
Abstract
Infectious bronchitis virus (IBV) is prevented primarily by the use of live attenuated vaccines, which are known to have a limited strain range of protection. Alternative vaccines against the emerging new virus strains can improve control of the disease. The aim of this study was to evaluate the immunogenic potential of two recombinant viral proteins, when administered by eyedrop, without the assistance of a vector. The recombinant S1 (rS1) and N (rN) proteins of the M41 strain expressed in E. coli were tested, and the live attenuated vaccine H120 was used as a positive control. Protection was evaluated by re-isolation of virus from tracheas of vaccinated chickens after challenge with strain M41. After three immunizations, rS1 glycoprotein induced 40% protection, while vaccination with rN provided no protection. Vaccination with rS1, rN, or H120 induced a cellular immune response as demonstrated by in vitro ChIFN-γ production by splenocytes of vaccinated birds. Vaccination with H120, and to a lesser extent rS1, induced HI and virus-specific IgG antibody production. These findings indicate that recombinant viral proteins administered through the mucosal route can evoke an immune response without the assistance of a vector.
Introduction
Prevention of the disease is achieved mainly through vaccination with live attenuated vaccines. Although some common antigenic types were found in different geographic regions, in most cases IBV strains within a geographic region are unique and distinct (6,13,20). IBV undergoes frequent changes in the viral genome, mainly in the S1 gene, as a result of point mutations or recombination, promoting the emergence of new antigenic variants (1,3,26). In most cases, vaccine strains are selected to represent the antigenic spectrum of isolates in a particular country or region. Yet attenuated vaccines are known to have a limited range of protection, confined in many cases to homologous strains, rendering vaccination only partially successful (23,27). The use of heterologous vaccine strains, either simultaneously or sequentially, has broadened the protection spectrum in some cases (10), but it is difficult to predict which combinations may confer the best protection. Although more virulent vaccine strains may have a broader range of protection, the use of such strains is not recommended because of the risk of a disease outbreak (11).
Development of an alternative to live attenuated IBV vaccines based on molecular technologies has been the endeavor of several studies in recent years. One approach was to use live vectors such as fowlpox virus or adenovirus to express IBV proteins (22,39). Although such vaccines could provide protection, the outcome of massive use of genetically engineered live vectors is still unknown. The use of viral DNA allows the creation of a multivalent vaccine without the potential risk of using live viruses, but it has to be applied by injection (14), therefore it has no practical benefits, especially in the broiler industry. Another approach makes use of reverse genetic techniques to create a virus with a manipulated genome such that it maintains its attenuated form, but can be adjusted to new virulent IBV strains (8). At present none of the above methods has been adopted for commercial use.
The goal of the present study was to express the structural proteins, recombinant S1 (rS1) and N (rN), in a relatively simple commercially available expression system, such as E. coli, and to test their immunogenic potential when applied by eyedrop, without the assistance of a live or inactivated vector.
Materials and Methods
Cloning the S1 and N genes
The S1 and N genes of the IBV M41 strain, a Massachusetts (Mass) serotype (GenBank accession no. AY561711 and EU116941, respectively) were amplified by RT-PCR as described previously (28), with the following primers: the S1 3′ homologous sequence (underlined), preceded by eight histidine codons (in bold letters), and EcoR I restriction site (5′-CGAGAATTCCTCGAG-CTAG
Expression, purification, and detection of the viral proteins
Bacteria carrying the rS1- and rN-containing plasmids with the correct sequences were grown on LB medium (10 g/L yeast extract, 5 g/L Bacto tryptone, and 10 g/L NaCl), supplemented with 50 mg/mL ampicillin, to a density of 0.5–0.7 OD 600 nm. Production of recombinant proteins was induced by adding 1 mM IPTG [isopropyl (b)-d-thiogalactoside] to the medium, and growing the bacteria for 3 h at 37°C. The cells were centrifuged, the pellet was dissolved in osmotic shock solution (20 mM Tris-HCl, pH 8.0, 50 mM NaCl, and 25 mM EDTA), and the cells were disrupted by three cycles of sonication followed by freezing in liquid nitrogen (−180°C) and thawing at 37°C. Screening for colonies expressing the proteins was performed by sodium dodecyl sulfate-polyacrylamide gel electrophoresis (SDS-PAGE) and Western blotting. The rS1 protein was purified by using the nickel chelate resin HIS-Select® iLAP® column (Sigma-Aldrich, St. Louis, MO), according to the manufacturer's instructions. A strong cation-exchange resin, Macro-Prep High S Support (BioRad, Hempstead, U.K.), in an AKTA prime device (Amersham Pharmacia Biotech, Uppsala, Sweden), was used for rN purification, according to the manufacturer's instructions. The eluted fractions were analyzed by 12% SDS-PAGE, and Western blotting.
