoclonal antibodies. On the other hand, the region between residues 161 and 172 was shown to contain T-cell epitopes that are recognized by T-helper lymphocytes in primed bovine animals (27).
A single dose of an attenuated Salmonella strain expressing multiple tandem copies of herpes simplex virus gD, fused to a non-toxic fragment of tetanus toxin, was able to protect mice against viral challenge with the same efficacy as a vaccine based on the attenuated virus (4). Fraillery et al. (12) demonstrated the efficiency of Salmonella serovar typhi expressing human papillomavirus proteins to induce cellular and humoral responses in mice, the latter yielding virus-neutralizing antibodies in serum and genital secretions. Additionally, intranasal (IN) immunization of mice with Salmonella serovar typhi CVD 908-htrA expressing B- and T-cell epitopes of a surface protein of Plasmodium falciparum induced production of IgG and interferon-γ (IFN-γ) (6).
The aim of this study was to evaluate the immune response elicited in mice by a Salmonella typhimurium LVR03 (aro A) strain expressing a peptide corresponding to residues 160–216 (P56) of BoHV-1 gD, which includes both T-cell- and neutralization-sensitive B-cell epitopes, according to the studies described above. With this aim, the bacteria were transformed with a recombinant pTECH2 vector containing tandem copies of the corresponding DNA fragment. The peptides produced using this system were fused to a non toxic fragment of tetanus toxin, which has been frequently used as a carrier to enhance the response to poorly immunogenic antigens or to overcome unresponsiveness to certain peptides (28).
The results of this study demonstrate that Salmonella live bacteria vaccines are an efficient delivery vehicle for foreign antigens, inducing specific immune responses. The ability of the live vector to induce such humoral and cell-mediated immunity likely reflects the capacity of Salmonella to provide immunomodulatory signals that enhance B- and T-cell function.
Materials and Methods
Plasmid and oligonucleotides
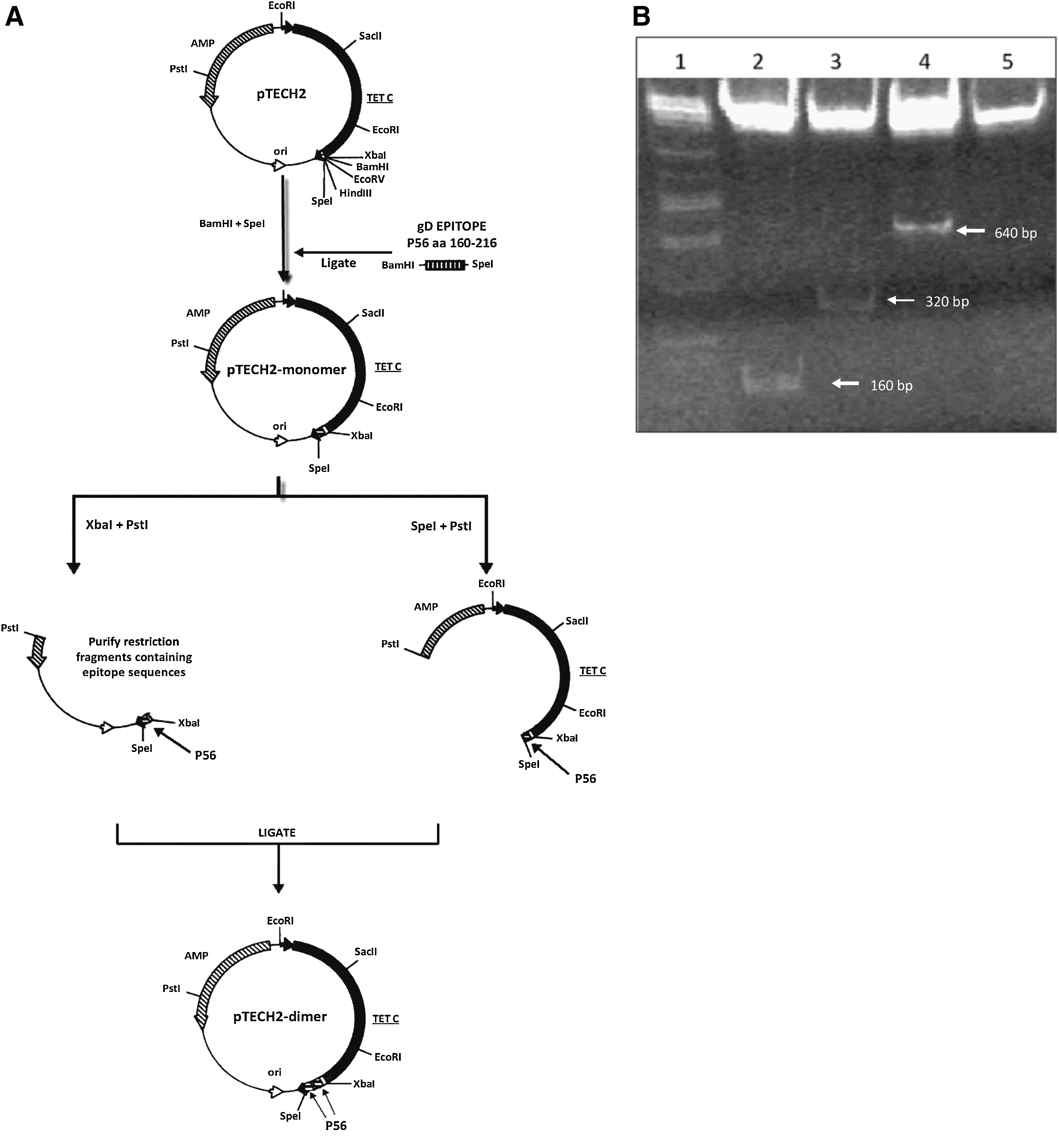
Plasmid pTECH2 has been previously described (19). For preparation of pTECH2 fusion constructs, a pair of complementary oligonucleotides encoding the aa 160–216 peptide (P56) of BoHV-1 gD (GenBank accession number CAA80604; (ctgattatggcggcgcccgcgcggctcgtcgagggccagtaccgacgcgcgctgtacatcgacggcacggtcgcctatacagatttcatggtttcgctgccggccggggactgctggttctcgaaactcggcgcggctcgcgggtacacctttggcgcgtgcttcccggcc) were designed, and tailored with BamHI and SpeI cohesive ends, which were generated upon annealing. The double-stranded oligonucleotide was cloned into pTECH2, which had been previously digested with BamHI and SpeI and transformed into E. coli. The preparation of fusion constructions containing tandem repeats (1–4 copies) of this sequence was carried out as previously described (19).
