
Abstract
Cytomegalovirus(CMV) reactivation causes immunopathy, graft malfunction, and even rejection. The traditional anti-CMV drug ganciclovir is not able to prevent reactivation of endogenous virus. Recent studies have found that proteasome inhibitor (PI) is able to suppress CMV replication. In this study we investigated the influence of proteasome inhibitor MG132 and ganciclovir on the CMV-specific CD8+ T-cell immune response. We found that interferon-γ (IFN-γ) production in response to CMV-infected fibroblasts was reduced under the influence of MG132 in a dose-dependent manner. A marked reduction was observed at 0.5 μM. Likewise, CMV-specific cytotoxicity of CD8+ T cells was decreased in the presence of MG132. In contrast, the traditional CMV replication inhibitor ganciclovir (10 μM) had no such effect. These findings might have important implications in reducing CMV-associated immunopathy by altering epitope generation through the application of selective proteasome inhibitors.
Introduction
Materials and Methods
Cell and virus
Human embryonic lung fibroblasts (HELF, Fi301) and CMV AD169 were kept in the Institute of Virology, Humboldt University of Berlin. The mutant CMV strain RV-TB40E-4 was kindly provided by G. Hahn, München, Germany.
Peptides
IE-1 peptides (Swiss-Prot accession numbers P06725 and P13202) (15 amino acids in length, 11 overlaps) were obtained from Jerini Biotools (Berlin, Germany). All peptides were analyzed by MALDI-TOF mass spectroscopy, and purity was 70% minimum by HPLC. The peptides were dissolved in DMSO at a concentration of 8 mg/mL and stored at –80°C.
Fibroblasts and preparation of virus stocks
Human embryonic lung fibroblasts (HELF, Fi301) were grown as a monolayer in Eagle's minimal essential medium (EMEM) containing 25 mM Hepes supplemented with 0.05 mg/mL gentamicin, 2 mM N-acetyl-L-alanylglutamine (all from Cambrex, Verviers, Belgium), and 10% fetal calf serum (FCS; Biochrom, Berlin, Germany). Medium as well as FCS are certified for low endotoxin and tested free of mycoplasm. For virus stock preparation a confluent monolayer of HELF was infected with CMV strain AD169 or the mutant virus RV-TB40E-4 at a multiplicity of infection (moi) of 0.01 to 0.1. Virus RV-TB40E-4 was kindly provided by G. Hahn, Munich, Germany, and was deleted for the immune evasion genes US2-6 and US11. For preparation of virus stocks, virus was harvested from the supernatant of the infected cell culture after complete cytopathogenic effect (CPE). Cell debris was eliminated by low-speed centrifugation for 20 min at 3500 rpm (Labofuge 400; Heraeus, Hanau, Germany). If necessary, virus was concentrated by ultracentrifugation (2 h, 21,000 rpm, 4°C; Beckmann Ultracentrifuge, Munich, Germany) and resuspended in fresh medium. Virus stocks were aliquotted and stored in liquid nitrogen. Virus titers were determined on HELF using a standard plaque assay.
Blood collection and preparation of PBMCs
Blood was collected from anti-CMV IgG-seropositive healthy donors. HLA genotyping (4-letter code) for all donors was obtained from the local blood bank. PBMCs were prepared from 50–80 mL of freshly drawn citrated venous blood by standard Ficoll-Paque density gradient centrifugation (Pharmacia, Uppsala, Sweden). The cells were washed twice with sterile phosphate-buffered saline (PBS) and resuspended in complete RPMI-1640 media (Biochrom) containing 2 mmol/L L-glutamine (Biochrom), 10% (vol/vol) heat-inactivated FCS (Biochrom), 100 IU/mL penicillin, and 100 μg/mL streptomycin (Biochrom). Cell concentration in the final suspension was between 1.0 and 2.0×106 cells/mL.