Vaccination trials
Two vaccination trials were performed as follows: 1-day-old specific pathogen-free (SPF) Leghorn chicks (Charles River SPAFAS, North Franklin, CT) were assigned to groups of 14–16 birds per group, and placed in Horsfall-type positive pressure isolation units. The chickens were vaccinated by eyedrop or by subcutaneous injection at 1, 14, and 28 days of age, with 150 μg/bird of a viral protein. Chicks in the positive control group were vaccinated by eyedrop with the commercial Mass serotype live attenuated vaccine H120 (Phibro, Bet Shemesh, Israel), one recommended dose per bird, at 1 and 14 days of age. The negative control consisted of non-vaccinated birds. At 35 days of age, 4–6 birds from selected groups were sacrificed, the spleens were removed, and tracheal washes were sampled (16).
Trial 1
Three groups of 16 birds each were vaccinated by eyedrop with H120, rN, or rS1, as described above. A fourth group of 16 birds was vaccinated subcutaneously with a mixture of rN and rS1, emulsified with Freund's adjuvant. The first vaccination was at day 1 with Freund's complete adjuvant, and the second and third vaccinations with Freund's incomplete adjuvant. A fifth group remained non-vaccinated and served as negative control. Six birds from the non-vaccinated group or from the groups vaccinated with H120 or rN were sacrificed. Blood was drawn from all birds 2 wk after the first vaccination, 12 d after the second vaccination, and 3 wk after the last vaccination with the recombinant proteins (14, 40, and 49 d of age, respectively).
Trial 2
Three groups of 14 birds each were vaccinated by eyedrop with H120, rN, or rS1. A fourth group of 14 birds remained non-vaccinated and served as negative control. All birds were challenged with the live reference strain M41 by eyedrop at a viral load of 104.5 EID50 per bird. Tracheal swabbing was performed on all chickens at 5 d post-infection, and the samples were used for viral RNA extraction and IBV detection by real-time RT-PCR as described previously (29). Protection efficacy was evaluated by detection of the challenge virus in tracheas 5 d post-challenge. Blood was drawn from all birds at the day of challenge, and 9 d post-challenge, at 44 d of age.
Humoral immune response
The potential of the vaccines to induce antibody production was assessed in sera and in tracheal washes, testing each sample individually. Sera samples were tested with a commercial IBV ELISA kit (FlockCheck; IDEXX Laboratories, Inc, Westbrook, ME), according to the manufacturer's instructions. In addition, specific IgA and IgG polyclonal antibodies in blood and tracheal washes, produced in response to rN, rS1, or H120 vaccinations, were tested by ELISA, using inactivated M41 as antigen, as follows: sera and tracheal washes were diluted in PBS at 1:200 and 1:10 dilutions, respectively. All samples and reagents were added at 100 μL/well. For the detection of IgA antibodies, the plates were coated with 10 μg/mL goat anti-chicken IgA (Bethyl, Montgomery, TX) in coating buffer (carbonate-bicarbonate buffer, pH 9.6), and incubated overnight at 4°C. After three washes with washing buffer (PBS+0.01% [v:v] Tween 20), sera or tracheal washing samples were dispensed onto the coated plates, and incubated for 1 h at room temperature. Following three washes, inactivated M41 was added at a concentration of 100 μg/mL protein, and incubated for 1 h at room temperature. After three washes, infectious bronchitis-Mass chicken antiserum (Charles River SPAFAS) was added at 1:500 dilution, and incubated for 1 h at room temperature. After three washes, 20 ng/mL HRP goat anti-chicken IgG-Fc conjugate (Bethyl) was added, and the plates were incubated for 40 min at room temperature. After three washes TMB substrate was added (KPL, Camarillo, CA), and after development of the reaction, the OD was measured at 620 nm. For the detection of IgG antibodies, the plates were coated with inactivated M41 at a concentration of 100 μg/mL protein, followed by sera or tracheal washing sample addition, followed by HRP goat anti-chicken IgG-FC conjugate and TMB, as specified above. Hemagglutination inhibition (HI) antibodies were tested using 8 units of a Mass type IBV antigen (Charles River) (36).