Briefly, aliquots of the recombinant fusion vector were simultaneously digested with either XbaI and PstI or with SpeI and PstI, and the two types of digested inserts were purified and ligated head to tail. The overhangs generated by XbaI and SpeI are compatible, but the recognition sites for these enzymes were destroyed upon ligation. Thus the XbaI and SpeI sites flanking the peptide-coding sequence remain unique, and the procedure could be serially repeated, doubling the copy number of peptide-coding sequences with each cycle (Fig. 1A). Sequencing of the obtained pTECH2-monomer DNA fragments and sequence analysis by Vector NTITM 8.0 verified that the expected constructs had been produced.
(
Plasmid construction
Plasmid pCIneogD was constructed by inserting a PCR product encoding the secreted version of the BoHV-1 gD in EcoRI-digested pCIneo vector (Promega, Madison, WI). Forward primer 5′-AAGAATTCCGGCTGCTGCGAGCGGGCGAACA-3′ and reverse primer 5′-AAGAATTCTCAGGCGTCGGGGGCCGCGGGCGTA-3′, both containing an EcoRI restriction site, were used to amplify a 1121-bp DNA fragment, which corresponds to 118874–119975 bp of the BoHV-1 gD gene (GenBank accession number Emb
Analysis of the recombinant pTECH2 constructs
Plasmids were digested with XbaI and SpeI, followed by 12% polyacrylamide gel electrophoresis and silver staining.
Bacterial strain
The bacterial strain used was Salmonella typhimurium LVR03 (aroA). Bacteria were grown aerobically on Luria-Bertani (LB) broth supplemented with 100 μg/mL ampicillin as required. Plasmid pTECH2, monomer, dimer, and tetramer were introduced into Salmonella typhimurium LVR03 (S. LVR03) using an electroporator (Bio-Rad, Hercules, CA). Vaccine inocula were prepared as follows: individual colonies were resuspended in LB (with ampicillin), and grown for 16 h at 37°C. Bacteria were harvested by centrifugation and resuspended in 0.2 mL of sterile PBS to a final concentration of 109 CFU in 15 μL, and 1011 CFU in 200 μL. The number of viable organisms was determined by plating serial dilutions of the inoculum onto LB agar with ampicillin as required.
Peptide aa 173–202 (P29)
Synthetic peptide within the amino acid sequence between residues 173 and 202 of gD BoHV-1 was synthesized using the t-Boc method on a DuPont coupler 2200 automated peptide synthesizer.
Vaccine formulations and animal treatment
Experiments with animals were performed in accordance with the guidelines of the institutional committee for the care and use of experimental animals.
Eight-week-old male BALB/c mice were used. The animals (obtained from Facultad de Ciencias Veterinarias de La Plata, Buenos Aires, Argentina) were acclimated and randomly distributed into experimental groups. The mice were kept in conventional animal facilities and received water and food ad libitum. Groups of mice were orally or IN inoculated with 1×1011 CFU/200 μL or 1×109 CFU/15 μL, respectively, with the different vaccines. The same dose and schedules for injection were used for the booster at day 35 after the first inoculation, and 7 d before the cellular immune response studies (67 dpv). Negative control groups were inoculated with S. LVR03-pTECH2, and the other groups were inoculated with S. LVR03-monomer, dimer, or tetramer. Serum samples were taken on days 0, 25, and 60 after the first inoculation.
P29 vaccine
Vaccination was performed IP (200 μL per mouse) with 10 μg of synthetic peptide containing aa 173–202 of the gD proteín of BHV-1, diluted in PBS to reach the final volume with adjuvant INTA (Bagó) and Tween.
pCIneo-gD
The adjuvant Cliptox™ is a variety of zeolite named clinoptilolite. The adjuvant was used according to the manufacturer's instructions. Cliptox was dissolved in PBS and used at 2 (g of Cliptox per (g of plasmid. In order to evaluate the safety of Cliptox, toxicity was assessed by the HET-CAM test at the Centro de Toxicología y Biomedicina (TOXIMED), Santiago de Cuba, Cuba. This procedure has been validated by the European Center for the Validation of Alternative Methods (ECVAM), Ispra, Italy. Cliptox was classified as safe.
At day 60, groups of 5 mice were intradermally (ID) inoculated in the back with 15 μg DNA (pCIneogD) formulated with Cliptox at a final volume of 0.2 mL.
Virus preparation
BoHV-1 strain LA (Los Angeles) was propagated in MDBK cells, supplemented with 10% fetal bovine serum (FBS; Gibco, Carlsbad, CA). Confluent monolayers in roller bottles were infected with the virus at a multiplicity of infection (MOI) of 0.1, and the virus was allowed to adsorb for 45 min at 37°C before the addition of fresh culture medium. When extensive cytopathogenic effects were observed, the supernatant was collected and cell debris was removed by centrifugation at 3000×g for 30 min. The virus suspension was stored at −70°C. Exposure to UV light for 30 min was used for viral inactivation.
Enzyme-linked immunosorbent assay for detection of anti-gD antibodies
Briefly, Immulon 2 (Dynatech Laboratories, Chantilly, VA) microtiter plates were coated with 1.4 μg/well of purified rgD in 0.1 M carbonate-bicarbonate buffer, pH 9.6, and incubated overnight at 4°C. Afterwards, the plates were blocked with PBS+0.05%Tween 20+1% ovalbumin (OVA), and then incubated with the serum samples for 1 h at 37°C. Anti-mouse IgG peroxidase conjugate was added, followed by ortho-phenylene-diamine (1,2-benzenediamine) dihydrochloride (OPD) (Sigma-Aldrich, St. Louis, MO) and H2O2 as substrate. Absorbance was measured at 492 nm 20 min later. Results show the average A492±SEM of 5 individual mice at a serum dilution of 1:50.
Enzyme-linked immunosorbent assay for detection of anti-P29 antibodies
MaxiSorp® (Nalge Nunc International, Rochester, NY) plates were coated either with peptide aa 173–202 (P29; 0.625 μg/well) in 0.1 M carbonate/bicarbonate buffer, pH 9.6. Afterward, the wells were blocked with PBS/0.05% Tween 20/0.5% gelatin, and then incubated with serum dilutions, followed by anti-mouse IgG peroxidase conjugate (1 h at 37°C) and OPD/H2O2 as colorimetric substrate. Absorbance was measured at 492 nm.
