
Abstract
Reactive oxygen species (ROS) are produced predominantly by phagocytic cells in response to microbial infections. When produced at optimal levels ROS have potent antimicrobial properties. However, excessive production of ROS induces apoptosis/necrosis of infected as well as bystander cells, resulting in inflammatory pathology. Previously we showed that vaccination of pigs with a modified live porcine reproductive and respiratory syndrome virus vaccine (PRRS-MLV) administered intranasally with a potent mucosal adjuvant M. tuberculosis whole-cell lysate (Mtb WCL) induces protective immunity against PRRSV challenge. In this study, using bronchoalveolar lavage fluid cells and peripheral blood mononuclear cells harvested from that study were quantified for the levels of ROS produced. Our results indicated that in vaccinated pigs, levels of ROS were lower compared to unvaccinated PRRSV-challenged pigs. In unvaccinated but PRRSV-challenged pigs, the higher ROS production was associated with increased inflammatory lung pathology. In conclusion, our results suggest that intranasal immunization using PRRS-MLV along with a potent mucosal adjuvant protects pigs against both homologous and virulent heterologous PRRSV challenge, which was associated with reduced ROS production and reduced lung pathology compared to control virus-challenged pigs.
Introduction
ROS refer to differential oxidation and excitation states of molecular oxygen, such as superoxide, singlet oxygen, hydroxyl radical, hydrogen peroxide, peroxide, and hypochlorite. ROS are an integral part of cell-signaling; however, at higher concentrations they cause severe tissue damage (7). They play an important role in physiological events like host defense, mitogenesis, hormone biosynthesis, apoptosis, and fertilization. During active infection, ROS are produced by phagocytic cells in large quantities at the local mucosal infection sites. Several enzymatic activities are involved in the production of ROS, like the membrane-bound nicotinamide adenine dinucleotide phosphate (NADPH) oxidase (8), and the xanthine oxidase in the mitochondria of phagocytic cells (9,10), that converts singlet oxygen to superoxide anions (O2 –). Furthermore, superoxides are converted to hydrogen peroxide (H2O2) by the action of superoxide dismutase. In neutrophils, H2O2 can further originate hydroxyl radicals (OH−), and/or it can combine with chloride ions (Cl−) to generate hypochlorite (HCLO−), mediated by myeloperoxidase. ROS are also produced at lower levels by non-phagocytic cells (11).
The steady-state production of pro-oxidants in cells and organs is balanced by an equal rate of consumption by antioxidants. Oxidative stress results from the imbalance in equilibrium between pro-oxidants and antioxidants, and unfortunately it has always been in favor of pro-oxidants (12). Extreme non-physiological concentrations of oxidants cause tissue destruction. Oxidative tissue damage caused by ROS has been detected in a large number of infectious settings, including viral pneumonitis (13 –15). Peroxynitrate (ONOO−) is a product of the reaction between nitric oxide (NO.) and O2 − in specific phagocytes. Influenza virus-induced lung pathology is mediated by the production of ONOO− in alveolar macrophages (16 –18). Pneumonia in Hantavirus cardiopulmonary syndrome (HCPS) (19), and in herpes simplex virus (HSV-1), is mediated through ROS (20). ROS are among the most important mediators of diffuse PRRSV-mediated pathology in the lungs and lymph nodes of infected pigs (5). In this study we quantified ROS production in immune cells present in the lungs and blood of PRRSV-challenged unvaccinated or mucosally-vaccinated (PRRS-MLV+Mtb WCL) pigs. Our results identified the reduced production of ROS in mucosally-vaccinated and PRRSV-challenged pigs, which supported our previously reported results of protective immunity associated with reduced lung pathology compared to control virus-challenged pigs.