Cell-mediated immune response
Spleens were sampled from 35-day-old chickens, and splenocytes were tested for their ability to secrete IFN-γ in response to in vitro stimulation, using the ELISPOT assay as follows. MultiScreen™-IP 96-well plates (Millipore, Billerica, MA) were coated with 50 μL/well of 5 μg/mL purified mouse-anti-ChINF-γ antibody (Biosource International, Camarillo, CA) in PBS, and incubated overnight at 4°C. Spleen tissue was squeezed through a 70-μm mesh in RPMI 1640 culture medium. Splenocytes were isolated by density gradient centrifugation using Ficoll-Hypaque 1.077 (Amersham Pharmacia Biotech), washed twice in PBS, and adjusted to 1×107/mL in complete culture medium consisting of RPMI 1640 with glutamine, supplemented with 2% chicken serum (Sigma-Aldrich), 8% FCS (Biological Industries, Bet-Haemek, Israel), and 1% tissue culture antibiotics (Biological Industries). The coated plates were washed three times with sterile distilled water and incubated with 250 μL/well of complete culture medium for 2 h at room temperature. The medium was discarded from the plates, and splenocytes were seeded in triplicates at 100 μL/well. Cells were incubated in the presence of either complete culture medium, or medium supplemented with one of the following antigens, to a final volume of 150 μL/well; inactivated IBV (M41), rS1, and rN. A negative protein control was the modified E. coli heat-labile enterotoxin (rLT), prepared as described previously (38). Concanavalin A (Con A; Sigma-Aldrich) was used as a cell-viability control. All antigens were used at a final protein concentration of 30 μg/mL. The cells were incubated for 20 h at 37°C and 5% CO2. Consequently, the cells were discarded and the plates were washed six times with washing buffer (PBS containing 0.01% [v/v] Tween 20), twice with distilled water, twice with washing buffer, and twice with PBS. In the following steps detection reagents were diluted in filtered PBS containing 0.5% (w/v) BSA. Biotinylated mouse-anti-ChINF-γ (Biosource International) was added at 1 μg/mL concentration. The plates were incubated for 2 h at room temperature, followed by six washes with washing buffer. Streptavidin-HRP conjugate (Biosource International) was added at 1:3000 dilution (100 μL/well), and the plates were incubated for 45 min at room temperature. After three washes with washing buffer, and three washes with PBS, 100 μL/well of the chromogen tetramethylbenzidine (TMB; KPL, Gaithersburg, MD) was added, combined with TMB membrane enhancer, according to the manufacturer's instructions. As the assay developed, dark blue spots appeared on sites bearing horseradish peroxidase. After 15 min, the plates were washed thoroughly with copious amounts of tap water. The plates were air-dried overnight in a dark, ventilated place. The results were recorded and analyzed using the ELISPOT Bioreader® 4000 B-Pro (Byosys, Leipzig, Germany), and calculated as the index of IFN-secreting cells, based on the net ratio of sample/positive-control counts.
Statistical analysis
The anti IBV antibody levels detected by ELISA and HI tests, and the ELISPOT mean values resulting from the different vaccines, were analyzed by SigmaStat software, version 2 (Jandel Scientific, San Rafael, CA). Pair-wise multiple comparison analysis with ANOVA or one-way testing (Dunn's method) was applied.