Enzyme-linked immunosorbent assay for detection of anti-LPS antibodies
Briefly, Immulon I (Dynatech Laboratories) microtiter plates were coated with 100 μL of 0.1 M carbonate/bicarbonate buffer, pH 9.6, containing 1 μg Salmonella LPS (Sigma-Aldrich) per well, and blocked with PBS+0.05% Tween 20 and 2% blocking reagent. Then the plates were sequentially incubated with serum dilutions of anti-mouse IgM peroxidase conjugate (1 h at 37°C), and OPD/H2O2 absorbance was measured 20 min later at 492 nm. Results show the average A492±SEM of 5 individual mice at a serum dilution of 1:50.
Enzyme-linked immunosorbent assay for detection of anti-Salmonella antibodies
Briefly, MaxiSorp microtiter plates were coated with 100 μL of 0.1 M carbonate/bicarbonate buffer, pH 9.6, containing 1×106 CFU Salmonella per well, and blocked with PBS+0.05% Tween 20 and 3% milk. Then the plates were sequentially incubated with serum dilutions of anti-mouse IgM peroxidase conjugate (1 h at 37°C), and OPD/H2O2. Absorbance was measured 20 min later at 492 nm. Results show the average A492±SEM of 5 individual mice at a serum dilution of 1:50.
Enzyme-linked immunosorbent assay for detection of anti-BoHV-1 antibodies in serum or saliva
Immulon I (HB) plates were coated with concentrated and semi-purified BoHV-1 (106 TCID50/mL) in 0.1 M carbonate/bicarbonate buffer, pH 9.6. Afterward, the wells were blocked with PBS+0.05% Tween 20/0.5% OVA (BoHV-1), and then incubated with serum or saliva dilutions followed by anti-mouse IgG peroxidase conjugate (1 h at 37°C) and OPD/H2O2 as colorimetric substrate. Absorbance was measured at 492 nm. Saliva was considered positive when the A492 was higher than the average A492 of the negative samples±2 standard deviations.
Isolation of spleen cells
Mice were killed by cervical dislocation and dissected. The spleens were carefully removed and placed in sterile Petri dishes. After cutting off their bottom ends, about 2 mL of PBS supplemented with 10 mM EDTA was injected into each spleen, in order to draw the cells from the capsules. The cells were centrifuged, counted in a Neubauer chamber, and adjusted to optimal concentrations.
T-cell proliferation
Seven days after the third booster (67 dpv), spleen cells were isolated from the animals (n=2), pooled, and suspended in RPMI 1640 medium supplemented with 10% fetal calf serum (FCS; Gibco), 2 mM L-glutamine (Sigma-Aldrich), 1 mM sodium pyruvate (Gibco), 5 mM HEPES (Gibco), 50 mM 2-mercaptoethanol, 1% (w/v) ampicillin-streptomycin (Sigma-Aldrich), and 10 mM MEM non-essential amino acid solution (Gibco). Aliquots of 100 μL containing 2.5×105 cells were distributed in individual wells of flat-bottom 96-well microtiter plates and incubated for 3 days at 37°C, either with medium alone, rgD (1.4 μg/μL), inactivated BoHV-1 (iBoHV-1), or P29 (0.625 μg/well). Incorporation of radioactivity into cellular DNA was measured after 18 h of incubation with 1 μCi of [methyl-3H] thymidine (Amersham Biosciences, Piscataway, NJ) per well. Cells were harvested using a semiautomatic cell harvester, and the amount of incorporated label was measured by liquid scintillation counting (Winspectral; Wallac, Roxbury, MA). The results are shown as the increase in cpm with respect to the negative control.
IFN-γ measurements
Secretion of IFN-γ from spleen cells stimulated for 48 h with rgD, or concanavalin A was assessed. IFN-γ (pg/mL) measurements were determined in the supernatants of cell cultures previously stimulated with medium, rgD (1.4 μg/μL) or ConA (20 μg/mL) by a commercial sandwich ELISA kit (eBioscience, San Diego, CA). Briefly, ELISA plates (MaxiSorp) were coated with rat anti-mouse IFN-γ antibody. The plates were blocked with PBST+10% FCS, and incubated with culture supernatants for 2 h at room temperature, followed by biotinylated anti-cytokine antibody (1 h at 37°C), and HRP-conjugated streptavidin. The substrate, 3,3′-5,5′-tetramethylbenzidine was added and A450 was measured 30 min later in a microplate reader. Cytokine concentrations were calculated based on the A450 measurements obtained with the commercial standards, after subtracting the average A450 of two wells incubated with medium alone.
Salmonella persistence in vivo
BALB/c mice (n=8) were intranasally immunized with S. LVR03-dimer (1×109 CFU/15 μL). At 24, 48, 96, and 120 h post-inoculation, two mice were euthanized and organs, esophagus, spleen, and Peyer's patches (PP) were extracted, polled, and homogenized in PBS. Dilutions of the homogenate (1:10) were plated on LB agar containing ampicillin (50 μg/mL), and incubated overnight at 37°C to determine numbers of bacterial colonies (CFU). The results are expressed as CFU obtained at each time point.
Statistical analysis
One-way analysis of variance (ANOVA) and Bonferroni post-hoc tests were used to compare data between three or more groups. Log10 transformation was done when necessary. p Values <0.05 were considered significant.
Results
Construction of pTECH 2-P56 (aa 160–216)
Plasmid pTECH2 (19) was kindly provided by Dr. Khan. This plasmid, which is a modification of pTECH1 (19), has an ampicillin-resistance gene, and under anaerobic conditions, an inducible promoter leads to the expression of recombinant antigens fused to a gly-pro-gly motif in the C terminus of tetanus toxin (TetC), which acts as adjuvant (28). A pair of complementary oligonucleotides encodes a 56-aa peptide (P56, residues 160–216) of BoHV-1 gD. Additionally, constructs that express protein fragments containing 2 and 4 tandem copies of the peptide fused to TetC were prepared as previously described (18).
Cloning of inserts containing 0, 1, 2, and 4 copies of P56-coding DNA into pTECH2 yielded the constructs pTECH2 and pTECH2-monomer, -dimer and -tetramer, respectively.
DNA digestion with XbaI and SpeI confirmed the presence and size of the insert (monomer: 160 bp, dimer: 320 bp, tetramer: 640 bp) and indicated the number of tandemly-cloned copies (Fig. 1B).