Materials and Methods
Pigs and inoculations
Conventional Large White-Duroc crossbred specific-pathogen-free piglets weaned at 3–4 wk of age were transported from the swine herd free from PRRSV, porcine respiratory corona virus, transmissible gastroenteritis virus, and porcine circo virus 2, to the animal facilities of the Food Animal Health Research Program at the Ohio Agricultural Research and Development Center, Wooster, Ohio. The piglets were bled on arrival, and their sera were tested to confirm the absence of PRRSV antibodies by ELISA. The animals were maintained in our large animal BSL2 facility. Throughout the duration of the study, all animals received food and water ad libitum. All inoculations including PRRSV challenges were administered intranasally. All pigs were maintained, samples were collected, and the animals were euthanized per the protocol approved by the Institutional Animal Care and Use Committee (IACUC) and Institutional Biosafety Committee (IBC) of The Ohio State University. Forty pigs were allocated to one of five groups: group 1, mock pigs (n=4) inoculated with vehicle (normal saline) and DMEM; for the other four groups (n=9 per group), groups 2 and 3 were unvaccinated, and groups 4 and 5 were vaccinated (PRRSV-MLV+Mtb WCL). Groups 2 and 4 were challenged with wild-type homologous PRRSV VR2332 (2×106 TCID50 per pig), and groups 3 and 5 were challenged with virulent heterologous PRRSV MN184 (21) (1×106 TCID50 per pig) in 2 mL on day 21 post-immunization (DPI). Three pigs each from groups 2 to 5 were euthanized at 15, 30, and 60 d post-challenge (DPC). Mock inoculated pigs (n=4) were euthanized separately prior to euthanizing any challenged animals.
Collection of blood and bronchoalveolar lavage fluid samples and isolation of immune cells
Peripheral blood mononuclear cells (PBMCs) were isolated from blood collected in acid citrate dextrose solution from euthanized pigs and processed as previously described (4,22). Bronchoalveolar lavage fluid mononuclear cells (BAL-MNC cells) were isolated as previously described (4,23).
Determination of ROS concentration by colorimetric assay
A colorimetric assay based on the ability of ROS to reduce nitroblue tetrazolium (NBT) to a blue-colored formazan was performed as previously described (24,25). The quantity of ROS produced is directly proportional to the optical density (OD) value measured at 650 nm. Briefly, BAL cells (0.5×106) and PBMCs (1×106) were suspended in enriched RPMI-1640 (containing 10%FBS, gentamicin [100 μg/mL], ampicilin [20 μg/mL], 20 mM HEPES, 2 mM l-glutamine, 0.1 mM non-essential amino acids, 1 mM sodium pyruvate, and 50 nM 2-ME), and restimulated in the presence or absence of killed crude PRRSV antigens (Ags) (50 μg/mL) at 37°C for 24 h to induce PRRSV-specific response. The cells were washed twice, suspended in PBS (pH 7.2), and seeded in 48-well tissue-culture plates in quadruplicate. Duplicate wells were treated with 0.1% NBT, and as a negative control cells treated with 0.1% NBT and 10 μM of diphenlyeneiodonium (DPI), a potent inhibitor of NADPH oxidase, were included (26,27). Cells treated with phorbol 12-myristate 13-acetate (PMA; 600 ng/mL) and 0.1% NBT were included as a positive control. Cells unstimulated with PRRSV Ags were included as a blank, and the plates were incubated for 45 min in a CO2 incubator at 37°C. The reaction was stopped using ice-cold PBS (pH 7.2) (500 μL/well), and washed three times in PBS by centrifugation at 2100×g for 3 min. Subsequently, the cells were treated with methanol to remove excess un-utilized NBT, and air dried at room temperature. The reduction of NBT by ROS into blue-colored formazan was measured by solubilizing the cell membrane by addition of 2 M KOH (120 μL/well), and subsequently DMSO (140 μL/well) was added to dissolve the formazan, and the mixture was subjected to gentle shaking for 10 min at 37°C. Harvested supernatants were transferred into a 96-well plate and read in a microplate reader at OD650 nm.