Infection of fibroblasts with CMV
HELF were grown as a monolayer in 24-well plates in Dulbecco's modified Eagle's medium (DMEM) containing 2.0% (v/v) FCS. Generally, confluent monolayers were infected at an moi of 1, and virus adsorption lasted for 1 h at 37°C. After absorption, free virus was removed and the cells were overlaid with fresh medium. In cultures treated with inhibitors, MG132 (Calbiochem, Darmstadt, Germany), or GCV was added at this time. All samples were cultured in a standard incubator at 37°C for 72 h. Before incubation with PBMCs, the supernatant was removed and HELF was washed. MG132 (Z-Leu-Leu-Leu-CHO) was dissolved in ethanol at a concentration of 5 mg/mL (stock) and stored at −20°C. To obtain working solution, stock solution was freshly diluted (1:100) in MEM medium.
Stimulation of PBMCs with fibroblasts
Two million PBMCs were incubated with CMV-infected HELF or peptide-loaded HELF in 2 mL RPMI 1640 medium supplemented with 10% FCS for 6 h at 37°C, or with medium alone as a negative control. For measuring intracellular cytokines, the assay was performed using brefeldin A (BFA; Sigma-Aldrich, St. Louis, MO) as a golgi-transport inhibitor at a final concentration of 10 μg/mL added 2 h after the beginning of the incubation. Following an additional 4 h of incubation, PBMCs were harvested, washed with FACS washing buffer (FB), and fixed with 1 mL diluted BD lysing solution for 10 min at room temperature in the dark. After repeated washing with wash buffer, 1 mL of diluted BD permeabilizing solution 2 (PB) was added and the cells were incubated for 10 min at room temperature in the dark. Finally, the cells were spun down and stained with anti-CD3-PerCP (BD Diagnostics, Heidelberg, Germany), anti-CD8-APC (BD Diagnostics), and anti-human interferon-γ (IFN-γ) FITC (BD Diagnostics) for 30 min at 4°C. After washing, the cells were resuspended in FB.
Staining with antibody against the intracellular cytokine IFN-γ
Following incubation, the cells were washed with 3 mL ice-cold PBS, spun down (430 g, 8 min, 4°C), resuspended in 3 mL of PBS containing 2 mM EDTA (Sigma-Aldrich), incubated for 10 min at 37°C (in a water bath), spun down again (430 g, 8 min, 4°C), and vortexed for 30 sec. The cells were then washed with FB, and the supernatant was carefully removed. Fixation and permeabilization were performed in two steps using commercial BD lysing solution (BD Diagnostics), and permeabilization solution 2 (BD Diagnostics), respectively. After careful re-suspension of the cell-pellets, 1 mL of diluted BD lysing solution was added, and the tubes were incubated for 10 min at room temperature in the dark. The cells were spun down (340 g, 8 min, 4°C), the supernatant was removed by decanting, and the pellet was resuspended. Then 1 mL of PB was added to each tube. Following an additional wash step (see above), combined surface and intracellular staining with monoclonal antibodies was carried out for 30 min at 4°C in the dark. Staining was performed in a final volume of 100 μL. All antibodies were titered beforehand to achieve optimum results. Following a final wash step all samples were analyzed on a BD LSR II (BD Biosciences, San Jose, CA), or a FACScalibur (BD Biosciences) flow-cytometer using the FACS-Diva or Cell-Quest software packages, respectively.
Flow cytometric analysis
Data acquisition was carried out on a BD FACScalibur, or an LSRII flow cytometer using CellQuest or FACS-Diva software, respectively. To analyze lymphocytes, a minimum of 250,000 lymphocytes (live count gate) were acquired for each sample. For data analysis IFN-γ+/CD3+/CD8+ T cells were gated, and the percentages of the reference population (total CD8+ T cells) were determined. To test fibroblasts, 100,000 fibroblasts (live count gate) were acquired for each sample. For data analysis HLA-A*0201+ fibroblasts stained with phycoerythrin (PE)-labeled anti-HLA-A02 (BD Diagnostics) were gated, and the percentages of the reference population (total live fibroblasts) were determined.