Salmonella typhimurium LVR03 was transformed with this constructions and expression of the TetC-P56 fusion proteins was demonstrated by Western blotting (data not shown).
Humoral responses elicited by live Salmonella P56 vaccines with different numbers of P56 copies and routes of delivery
To determine the route that elicits optimal immunological results against BoHV-1, mice (n=5 in each group) were vaccinated and boosted 35 d later with S. LVR03-pTECH2, monomer, dimer, or tetramer. Administration took place either orally or IN with 1011 CFU/200 μL or 109 CFU/15 μL bacteria per mouse, respectively. Sera were analyzed at 25 and 60 d post vaccination (dpv) using an anti-gD ELISA (32)
In the orally-immunized group, anti-gD antibodies could be detected in the S. LVR03-monomer- and tetramer-vaccinated groups at 60 dpv. For the IN immunized group, anti-gD antibodies were detected in the S. LVR03 monomer and dimer groups (Fig. 2).
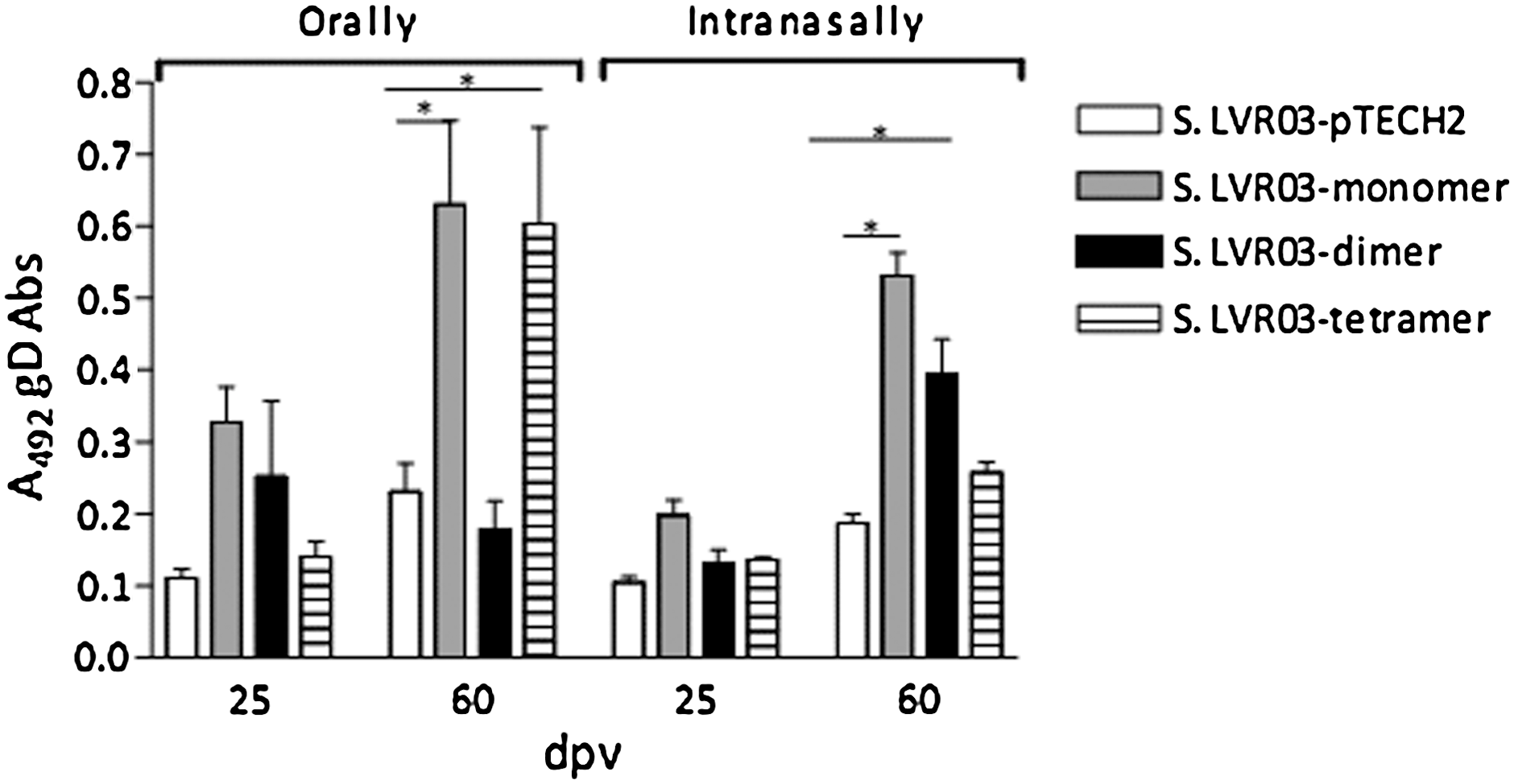
Humoral response against gD protein in mice vaccinated orally (1011 CFU/200 μL) or intranasally (109 CFU/15 μL) with S. LVR03. Groups of 5 mice were vaccinated with S. LVR03-pTECH2, S. LVR03-monomer, S. LVR03-dimer, S. LVR03-tetramer and serum antibodies (Abs) at 25 and 60 dpv and evaluated by ELISA. Results are shown as the average A492±SEM of 5 individual mice at a serum dilution of 1:50. Significant differences (p<0.05) between the average of a group and the control group are indicated by an asterisk.
It is known that IN immunization stimulates both mucosal and systemic immunity, and antigen-specific mucosal immune responses can be expressed at different mucosal sites. Since vaccination via the IN route induced a humoral response in the case of the three constructions, we investigated the induction of IgG1 and IgA in saliva of inoculated mice.
Interestingly, a high percentage of mice showed anti-BoHV-1-specific mucosal antibodies at 60 dpv, (80% IgA and 60% IgG1) when vaccinated IN with S. LVR03-monomer or S. LVR03-dimer. The group that received S. LVR03-pTECH2 or S. LVR03-tetramer did not develop anti-BoHV-1 antibodies in saliva during the time course of the experiment (Table 1), so Salmonella LVR03-tetramer was not selected for further study.
Samples were considered positive when the A492 was higher than the average A492 of the negative samples±2 standard deviations.
Results are shown as percentages of positive animals in each group and the number of positive animals/number of animals vaccinated in parentheses.