Flow cytometric analysis to quantify the frequency of ROS-producing cells
The principle of this assay is based on the ability of ROS to reduce NBT to formazan, which in turn quenches the cell-bound immunofluorescent molecule, resulting in reduction in the frequency of fluorescence-positive cells. Therefore, the frequency of ROS-producing cells is inversely proportional to that of fluorescence-positive cells, as measured by flow cytometry. The assay was performed as previously described (28) with a few modifications. Briefly, fluorescein isothiocynate (FITC)-conjugated concanavalin A (FITC-ConA) (Sigma Chemical Co., St. Louis, MO) was used as a cell surface probe. ConA is a plant mitogen that binds to cell surface receptors containing mannose, which are present on many cell types including phagocytes (29). ConA does not influence the phagocytic activity and cell proliferation seen under the conditions described in our assay, such as dose, time of incubation, and temperature (30).
BAL cells and PBMCs suspended in enriched RPMI-1640 were either unstimulated or stimulated using killed PRRSV Ags (50 μg/mL) at 37°C for 24 h. The cells were washed twice in PBS and labeled with FITC-ConA (25 μg/mL) on ice for 30 min. The cells were washed once each using RPMI-1640 and PBS to remove the unbound FITC-ConA, resuspended in PBS, and seeded onto a 96-well plate. The cells were treated with 0.1% NBT in PBS, and a control well with unlabeled cells not treated with NBT was included in the assay. Cells stimulated with PMA (150 ng/mL) and 0.1% NBT in the presence or absence of DPI (10 μM) were included as controls, and the plates were incubated for 45 min at 37°C in a CO2 incubator. The reaction was stopped by addition of ice-cold PBS, then they were washed twice, and resuspended in ice-cold PBS and stored at 4°C until analyzed by flow cytometry. Unlabeled cells were used to set up the flow cytometer, and the frequency (percentage) of fluorescence-positive cells was recorded from a total of 20,000 acquired events using the flow cytometer (FACS Aria II; BD Biosciences, San Jose, CA), as previously described (4,31). Data were analyzed using FlowJo software (Tree Star, Inc., Ashland, OR).
Statistical analysis
All the data were expressed as the mean±standard error of the mean (SEM) of three pigs. Statistical analyses were performed by a non-parametric Wilcoxon t-test (SAS software; SAS Institute Inc., Cary, NC). Statistical significance was set at p<0.05, and only significant data are indicated in the figures.
Results and Discussion
PRRSV replicates in the lungs of infected pigs, and the virus-specific immune responses are suppressed and/or delayed. Also, PRRSV-induced immune-mediated inflammatory responses result in patches of dark brown consolidation in the lungs. Recently we evaluated the efficacy of PRRSV-MLV administered with an adjuvant Mtb WCL intranasally to pigs. Our results detected enhanced immune correlates of protection associated with significantly reduced gross lung pathology and protective immunity to a virulent heterologous PRRSV MN184 challenge, and also to a homologous VR2332 challenge (31) (unpublished data). In contrast, unvaccinated PRRSV-challenged pig lungs had significantly increased gross lung consolidation (31). Pathological changes in the lungs of PRRSV-infected pigs are due to necrosis of virus-infected and bystander cells mediated by aberrant host immune responses (5,32,33). In the lungs of PRRSV-infected pigs, viral replication, generation of mutagenic viral quasispecies, and the cytokine storm caused by excess secretion of proinflammatory cytokines, result in ROS-mediated oxidative stress and lung pathology (5,6,34).
Consistent with previous studies, BAL mononuclear cells (BAL-MNC) of unvaccinated PRRSV VR2332-challenged pigs produced a significant increase in the amount of ROS compared to vaccinated pigs at 30 and 60 DPC (Fig. 1A and Table 1A). In PBMCs a comparable increase in ROS production was also detected in unvaccinated compared to vaccinated pigs (Fig. 1B and Table 1B). Similarly, in unvaccinated compared to vaccinated and virulent heterologous PRRSV MN184-challenged pig BAL cells, a significantly increased production of ROS at an earlier time point (DPC 15) was detected (Fig. 1C and Table 1A). A significant increase in ROS production was also detected in PBMCs at DPC 15 and 60 (Fig. 1D and Table 1B). These results correlate with the virulent nature of PRRSV MN184 associated with the severe gross lung consolidation observed at DPC 15 (31).