Peptide stimulation, IFN-γ selection, and peptide-specific T-cell expansion
Peripheral blood samples were donated by healthy CMV IgG-seropositive adults with a certain HLA-type. Then 1×108 PBMCs were diluted at 1×107/mL CTL medium (RPMI 1640; Gibco BRL, Eggenstein, Germany; plus 10% autologous heat-inactivated serum, and 500 IU/mL penicillin G and 500 μg/mL streptomycin). HLA-restricted synthetic peptides (IE-1 peptide pool) of CMV were added to a final concentration of 1 μg/mL, and incubated for 6 h in a 37°C humidified incubator. Magnetic enrichment of cytokine-secreting cells was performed similarly according to the manufacturer's instructions (Miltenyi Biotec, Bergisch Gladbach, Germany). The cells were labeled with anti-IFN-γ monoclonal antibody conjugated to the leukocyte-specific (CD45) antibody (Miltenyi Biotec), diluted at 1×106/mL in CTL medium, and incubated for the IFN-γ capturing period (45 min). Thereafter the cells were magnetically labeled with anti-IFN-γ microbeads (Miltenyi Biotec), incubated for 15 min at 4°C, and washed with PBS (containing 1% autologous serum and 2 mM EDTA). The cells were resuspended in 2 mL complete medium, and then selected with a MACS device (Miltenyi Biotech). Isolated cells were washed twice and seeded in a 96-well plate (2×104, 5×104 per well), together with irradiated autologous PBMCs (5×105 per well, 30 Gy) in IL-2-supplemented complete medium (50–100 IU/mL proleukin; Chiron, Emeryville, CA). For expansion, T cells were stimulated weekly with autologous/allogeneic PBMCs at a ratio of 1:4 in rIL-2-supplemented medium.
Cytotoxicity assay
For cytotoxicity assay, a modified Calcein-AM release assay was used (22). Target cells (HELF and CMV-infected HELF) were labeled with Calcein-AM (15 mM; Molecular Probes, Eugene, Oregon), according to the manufacturer's instructions and seeded in 96-well plates (4×103/well). IE-1-specific T cells were propagated with different target cells for 4 h at effector-to-target ratios of 40:1, 20:1, 10:1, and 5:1. Triplicates were used for all tested samples, including controls with medium for spontaneous and Triton-X for maximum release. Lytic activity was calculated as follows: % specific cytotoxicity=(sample release – spontaneous release)/(maximum release – spontaneous release)×100.
Statistical analyses
Comparisons between different groups were done by pairs-samples t-test using SPSS 11.5 for Windows software. Significance was set at p<0.05.
Results and Discussion
Response of different donors to CMV AD169-infected fibroblasts
The present study aimed to investigate the effect of proteasome inhibitor on the CMV-specific CD8+ T-cell immune response. In order to select one donor who responded to CMV-AD169-infected HELF, we compared class I MHC type of 50 anti-CMV IgG seropositive donors, and two negative healthy volunteers with that of HELF (i.e,. HLA-A*0201, HLA-A*24, HLA-B*0702, and HLA-B*27). Among them, 12 donors whose MHC-I types matched one of the four alleles of HELF were selected. Ten were CMV seropositive (D1-10), and two were CMV seronegative (D11 and 12; Table 1). Only D3 and D9 responded to AD169-infected HELF.
a: anti-HCMV IgG.
b: uninfected human embryo lung fibroblast.
c: HCMV-infected human embryo lung fibroblast.