BoHV-1, bovine herpes virus-1; S. LVR03, Salmonella typhimurium LVR03; ELISA, enzyme-linked immunosorbent assay.
Groups of mice vaccinated with S. LVR03-monomer orally and S. LVR03-dimer intranasally, as well as their corresponding pTECH2 controls and peptides 173–202 (P29 vaccine) were selected to assess IgG levels against peptide aa 173–202 (P29) and BoHV-1, as well as IgM levels against whole Salmonella or Salmonella LPS at 60 dpv.
The group of mice vaccinated orally with S. LVR03-monomer developed IgG antibodies that recognized P29 and whole BoHV-1 (Fig. 3A). When the humoral response against Salmonella or Salmonella LPS was studied, higher IgM levels were found in the group inoculated orally with S. LVR03-pTECH2 than in the S. LVR03-monomer group (Fig. 3B), suggesting that S. LVR03-pTECH2 replicates better than S. LVR03-monomer when administered orally. In the case of the IN-immunized animals, specific antibodies against P29 and the virus were observed in the S. LVR03-dimer group, but not in the S. LVR03-pTECH2 group (Fig. 3C). The P29 vaccine control group had a level of IgG similar to that of the group immunized with S. LVR03-dimer. IgM levels against Salmonella or Salmonella LPS were similar in the S. LVR03-pTECH2 and S. LVR03-dimer groups (Fig. 3D), indicating that S. LVR03-pTECH2 replicates in a similar way to S. LVR03-dimer when administered IN. This result is consistent with the observation of differences in in vitro growth of each of the Salmonella groups (Fig. 4).
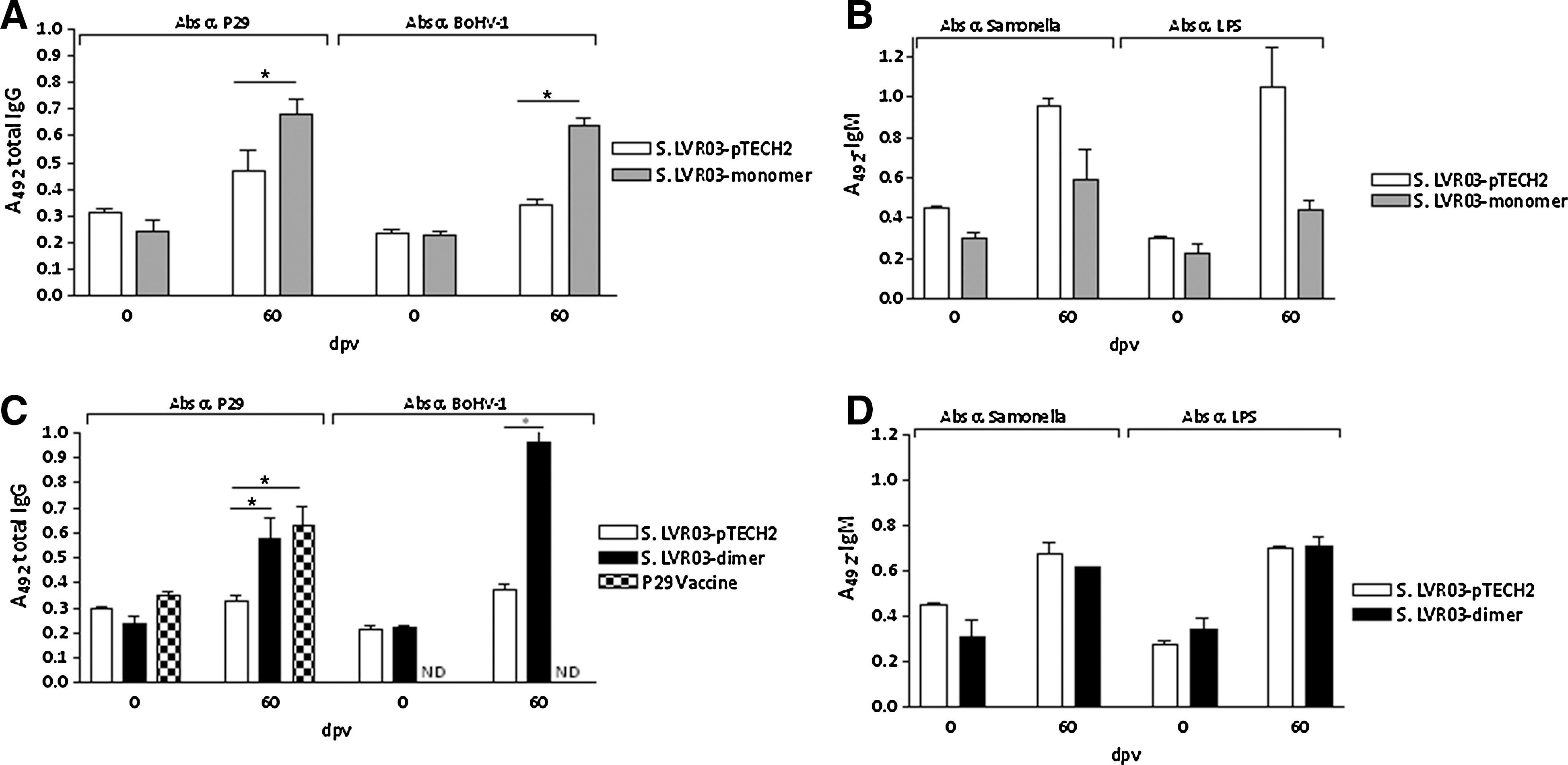
Humoral response of mice vaccinated orally (1011 CFU/200 μL) and intranasally (109 CFU/15 μL) with Salmonella LVR03, or intraperitoneally with P29. Groups of 5 mice were vaccinated with S. LVR03-pTECH2 and S. LVR03-monomer orally, S. LVR03-pTECH2 and S. LVR03-dimer intranasally, or with P29 vaccine intraperitoneally, and serum antibodies at 60 dpv were evaluated by ELISA. (
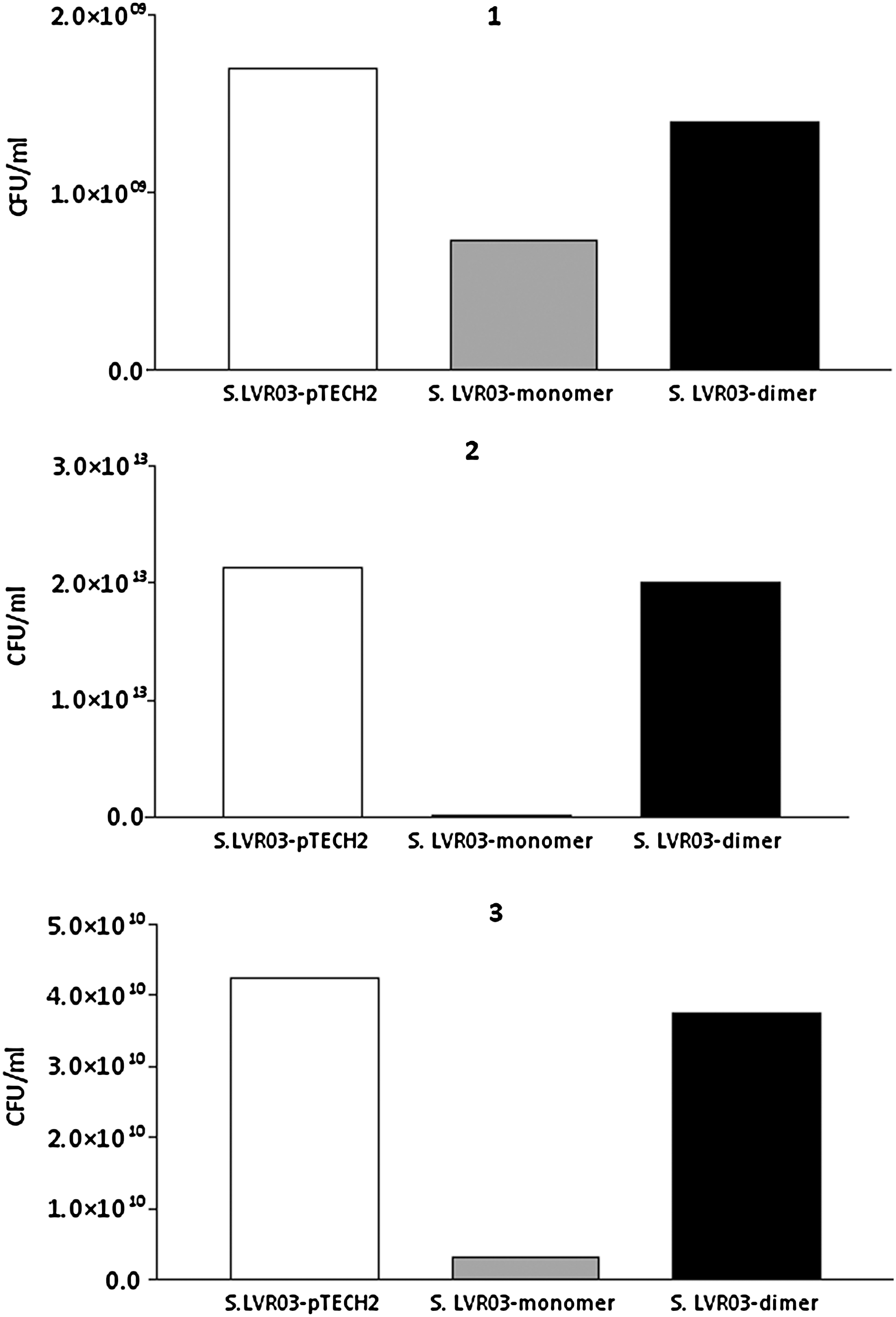