Results
Expression of rS1 and rN in E. coli
Production of rN and rS1 was induced in all colonies carrying the transgenic genes. A positive colony for each protein was chosen for further study. Following 3 h of stimulation with IPTG, the bacteria were disrupted and centrifuged. The supernatant and pellet of the selected colonies were analyzed by SDS-PAGE and Western blotting (Fig. 1). Both fragments were detected by antibodies against poly-histidine. The 54-kDa rN fragment was detected in the supernatants of the centrifuged cultures, and the 59-kDa rS1 fragment was detected in the pellet.
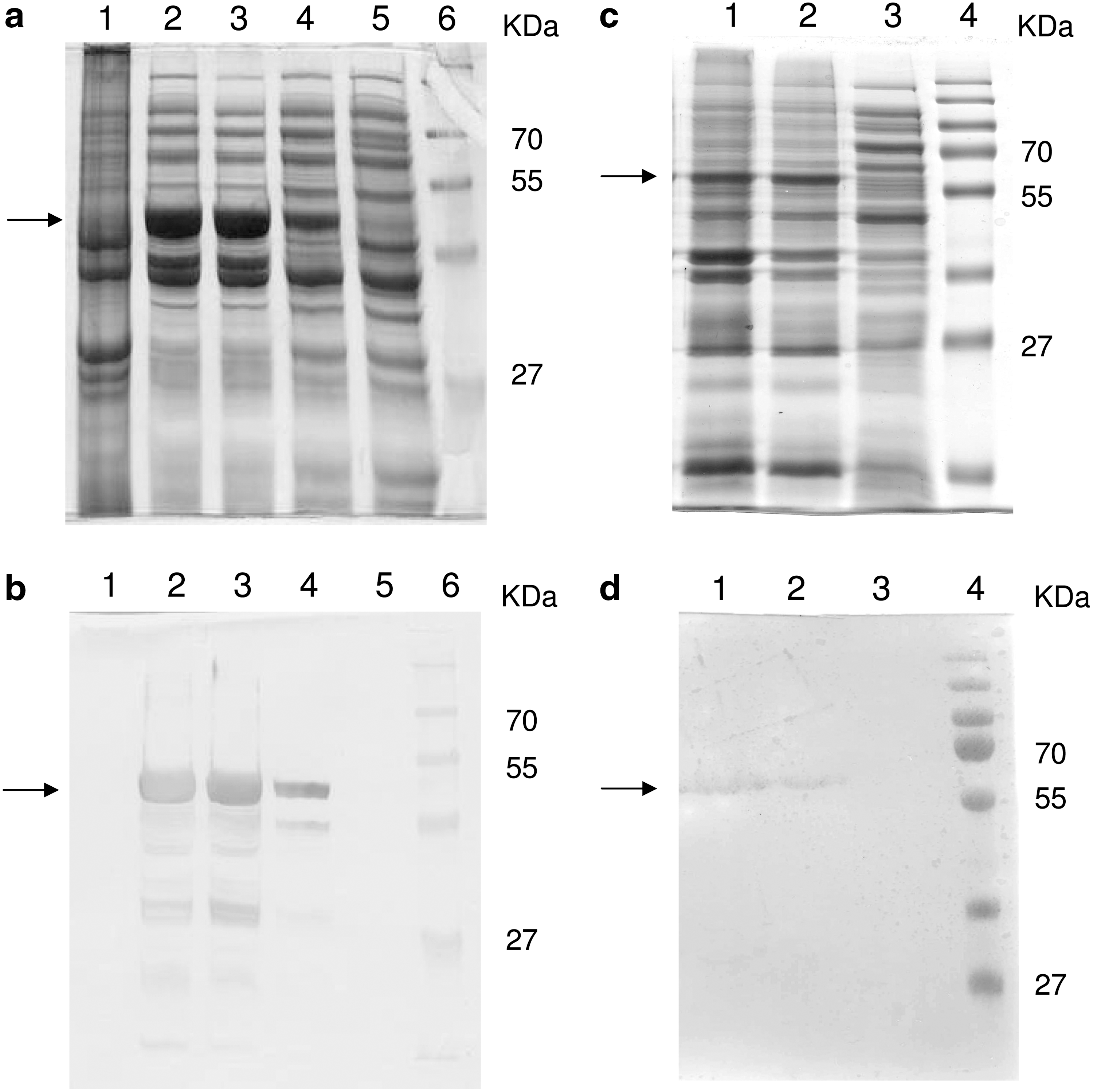
Identification of rN and S1 expressed in E. coli. (
Systemic humoral immune response
Induction of systemic antibody levels by the different vaccines was measured at 14 and 49 d of age, and before and after challenge in trial 2, at 35 and 44 d of age, respectively. The group vaccinated with rN did not survive the trial due to a technical complication, thus the partial serology results of this group were not included in the final analysis. An increase in HI antibody titers was measured at 14 d, and again at 49 d of age, in chickens vaccinated by eyedrop with either H120, or with rS1 (p<0.05), compared to chickens vaccinated subcutaneously with rS1+rN or non-vaccinated chickens (trial 1, Fig. 2a). When tested 1 week after the last vaccination at 35 d of age (trial 2), an increase in HI titers was measured only in the group vaccinated with the commercial vaccine H120 (p<0.05; Fig. 2b).