Proteasome inhibitor MG132 reduced cytokine secretion from CD8+ T-cells in response to AD169-infected HELF
To investigate the effect of MG132 and GCV on Ag presentation of CMV-infected HELF, we selected donor 3 as a responder to CMV-infected fibroblasts. The proportion of CD8+ T cells producing IFN-γ in response to infected fibroblasts was reduced under the influence of MG132 in a dose-dependent manner, in the range from 0.05 μM and 0.5 μM in comparison with the response to untreated infected fibroblasts (control) (Fig. 1). At the concentration of 0.5 μM a marked reduction was observed in three independent experiments. Namely, the percentage of CD8+ T cells secreting IFN-γ was reduced by about 49% at 24 h p.i. (p=0.025; Fig. 1A and D), by 40% at 48 h p.i. (p=0.099; Fig. 1B and D), and by 29% at 72 h p.i. (p=0.024; Fig. 1C and D), respectively. However, the amount of CD8+ T cells producing IFN-γ was not influenced by GCV (p>0.1; Fig. 1D). Thus MG132 at the concentration of 0.5 μM, which blocked CMV replication completely and reduced IE and pp65 protein expression, showed a significant suppression of the specific CD8+ T-cell response to CMV-infected fibroblasts. This is in agreement with the latest findings from Blanco et al., that PBMCs stimulated with allogeneic cells and treated with bortezomib markedly reduced their ability to produce IFN-γ when re-stimulated with the same allogeneic cells (23), although the reduction of IFN-γ-secreting CD8+ T cells dropped over the time period, which may indicate that more late proteins were expressed, or that the function of MG132 was no longer active/available at later time points, while the CMV replication inhibitor GCV (10 μM) had no such effect.
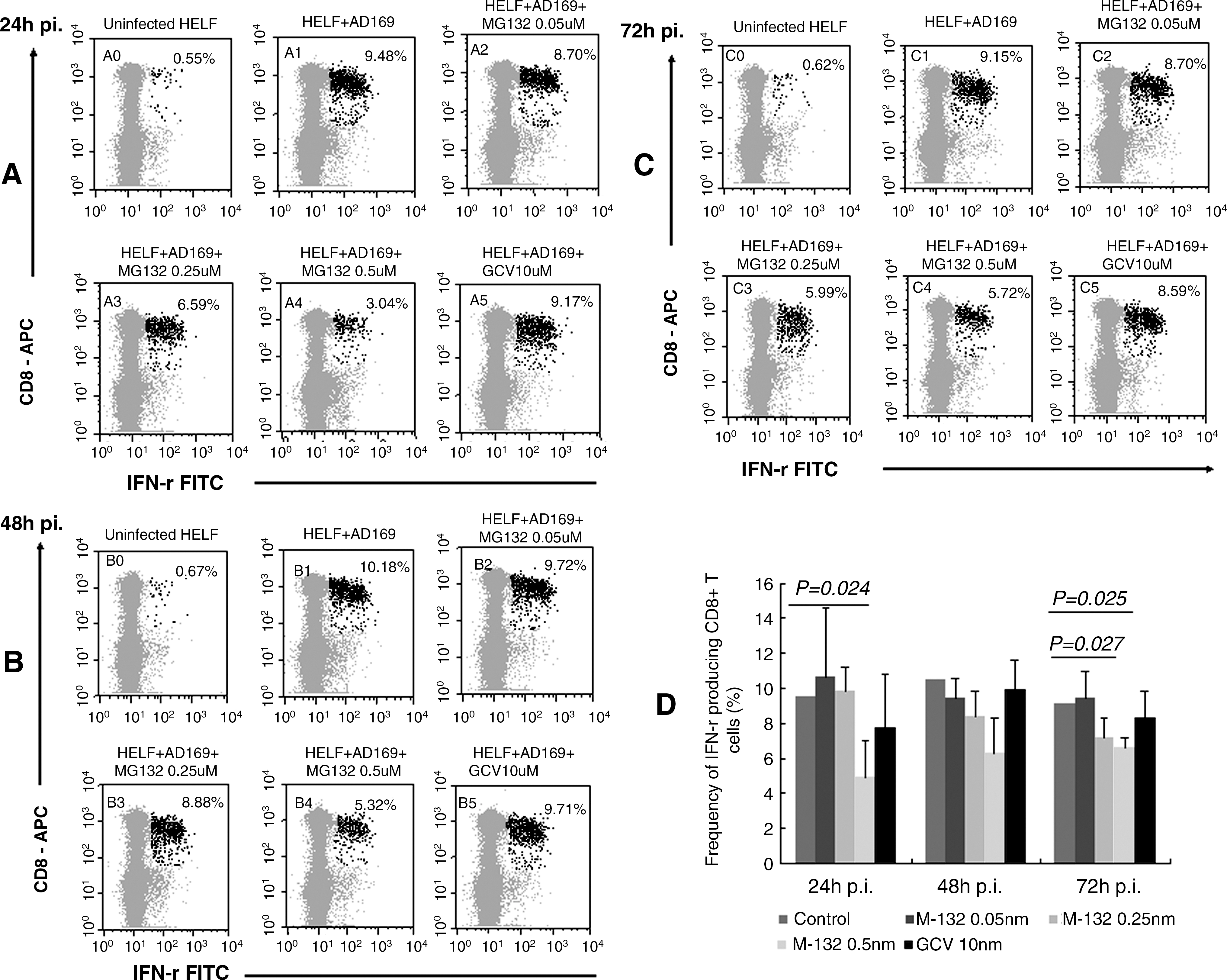
Effect of MG132 and GCV on the secretion of IFN-γ from CD8+ T cells in response to AD169-infected HELF. HELF were infected with CMV AD169 at an moi of 1, and incubated with different dosages of MG132 or GCV (10 μM) for 24 h, 48 h, or 72 h. Untreated AD169-infected HELF served as control. The red dots in