Salmonella in vitro growth. S. LVR03-pTECH2, S. LVR03-monomer, and S. LVR03-dimer were grown aerobically on Luria-Bertani (LB) broth supplemented with 100 μg/mL ampicillin for 16 h at 37°C. Bacteria were harvested by centrifugation and resuspended in sterile PBS in serial dilutions from 10−1 to 10−12, then they were plated on LB plates (with ampicillin) using 4 drops of 10 μL of each dilution and grown at 37°C overnight. Dilutions were counted if they grew 3–30 colonies per drop, and the CFU/mL was calculated.
Salmonella LVR03-dimer induces lymphoproliferation and IFN-γ secretion
Despite that fact that the humoral immune responses induced by S. LVR03-monomer and S. LVR03-dimer were similar, the cellular immunity was difficult to study in the group with S. LVR03-monomer, because the non-specific immune response in the S. LVRO3-pTECH2 group was very strong, which was likely due to the better ability to replicate S. LVRO3-pTECH2 than the S. LVR03-monomer.
Analysis of the cellular response was conducted on animals inoculated with the S. LVR03-dimer, because this group of animals developed a consistent humoral response and it was possible to evaluate the specific cellular response.
In order to induce a specific cellular response, the region of interest (P56) included in the vaccine pCIneo gD, and not against any epitope from Salmonella, the booster was performed at 60 dpv with genetic vaccine encoding for gD (pCIneo-gD) in all groups.
At 60 dpv mice vaccinated IN with S. LVR03-dimer, P29 vaccine, S. LVR03-pTECH2, or non-vaccinated mice were boosted with pCIneo-gD (a genetic vaccine coding for gD protein) and Cliptox as adjuvant, as described by Langellotti et al. (20), and 7 d later specific cell-mediated responses were measured. Spleen cells from these groups of mice were stimulated in vitro with a recombinant form of gD (rgD) (32), inactivated BoHV-1 (iBoHV-1), or P29 in a proliferative assay, or with rgD or ConA for IFN-γ determination. The antigen-specific proliferative response was significantly higher (p<0.01) in the group immunized with S. LVR03-dimer than in the S. LVR03-pTECH2 group (Fig. 5A). However, P29 vaccine showed a low level of cell stimulation.