Two weeks after the first vaccination by eyedrop, antibodies against IBV were undetectable by the commercial ELISA kit in all groups, with measured levels being below the ELISA kit threshold (<396) in both trials. Low antibody levels were maintained throughout trial 1 and increased only after challenge in trial 2. The same low results were obtained after a subcutaneous vaccination with rS1+rN (results not shown). Nine days post-challenge, a significant increase in IBV antibody levels was detected in the groups vaccinated with either H120 or rN, compared to the rS1 or non-vaccinated groups (p<0.05; trial 2, Fig. 2c).
With all vaccinating antigens, no statistically significant increase in systemic IgA production was detected. Specific IgG levels against M41 increased significantly at 35 d of age in the group vaccinated twice with H120 (0.355 OD 620 mm), compared to the non-vaccinated group (0.149 OD 620 mm; p<0.05). In chickens vaccinated with one of the recombinant proteins, the mean IgG values were below those of the non-vaccinated group.
Local humoral immune response
Levels of IgA and IgG antibodies against M41 were measured in tracheal washes sampled at 35 d of age from non-vaccinated and birds vaccinated with H120, rS1, and rN. IgA levels were significantly higher in the birds vaccinated with H120 (p<0.05), compared to all other groups tested (Fig. 3a). Low IgG levels were measured in birds from all groups, with some increase in the group vaccinated with H120 (not statistically significant; Fig. 3b).