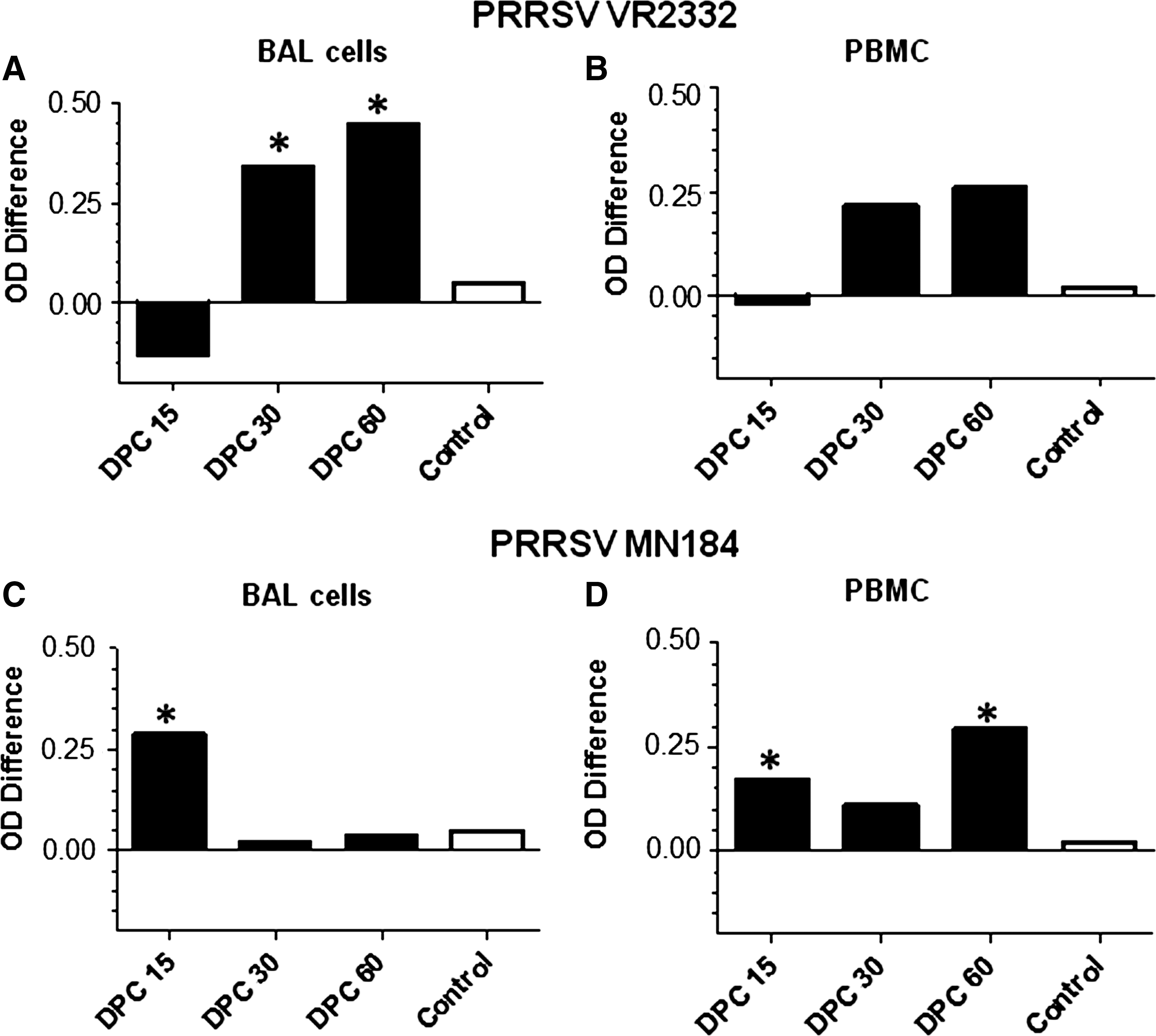
Enhanced ROS production in unvaccinated PRRSV-challenged pigs was detected by a colorimetric assay. Groups of three pigs were unvaccinated or vaccinated (PRRS-MLV+Mtb WCL) intranasally, and challenged using either PRRSV VR2332 (
Each number is an average OD value from three pigs±SEM. The higher the OD650nm value, the higher the production of ROS. Asterisks indicate statistically significant differences (p<0.05) between vaccinated (Vac) and unvaccinated (Unvac) pig groups.