The maximum inhibition at 0.5 μM MG132 (which caused nearly complete inhibition of viral replication and spread) was only up to 50% in IFN-γ secretion, indicating that antigen-presenting machinery of the treated cells was incompletely blocked at this non-toxic concentration. The partial inhibition of Ag presentation and the CD8+ T-cell response may arise from the fact that MG132 blocks generation of only one or a few of the specific epitopes recognized by the donor, while generation of the others remained unchanged. This is supported by the observation that the level of inhibition did change over the time of infection. Analysis of other peptides should be done to verify the specific effects of MG132 on CMV.
Effect of MG132 on the expression of MHC class I molecules on HELF
To determine whether the inhibitory effect of MG132 was due to suppression of MHC I molecule expression, we tested class I MHC expression on HELF in the absence or presence of 0.5 μM MG132. As shown in Fig. 2, MHC I expression on MG132-treated uninfected HELF was reduced by 64.66% (Fig. 2A) compared to untreated uninfected cells, and its expression on AD169-infected cells was decreased by 89.57% of uninfected cells (Fig. 2B). However, MHC I expression on AD169-infected HELF post-MG132 treatment was increased by 15.08% compared to untreated infected cells (Fig. 2B), and MHC I expression on RV-TB40E-4-infected HELF treated with MG132 was decreased by 43.79% compared to untreated infected cells (Fig. 2C). As RV-TB40E-4 does not contain US2-6/US11, the MHC I downregulatory immune evasion genes (24) and ubiquitination are crucial for CMV US11-mediated dislocation of MHC class I molecules from the endoplasmic reticulum to the cytosol (25). These results imply that MG132 is able to reduce the viral MHC-downregulatory effect via inhibition of viral gene expression. Previous studies demonstrated the antiviral effect of MG132, and showed that viruses were interfered with at various steps of viral replication (26,27). Therefore, we deduced that inhibition of IFN-γ secretion from CD8+ T cells in response to AD169-infected cells in the presence of MG132 was not due to suppression of MHC I expression, but might be attributable to a reduction in antigen presentation of immunodominant CMV epitopes. The findings in the treatment of cancers have demonstrated that proteasome inhibitors decrease the presentation of antigenic peptides to reduce tumor cell recognition by CTLs (28).