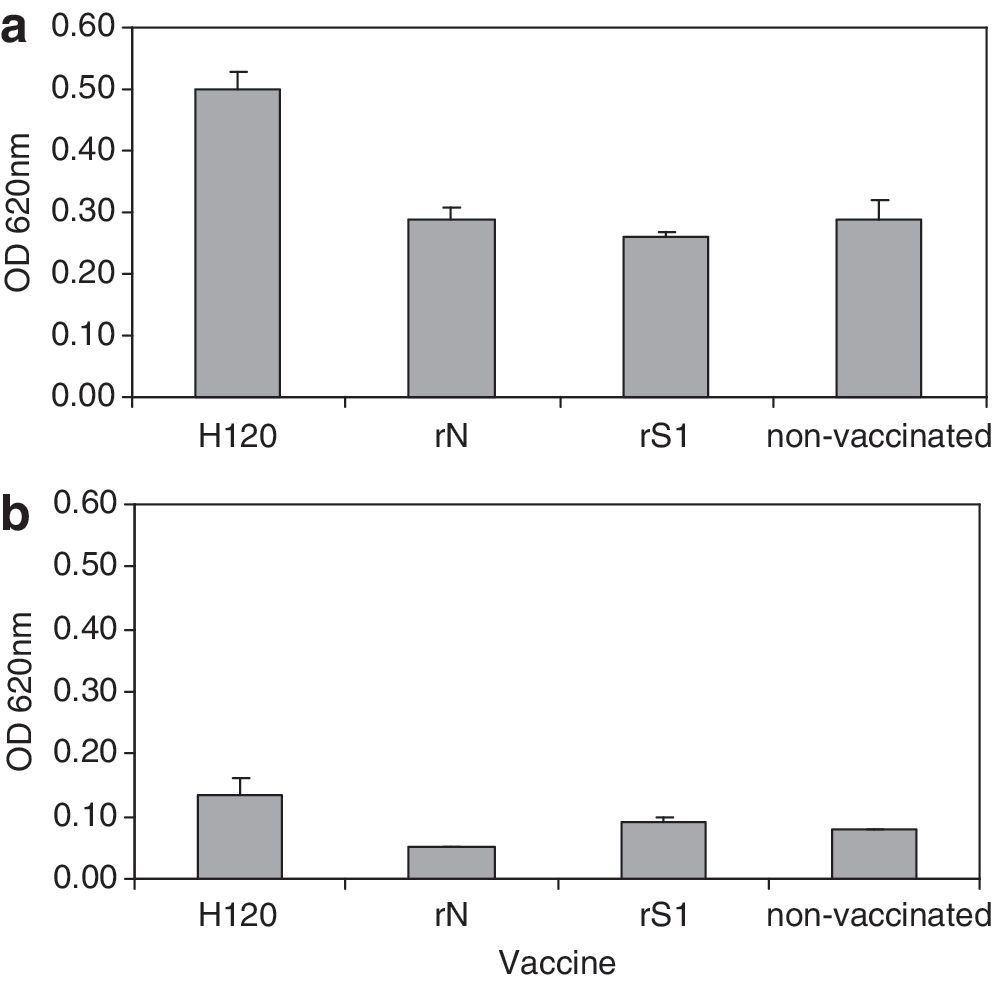
IgA (
Cell-mediated immune response
The cell-mediated immune response was assessed using the ChIFN-γ ELISPOT assay with spleens from non-vaccinated and vaccinated chickens with rS1, rN, and H120 from trial 1. ChIFN-γ production by splenocytes increased significantly following in vitro stimulation with either the inactivated M41 virus, rN, or rS1, in chickens vaccinated with the commercial vaccine H120 (p<0.05), compared to unvaccinated chickens (Fig. 4). Likewise, an increase in ChIFN-γ production by splenocytes was recorded after stimulation with the inactivated virus, in chickens vaccinated with either rN or rS1, compared to unvaccinated chickens (p<0.05), indicating a cellular response to stimulation by the whole virus. A low, statistically non-significant response was obtained in the groups vaccinated with H120 or rS1, after in vitro stimulation with the unrelated protein rLT.
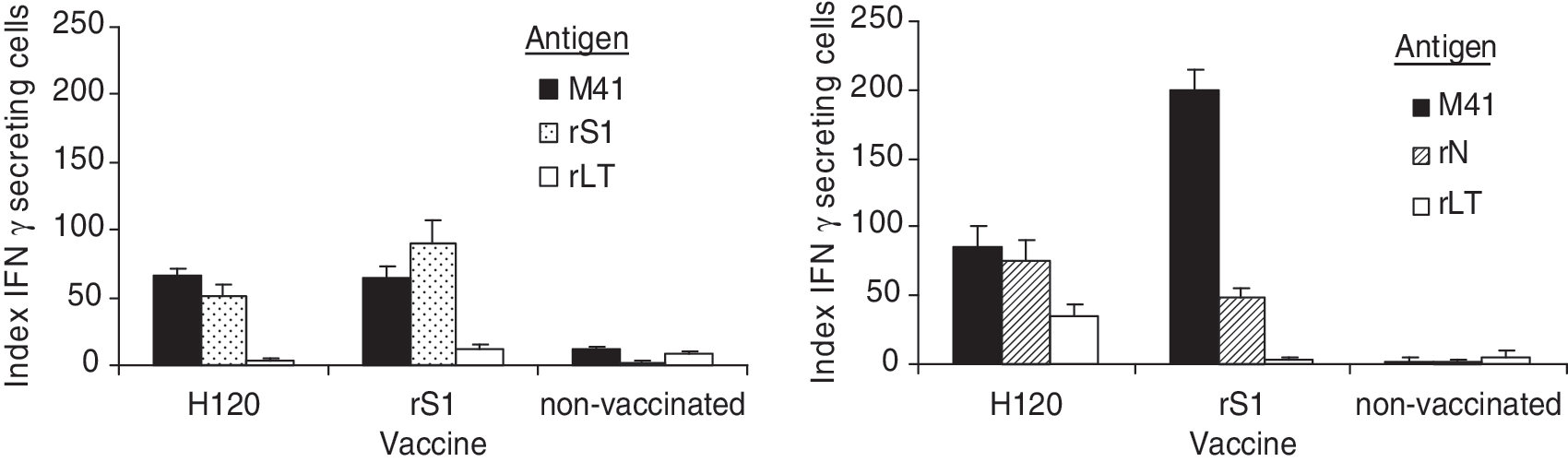
ChIFN-γ secretion by splenocytes of vaccinated and unvaccinated chickens, following in vitro stimulation. Results are expressed as the index of IFN-γ secreting cells (the net ratio of sample/positive control [ConA-stimulated] counts).