Vac or Unvac+VR2332, and Vac or Unvac+MN184, pigs vaccinated or unvaccinated with PRRSV-MLV+Mtb WCL, and challenged using PRRSV VR2332 or MN184; OD, optical density; DPC, days post-challenge; PBMC, peripheral blood mononuclear cell; NBT, nitroblue tetrazolium; PMA, phorbol 12-myristate 13-acetate; DPI, diphenlyeneiodonium; SEM, standard error of the mean; ROS, reactive oxygen species; PRRSV, porcine reproductive and respiratory syndrome virus; BAL-MNC, bronchoalveolar lavage fluid mononuclear cell; Mtb WCL, M. tuberculosis whole-cell lysate.
The reduction of NBT to formazan by ROS produced by pig immune cells was specific, as the DPI treatment inhibited the mitogen PMA-induced ROS production (26,27), or PRRSV-induced ROS (Fig. 2A). Our results are consistent with the quantitatively increased ROS production detected in the lungs of unvaccinated PRRSV-challenged pigs (32). Flow cytometric analysis was used to measure the frequency and function of ROS-producing cells (28), based on the principle that ROS mediate quenching of the cell surface fluorescent molecule, here FITC, tagged to ConA. Quenching is not directly proportional to the amount of ROS produced, mainly due to the difference in the quantity of ROS produced by each type of phagocytic cell. For example, neutrophils produce 2–3 times more ROS than macrophages (13,35). In flow cytometric analysis, only the frequency of cells producing the ROS is determined, and it is not possible to quantify the