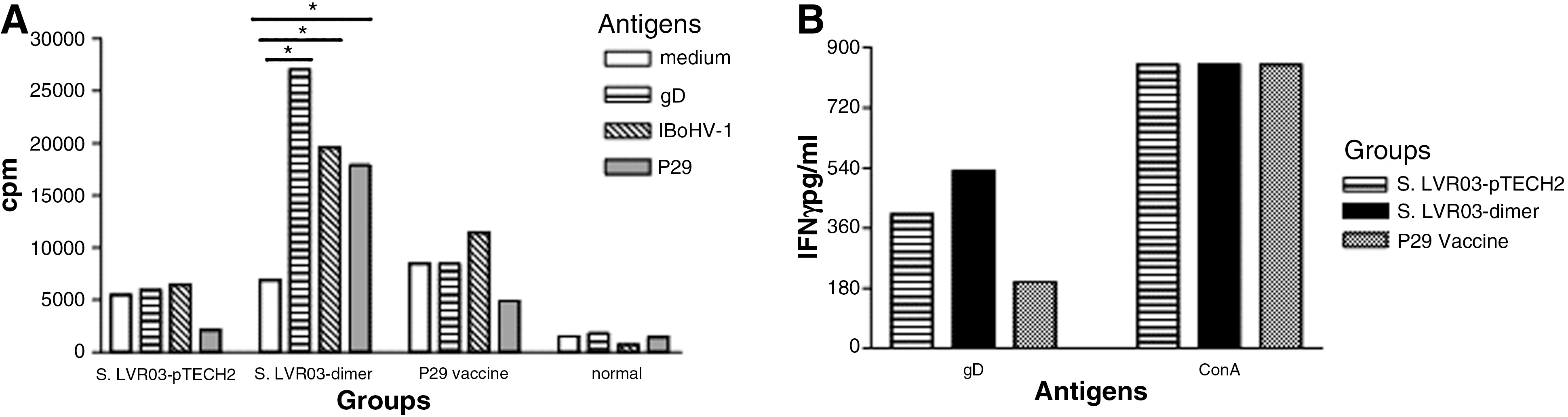
Cellular response to S. LVR03-pTECH2 and S. LVR03-dimer given IN, and IP P29 vaccination. (
To further examine the cell-mediated immune response induced by S. LVR03-dimer, we assessed the IFN-γ levels elicited in the spleens of immunized animals. Following in vitro stimulation with rgD, the S. LVR03-dimer group showed higher IFN-γ levels than the S. LVR03-pTECH2 group (Fig. 5B). The P29 vaccine group did not secrete IFN-γ in vitro.
Our experiments clearly show that the S. LVR03-dimer vaccine induced both specific humoral and cellular immune responses when administered IN.
in vivo colonization and persistence of Salmonella vaccine organisms
The ability of vaccine organisms to invade and thrive in host tissues is considered to be a critical element for live vectors to efficiently prime immune responses against heterologous antigens. We assessed the ability of S. LVR03-dimer to invade and persist in different tissues following IN delivery. At 24, 48, 96, and 120 h post-inoculation (hpi), two mice at each time point were euthanized, and the esophagus, spleen, and PP were collected and analyzed. The kinetics of recovery of bacteria in these samples is shown in Fig. 6. Positive cultures were found in the spleen at 96 and 120 hpi. The bacterial load was particularly abundant in PP and esophagus. No growth was observed in control tissues from non-vaccinated animals used as negative controls (data not shown). These results confirm that the S. LVR03-dimer administered IN can invade and persist in host tissues.
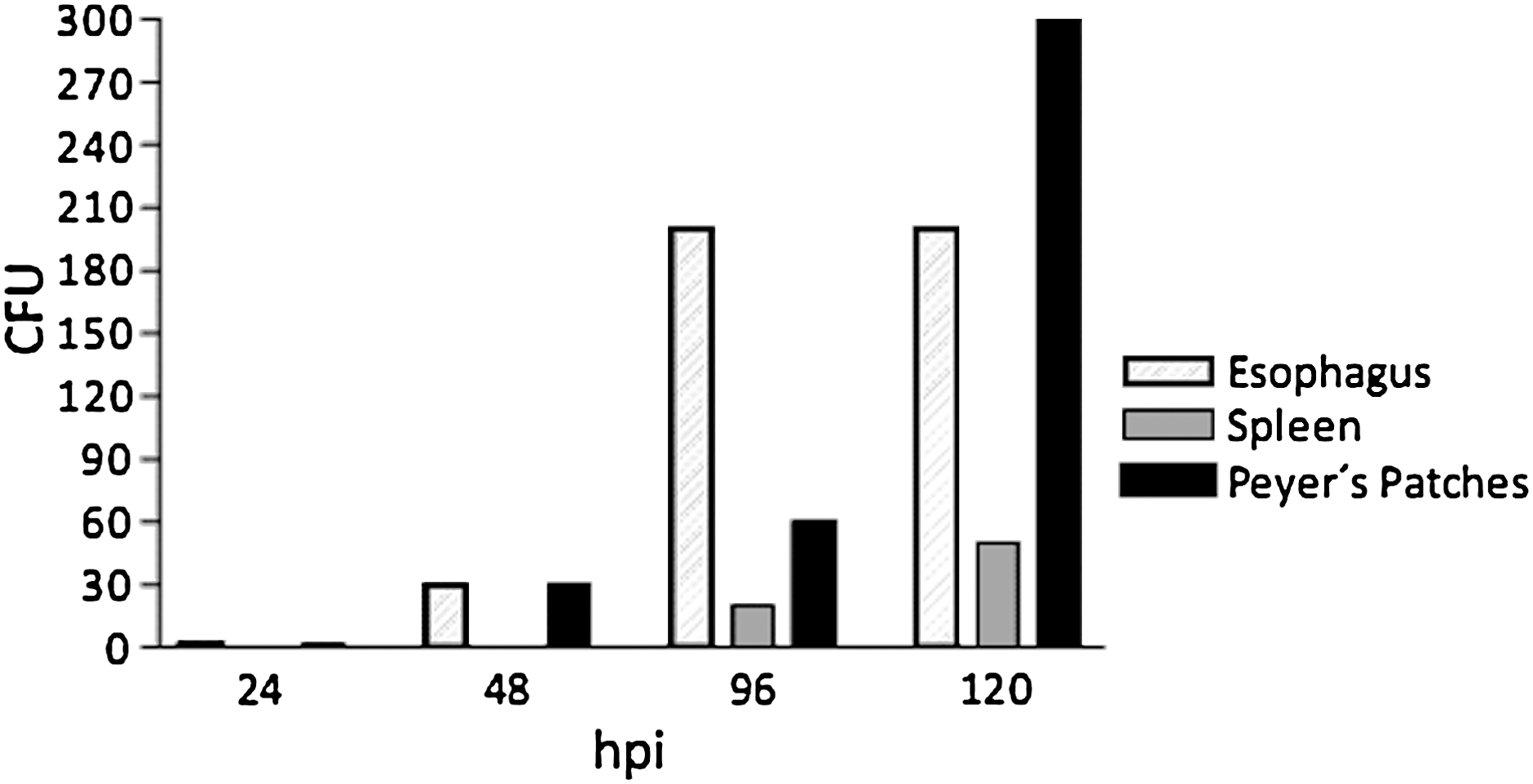
Salmonella in vivo colonization. BALB/c mice were IN immunized with S. LVR03-dimer (1×109 CFU/15 μL). At 24, 48, 96, and 120 h post-inoculation, samples from two mice were euthanized and pooled, and the esophagus, spleen, and Peyer's patches were extracted and homogenized in PBS. Dilutions of the homogenate (1:10) were plated and incubated to determine the numbers of bacterial colonies (CFU). Results are expressed as mean CFU obtained at each time point.