Protection
The efficacy of the different vaccines was evaluated in trial 2 by detection of the challenge virus in tracheas 5 d post-challenge, at 35 d of age. Two consecutive vaccinations with H120 conferred 91% protection (1/9 IBV positive/no. of challenged birds). Three vaccinations with rS1 provided 40% protection (6/10 positive birds). Three vaccinations with rN provided no protection, compared to the non-vaccinated group (9/10 positive birds).
Discussion
The viral glycoprotein S1 and the nucleocapsid protein N of IBV are known for their immunogenic properties (17). In the present study we tested the potential of these recombinant proteins to induce an immune response by mucosal vaccination.
The S1 and N DNA fragments of the expected sizes (1610 bp and 1230 bp, respectively) of the reference strain M41 were inserted into plasmids, transformed in E. coli cells, and successfully expressed. The recombinant proteins were tested for their ability to induce humoral and cell-mediated immune responses, compared to the live attenuated vaccine H120. Although it is well known that the S1 subunit induces humoral protective immunity (7,17), knowledge of the mechanism behind protection against clinical disease is incomplete. While there is evidence that local antibody secretion prevents re-infection (37), circulating and local antibodies at high titers do not always correlate with protection (12,24).
Vaccination with the live attenuated vaccine H120 conferred protection against the homologous live virus, although it induced low IBV antibody levels, below the sensitivity threshold of a commercial IBV ELISA kit. Inefficient induction of antibodies by live attenuated vaccines has been demonstrated before (12,32). A more pronounced systemic response to the live vaccine was detected by ELISA 9 d post-challenge, indicating a secondary immune response. An increase in specific IgG or HI antibodies against the homologous virus was measured as well. HI levels increased with time and were higher at 49 d of age (trial 1), compared to the levels measured at 35 d of age (trial 2). In addition, H120 induced a local humoral response, as demonstrated by an increase in IgA levels in tracheal washes of vaccinated birds. Vaccination with rN induced cross-reactive antibody production as well, as was detected 9 d post-challenge, in accordance with previous reports (17).
Vaccination with rS1 induced humoral immune responses similarly to the live vaccine (i.e., very low cross-reactive antibody levels). HI antibody levels were low at 14 d of age, but reached levels similar to the group vaccinated with H120 at 49 d of age (trial 1). In trial 2, HI antibodies were measured only at day of challenge (i.e., 35 d of age), and were lower than in the control group. This difference between trials may be due to genetic differences in the participating chickens. Significant differences in the immune response to rS1 compared to the live virus included lack of a tracheal IgA and of specific IgG antibodies in sera of rS1-immunized birds, and a lack of any secondary humoral response after challenge in that group. As local antibody production seems to play an important role in protection against IBV (12,37), the bacteria-expressed rS1 seems to be inferior to the live vaccine in this respect.