ROS produced by each cell, while in a colorimetric assay, total or pooled ROS produced by a given number of cells can be determined (36,37). Since BAL cells and PBMCs contain varying proportions of different phagocytic cell types, results from the two assays are not comparable, but they are complementary to each other in drawing a meaningful conclusion about ROS production. In other words, flow cytometric analysis determines a qualitative estimation, and colorimetric analysis estimates a quantitative production, of ROS.
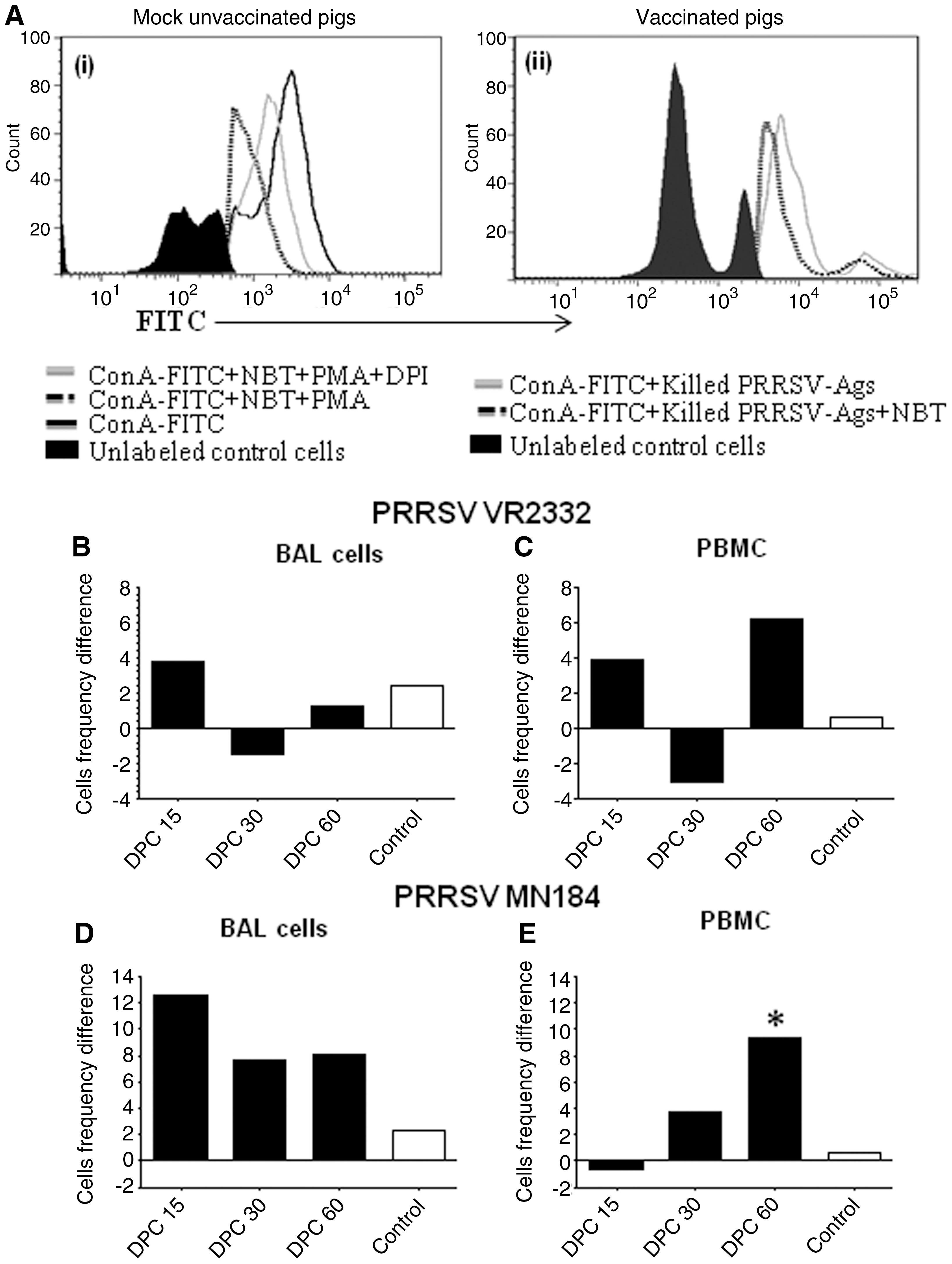
Quantification of the frequency of PRRSV-induced ROS-producing cells by flow cytometry. Standardization of the detection of ROS-producing pig immune cells: (
An experiment showing the standardization of the flow cytometric assay to quantify ROS production by pig immune cells was performed (Fig. 2A). Treatment of BAL cells with PMA results in production of ROS, which reduced the frequency of fluorescence-positive cells, and this was rescued by DPI treatment (Fig. 2Ai). Furthermore, to demonstrate the PRRSV-induced ROS production, treatment of BAL cells obtained from PRRSV-challenged vaccinated pigs restimulated in vitro with killed PRRSV Ags were treated with NBT. A reduction in the frequency of fluorescence-positive cells was detected, indicating reduction of NBT by ROS to formazan, which in turn quenched the immunofluorescent molecules present on the cell surface of labeled cells (Fig. 2Aii).