Discussion
Live attenuated Salmonella strains have been extensively studied to elicit mucosal as well as systemic immune responses against antigens from other infectious agents, such as E. coli, Shigella sonnie, Plasmodium falciparum, Bacillus anthracis, Helicobacter pylori, Listeria monocytogenes (3), HPV-16, and hepatitis B virus, as well as against tumor antigens (10).
Synthetic peptide immunogens displaying epitopes of pathological relevance can be used in the rational design of vaccines. However, peptides show poor immunogenicity, a disadvantage that can be overcome by coupling them to larger molecules. Alternatively, this can be achieved by synthesizing peptides in tandem repeats. For this study, multiple tandem copies (1–4 copies) of an immunogenic peptide (P56) comprising amino acids 160–216 of glycoprotein D of BoHV-1 were expressed as a TetC fusion protein in a Salmonella live attenuated vaccine.
The pTECH2 vector used, which contains the anaerobically inducible promoter nirB, allows the expression of heterologous antigens either as full-length proteins (18), or as multiple tandem copies of a relevant peptide (17). Expression of the full-length Schistosoma mansoni glutathione S-transferase P28 as a fusion to TetC has led to an experimental vaccine capable of protecting mice against salmonellosis, tetanus, and schistosomiasis, after a single dose (18). Furthermore, it has been reported that a Salmonella vaccine strain expressing tandem copies of a gD peptide of herpes simplex virus as a fusion with TetC protected mice against virus challenge (4).
In order to evaluate the vaccine potential of the recombinant S. LVR03 strains, mice were immunized with live bacteria and with P29 vaccine to compare the immunogenicity. We investigated the humoral response elicited, as measured by levels of serum IgG and mucosal IgA and IgG1 against BoHV-1, rgD, and P29 containing a T-cell epitope. In addition, the cellular immune response was assessed by the induction of viral-specific lymphoproliferation and IFN-γ secretion.
Only the groups of animals immunized with S. LVR03-dimer or S. LVR03-monomer developed IgA and IgG1 titers against BoHV-1 in saliva, while mice immunized with S. LVR03-pTECH2 or with S. LVR03-tetramer never induced these types of antibodies. These results demonstrated that our vaccine candidate can effectively induce both systemic and mucosal immune responses when the IN administration route is used.
When we compared the levels of IgG antibodies against P29 of mice immunized with S. LVR03-dimer with those of the group vaccinated with P29, no significant differences were found between them. However, the data also show that the viral-specific cellular immune response was higher in mice vaccinated IN with S. LVR03-dimer than in the control group immunized with S. LVR03-pTECH2 or with P29, which suggests that this vaccine candidate induces a Th-1 type of cellular immune response. These results are in accordance with those of Karem et al. (17), who found that the anti-tetanus toxoid response in serum of mice immunized with an aroA mutant of Salmonella expressing the Tet-C fragment was predominantly IgG2a, suggesting a Th-1 type of immune response. Our experiments showed that IN immunization with Salmonella induces IFN-γ, which is related to cellular and humoral immune responses.
It is likely that the IFN-γ levels were increased in the S. LVRO3-pTECH2 group due to the immunogenicity elicited by Salmonella.
Analysis of the cellular response was conducted in animals inoculated with the S. LVR03-dimer, because this group of animals developed a consistent humoral response, and it was possible evaluate the specific cellular response.
The cellular immunity was difficult to study in the S. LVR03-monomer group, because the non-specific immune response in the S. LVR03-pTECH2 group was very strong, likely due to the better ability to replicate in vitro of the S. LVRO3-pTECH2 compared to the S. LVR03-monomer. We were surprised to see differential growth between the S. LVR03-monomer and the S. LVR03-dimer and S. LVR03-pTECH2. So we confirmed that the sequence was correct and repeated the in vivo and in vitro experiments, and the results obtained were the same. It has been reported that when a recombinant protein is expressed, if the bacteria detects that there is an unusual type of folding, they receive a stress signal and stop growing. It is possible that the basal expression of some of the proteins (such as the monomer) inhibited growth, and this would explain the different cell density reached by Salmonella or its LPS, and why it was higher in the S. LVR03-dimer and S. LVR03-pTECH2 groups than in the S. LVR03-monomer group.
On the other hand, a requirement for an effective Salmonella vaccine vector is the efficient colonization of PP. This was the main location where bacteria were found after IN administration of the S. LVR03-dimer. Dunstan et al. (9) demonstrated that the level of total Ig elicited against a carried antigen does not necessarily correlate with the ability of the Salmonella strain to colonize and persist in the spleen, but does correlate with the ability of the strain to colonize and persist in PP following oral immunization. The importance of PP colonization in the induction of specific immune responses following oral immunization with attenuated S. typhimurium was originally indicated in studies by Galan et al. (13). S. typhimurium harboring an attenuating mutation in phoP (of the phoP-phoQ two-component regulatory system) (22) was able to colonize the PP of orally-immunized, mice but were relatively deficient in splenic colonization. This supports our results of the analysis of colonization after IN administration of the S. LVR03-dimer.
The results presented in this study reinforce previous observations (4,17,18) on the feasibility of developing vaccines by constructing C-terminal fusions of recombinant bacterial, viral, or parasitic antigens with TetC using the pTECH2 vector and live attenuated Salmonella strains as a delivery system. Thus our results support the use of peptides engineered to be expressed in a recombinant Salmonella vector as a vaccine to elicit cellular and humoral immune responses. Importantly, this is the first time that Salmonella has been used as a vector vaccine against BoHV-1.
Footnotes
Acknowledgments
The authors wish to thank Osvaldo Zabal for valuable assistance with cell cultures, to Dr. O. Taboga and V. Bianco for helping with plasmid construction, and to Dr. E. Mercado for biochemical tests for Salmonella. This project was funded by two research grants from the Agencia Nacional de Promoción Científica y Tecnológica (PIC
Author Disclosure Statement
No competing financial interests exist.