An important element in conferring resistance to IBV is the cell-mediated immune response. Its significance was demonstrated by adoptive transfer of IBV-primed cytotoxic T cells (34), delayed-type hypersensitivity assays (9), and evidence of significant helper T-cell infiltration into the respiratory and kidney tissues of IBV-infected chickens (21). In this study ChIFN-γ secretion was used as a metric of cell-mediated immunity in response to the vaccinations, based on studies demonstrating its involvement in the acquired cellular response to IBV (15,30). The ELISPOT assay was used to quantify ChIFN-γ-secreting splenocytes of vaccinated chickens. Both rS1 and rN proteins, as well as the live H120 vaccine, induced enhancement in the number of splenocytes that secrete ChIFN-γ following in vitro stimulation with the inactivated strain M41, while secretion by cells derived from non-vaccinated chickens remained low. These results indicate the presentation of epitopes by antigen-presenting cells to the cellular immune system following uptake of recombinant proteins or the whole virus. This finding is supported by studies that established the existence of such epitopes on the S1 and N proteins of IBV (4,5,19). The cell-mediated immune response induced by rN was significantly higher than that induced by either rS1 or H120. Although the S1 protein carries T-cell epitopes (19), it appears that rS1 was less effective than rN in eliciting a cell-mediated immune response to the whole virus. One possible explanation for the difference in immune responses might lay in a more efficient presentation of N peptides by antigen-presenting cells following effective uptake, processing, or presentation on MHC receptors of these cells. The relatively low response in splenocytes derived from H120-vaccinated chickens may be attributable to differences between H120 and M41 in the composition of their structural proteins (unpublished data), indicating a specific response by the rN-vaccinated chickens to stimulation by the homologous virus M41. It was not possible to characterize subpopulations of ChIFN-γ-secreting cells, as depletion of either CD4+ or CD8α+ T cells decreases ChIFN-γ production considerably (2). A longer incubation time of the cells with an antigen might have been useful in observing changes in the subpopulation ratios, but most chicken splenocytes do not survive after 40 h of incubation, due to a lack of commercial growth factors for chicken cell cultures. The cell-mediated response induced by rN, and possibly by rS1, failed to protect against infection with live virus. Pei et al. showed that protection by adoptive transfer of primed T cells can be dose-dependent (31). Thus, while in an in vitro assay a known number of viable cells are in contact with the antigen, in the in vivo rN- and rS1-vaccinated chickens, effector T cells may have been below protective levels.
Presentation of peptides by MHC, followed by induction of a T- cell response, is solely dependent on the protein sequence. While some B-cell epitopes are linear, others are conformational epitopes. Thus denatured proteins can induce antibody production, which would then be measured by ELISA, as was the case with the rN vaccination, or possibly by the HI assay in case of the rS1 vaccination. Consequently the limited humoral response, and the inefficient protection by rS1, can be attributed to a lack of some conformational B-cell epitopes, the formation of which may depend on post-translational modifications (e.g., glycosylation). This may explain the absence of local IBV antibodies, in parallel with a significant increase in the cellular immune response in birds vaccinated with rS1.
The aim of this study was to estimate the immunogenic potential of two IBV structural proteins, expressed in a prokaryotic system, and administered by eyedrop. An innovative finding is that by mucosal vaccination, and without the assistance of live vectors or any adjuvant, both rS1 and rN induced a specific cell-mediated immune response against the whole virus. This finding has important applications for the development of subunit vaccines to IBV, as well as other pathogens. In order to improve the humoral response to rS1, future studies should be directed towards a search for an efficient eukaryotic expression system that will incorporate post-translational glycosylation, and possibly improve the presence of conformational-dependent B-cell epitopes. As mentioned previously, the major problem in vaccination against IBV is the frequent changes in the virus. Vaccines based on selected fragments of the S1 protein of different viral strains, combined with the more conserved N protein, may provide a broader range of protection, and at the same time avoid the risk of genomic recombination. An efficient expression system, and a practical route of administration, may render the viral proteins rS1 and rN as suitable candidates for vaccines based on recombinant viral proteins, which would be safe to use and relatively simple to adapt to new variants.
Footnotes
Acknowledgment
This study was supported by Phibro, Bet Shemesh, Israel.
Author Disclosure Statement
No competing financial interests exist.