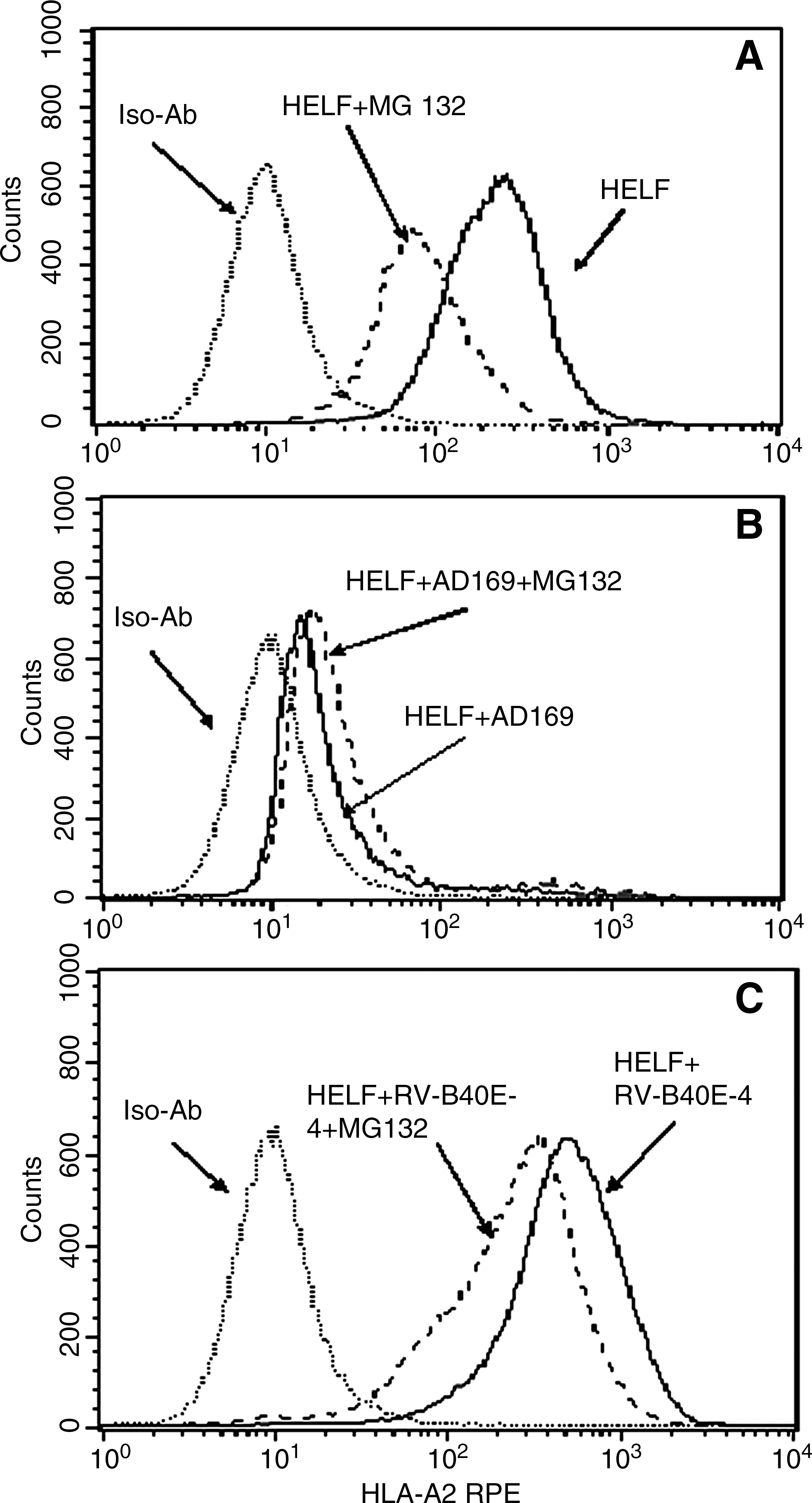
Influence of MG132 on the expression of MHC I molecules on HELF. HELF were infected with AD169 or RV-TB40E-4 at an moi of 1 for 72 h in the absence or presence of 0.5 μM MG132, and tested for fluorescence intensity of anti-HLA-A*0201-PE with the FACScalibur. Iso-Ab is the abbreviation for the isotype antibody. HELF+MG132 is the abbreviation for HELF treated with 0.5 μM MG132. HELF+AD169/RV-TB40E-4 is the abbreviation for HELF infected with CMV AD169 or RV-TB40E-4, respectively. HELF+AD169/RV-TB40E-4+MG132 is the abbreviation for CMV AD169 or RV-TB40E-4-infected HELF treated with 0.5 μM MG132, respectively. Cell counts are indicated on the y-axis. Log fluorescence intensity of anti-HLA-A*0201-PE is indicated on the x-axis.