In PRRSV VR2332-challenged unvaccinated pig PBMCs and BAL cells compared to their respective vaccinated pig cells, there was an increase in the frequency of ROS-producing cells at DPC 15 and 60 (but not at DPC 30) (Fig. 2B and C and Table 2A and B). In unvaccinated, virulent PRRSV MN184-challenged pigs an increased frequency of ROS-producing cells was detected in BAL cells at all DPC tested (Fig. 2D and Table 2A). In PBMCs a similar increase in ROS production was detected at DPC 30, and at DPC 60 the increase in ROS-producing cells was statistically significant (Fig. 2E and Table 2B). The control bars and values in the figures and tables signify that the assays used in this study are specific to ROS production by porcine phagocytic cells. The control bars/values shown (Figs. 1 and 2 and Tables 1 and 2) indicate the difference between the cells stimulated using PMA (positive control), and cells stimulated using PMA, but treated with DPI to inhibit ROS production (negative control). Performing both the assays simultaneously on the same set of pig immune cells helped us to interpret both the quantity of ROS and the frequency of immune cells producing ROS. Restimulation of immune cells using killed PRRSV Ags helped us to determine the PRRSV-mediated recall immune response, which we could quantify ex vivo.
Each number is an average frequency of fluorescent cells in percentages of three pigs±SEM. The higher the value, the lower the production of ROS. Asterisks indicate statistically significant differences (p<0.05) between the vaccinated and unvaccinated pig groups.
Vac or Unvac+VR2332, and Vac or Unvac+MN184, pigs vaccinated or unvaccinated with PRRSV-MLV+Mtb WCL, and challenged using PRRSV VR2332 or MN184; DPC, days post-challenge; PBMC, peripheral blood mononuclear cell; NBT, nitroblue tetrazolium; PMA, phorbol 12-myristate 13-acetate; DPI, diphenlyeneiodonium; SEM, standard error of the mean; ROS, reactive oxygen species; PRRSV, porcine reproductive and respiratory syndrome virus; BAL-MNC, bronchoalveolar lavage fluid mononuclear cell; Mtb WCL, M. tuberculosis whole-cell lysate.
At certain DPC there was no association between the quantity of ROS and the frequency of ROS-producing cells, suggesting that at times only a very few cells can secrete large amounts of ROS. Overall, PRRSV-challenged unvaccinated pig immune cells secreted more ROS, both in terms of quantity, and also by the frequency of the cells. Compared to the colorimetric assay, the cytometric analysis is more sensitive, objective, and functional, as it determines the frequency of ROS-producing cells (36,37), and this information is critical to estimate the extent and duration of the tissue damage mediated by ROS.
Results of our previously published study (31), and those of the current report, together indicate that mucosal immunization to PRRS induced adequate protective immunity against PRRSV challenge, which may be promoted by optimal levels of ROS production. Optimal production of ROS by immune cells is essential for antimicrobial activity (7,8,38,39), and excess production of ROS causes necrosis of infected cells, as well as apoptosis of bystander cells and phagocytes due to irreversible membrane lipid degradation and DNA fragmentation (40 –42). The severe lung pathology observed in unvaccinated, virulent PRRSV-challenged pigs (31) might be due to apoptosis of bystander cells, along with necrosis of virus-infected cells, consistent with the results of previous studies (5,43). The enhanced lung pathology observed in unvaccinated but virulent PRRSV-challenged pigs might be due to increased expression of the genes involved in excess ROS production (6,44). In conclusion, intranasal immunization of PRRS-MLV with a potent mucosal adjuvant Mtb WCL induces protective immunity, and importantly, limited the PRRSV-induced pulmonary pathology due to ROS.
Footnotes
Acknowledgments
This work was supported by a National Pork Board award and a USDA-AFRI PRRS CAP2 award to G.J.R. Salaries and research support were provided by the state and federal funds appropriated to the Ohio Agricultural Research and Development Center, The Ohio State University. We would like to thank Drs. Juliette Hanson, Mahesh Khatri, and Hadi Yassine, and Todd Root and Matthew Weeman, for their help with the animal studies. We also thank Drs. Eric Nelson, Michael Murtaugh, Mike Roof, Karen Dobos, and John T. Belisle for providing reagents.
Author Disclosure Statement
No competing financial interests exist.