MG132 decreased the cytotoxicity of an IE-1-specific T-cell line against CMV-infected HELF
Further, we tested the cytotoxicity of an IE-1-specific T-cell line against CMV-infected HELF in the presence of MG132 or GCV. As demonstrated in Fig. 3, killing was 52% in untreated infected HELF (control), and killing was only 10% in MG132-treated infected HELF at a ratio of 40:1. Thus, MG132 caused a significant reduction of the lysis of CMV-infected HELF, consistent with recent findings from Basler et al. (29), that blockage of LMP7 by proteasome inhibitor PR-957 greatly reduced pathological symptoms of dextran sulfate sodium-induced colitis, and production of various cytokines in PR-957-treated mice was suppressed, resulting in reduced inflammation and tissue destruction (30). In contrast, GCV had no influence on T-cell-specific killing of target cells (GCV versus control=55% versus 52%).
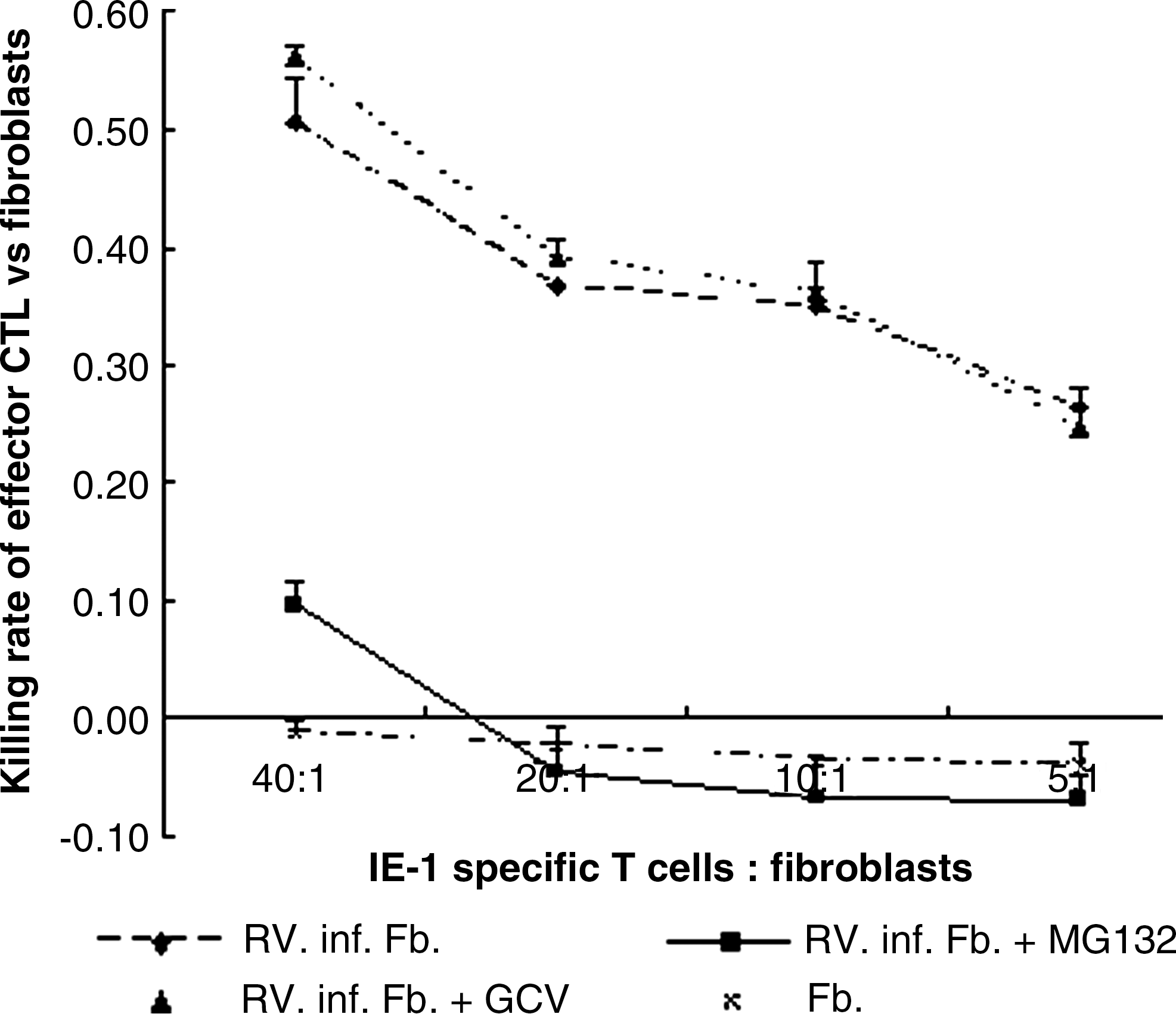
Cytotoxicity of the IE-1-specific T cells against CMV-infected HELF treated with MG132 or GCV. Fibroblasts were infected with RV-TB40E-4 at an moi of 1 and incubated in the absence or presence of MG132 or GCV for 48 h. After that the cells were harvested and labeled with Calcein-AM for 30 min. IE-1-specific T cells were added to fibroblasts at different ratios, which are indicated on the x-axis, and incubated for 4 h. Supernatant was collected to measure the fluorescence intensity of Calein-AM released from the killed fibroblasts. Fb is the abbreviation for fibroblasts. Rv.inf.Fb+ MG132/GCV are the abbreviations for CMV RV-TB40E-4-infected fibroblasts treated with 0.5 μM MG132 or 10 μM GCV. The killing rate and the effector T cell-to-target fibroblast ratio are indicated on the y-axis and x-axis, respectively. One representative experiment of two is shown.
By comparing the effects of MG132 with GCV it becomes obvious that GCV has no impact on the CMV-specific CD8+ T-cell response, at least for the observed donor. IE-1 and pp65 as the most immunodominant proteins were present as early as the immediate stage, and GCV has no suppressive effect on their expression, indicating that GCV is not able to prevent CMV-associated immunopathy.
Conclusions
As CMV reactivation and its related immunopathy tend to result in graft malfunction or even rejection, it may be feasible to suppress CMV-related immunopathogenesis via altering epitope generation by proteasome inhibitors. Our data suggest that proteasome inhibitor MG132 at a non-toxic concentration is able to inhibit the CMV-specific CD8+ T-cell immune response in a dose-dependent manner. These findings may have important implications in relieving CMV-associated immunopathy for transplantation, as well as for treating autoimmune disease.
Footnotes
Acknowledgments
We are grateful to the China Scholarship Council for their financial support. We sincerely thank Dr. Markus Hammer, Dr. Sebastian Pfeiffer, Dr. Lydia Tesfa, and Mrs. Rita Schilf, for their technical support. This paper is in memory of Dr. Susan Prosch for her serious attitude toward scientific research and her profound knowledge.
Author Disclosure Statement
No competing financial interests exist.